Individuals with anorexia nervosa are able to severely restrict food consumption and maintain an extremely low weight (
1). Because there are no proven treatments that normalize core symptoms in adults with anorexia nervosa, this is often a chronic disorder resulting in high morbidity and mortality. In healthy individuals, hunger is dysphoric and increases reward salience and subjective value of food to drive consumption (
2). In contrast, individuals with anorexia nervosa often describe eating as anxiogenic, and food refusal may reduce dysphoric mood (
3). Do individuals with anorexia nervosa have an altered response to the motivating signals of hunger? While individuals with anorexia nervosa have been shown to have alterations in neural mechanisms coding motivation, salience, and valuation of food when ill and after weight restoration (
4), few studies (
5) have systematically examined whether there is altered functioning in this circuitry in individuals with anorexia nervosa when hungry that might persist after recovery. Understanding the mechanisms contributing to the neurobiology of self-starvation in anorexia nervosa could identify new treatment targets.
One method of identifying food-related neural mechanisms coding motivation, salience, and valuation is to measure brain response to palatable tastants, such as sucrose, using neuroimaging (
6). This approach has identified a neural circuit in humans (
7) that extends beyond pure gustatory chemosensory processing to include regions involved in motivation and reward processing, and which maps onto analogous circuits in rodents (
8) and nonhuman primates (
9). Specifically, tastes of sucrose signals are transmitted from sweet taste receptors in the tongue through the brainstem and thalamus to the primary gustatory cortex in the insula. In humans, the anterior insula receives chemosensory taste input from the mid-insula (
10). Recent studies suggest that the insula taste cortex identifies sweetness, and the amygdala, which is involved in processing emotions, codes the valence, specifies its hedonic value, and elicits execution of selective behaviors (
8). The hypothalamus, a homeostatic center, regulates metabolic processes, including hunger and food intake, with motivation-reward systems associated with the hedonic drive to eat (
11). In primates, the orbitofrontal cortex processes the reward value of taste (
7) and the ventromedial prefrontal cortex acts as a visceromotor area and governs the hypothalamus, amygdala, and insula. Ultimately, taste information is integrated with reward value and homeostatic drives (
7,
11) via projections from the anterior insula and amygdala to the striatum (
9) to guide motivated eating behavior. In healthy individuals, hunger, compared with satiety, increases brain response in these reward, attention, and motivation regions in response to palatable foods (
6,
7,
12,
13).
In this study, we used a palatable taste task with functional MRI (fMRI) to investigate the response to tastants in the hungry and fed states in the food reward-motivation circuit. We studied women in remission from anorexia nervosa to avoid the confounding effects of malnutrition on this circuitry, in comparison to healthy control women. Our previous studies (
14,
15) supported our hypothesis that women in remission from anorexia nervosa relative to control women would show a diminished response to taste of sucrose in the insula and striatum when in a hungry compared with a fed state, suggesting that hunger may not generate signals to motivate eating (
7) in anorexia nervosa. We also considered whether anorexia nervosa might entail an exaggerated response to being fed (enhanced satiety) and a normal response to hunger. Thus, we performed a group-by-condition-by-tastant interaction analysis in the insula, striatum, amygdala, and medial orbitofrontal cortex to test these hypotheses in regions that integrate sensory/hedonic aspects of taste and interoceptive awareness in the service of homeostasis.
Discussion
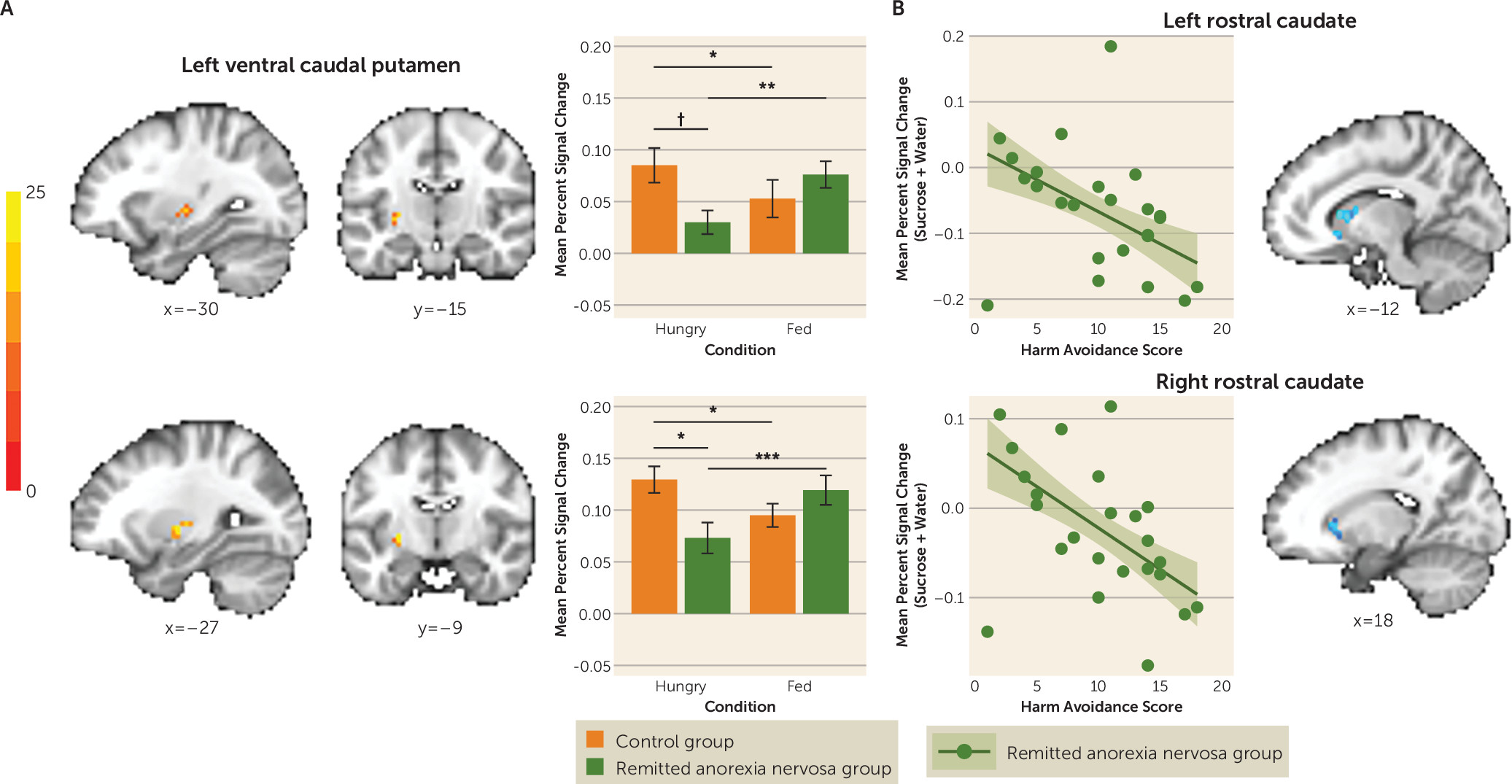
The relentless ability to restrict eating and become severely emaciated has been one of the most puzzling symptoms of anorexia nervosa. In this study we found that in women in remission from anorexia nervosa, striatal and, at a lower statistical threshold, insular activation was abnormally unresponsive to taste stimulation in the hungry state. As expected, this circuitry, which guides motivated behavior, such as eating (
7), was activated in control women when hungry compared with being fed (
6,
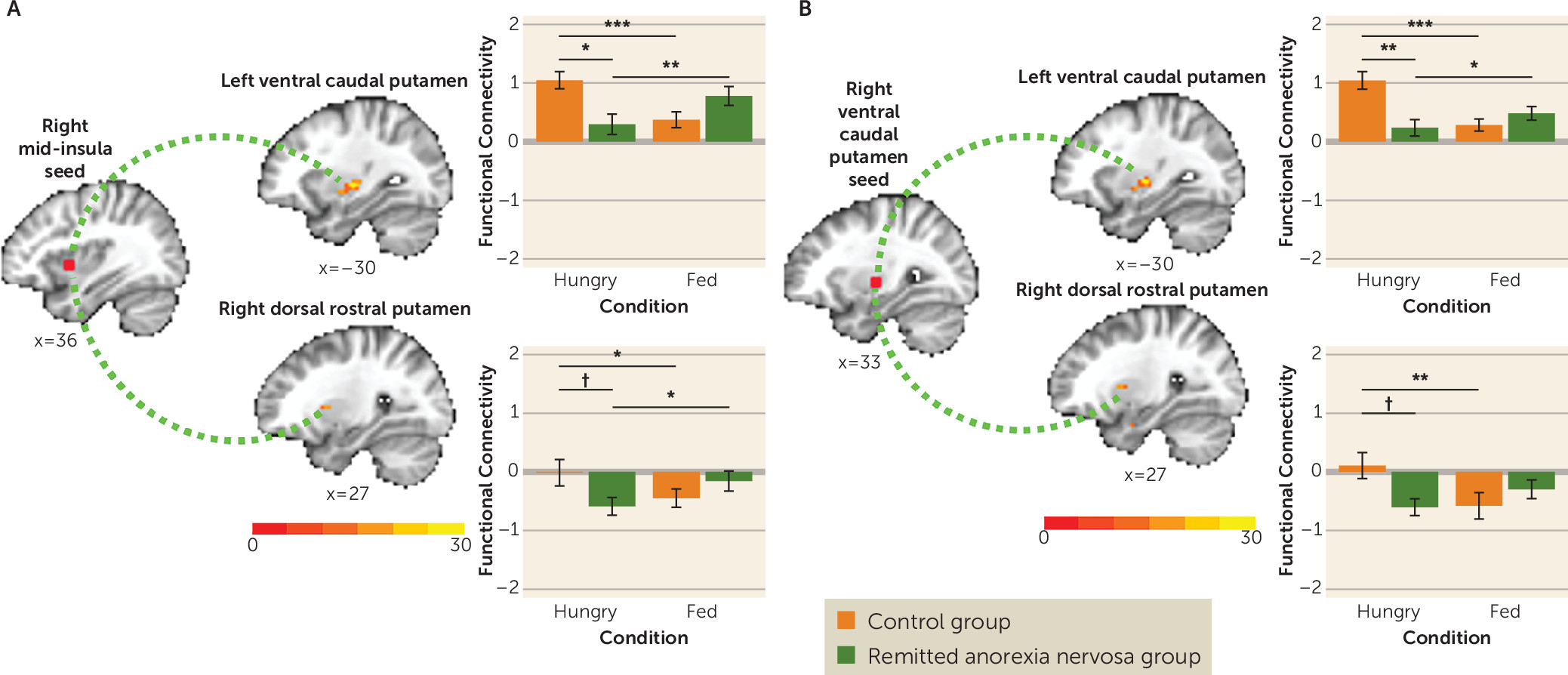
12). Specifically, we identified a region of the ventral caudal putamen in which 1) the control group showed an increase but the remitted anorexia nervosa group showed a decrease in BOLD response to tastants when hungry compared with when fed, and 2) the remitted anorexia nervosa group showed a lower BOLD response to tastants compared with the control group only when hungry. Functionally, this region of the putamen is connected to the anterior and mid-insula (
22), and, at a lower statistical threshold, the remitted anorexia nervosa group showed a similar response pattern that was opposite that of the control group in the left anterior insula. Moreover, the control group showed increased mid-insula-to-ventral caudal putamen functional connectivity to tastants when hungry compared with when fed, whereas the remitted anorexia nervosa group showed decreased functional connectivity between these regions when hungry compared with when fed, and compared with control women when hungry. Overall, the interaction suggests that in women with remitted anorexia nervosa, the response to taste after eating may be “normal,” but the response to taste during hunger is abnormal. Research is needed in symptomatic groups using paradigms that measure food-specific reward value. We speculate, however, that our results are compatible with two possible main processes: the afferent metabolic signals that are translated into motivational behavior (food seeking) are attenuated and/or the afferent signals are excessively suppressed by top-down modulatory brain regions. Future investigation will need to disambiguate these possibilities.
We postulated that when hungry, women with remitted anorexia nervosa would show less response to sucrose (a food and primary reward) compared with water. If there was a deficit specifically related to sucrose consumption, it might be possible to test this, because water and sucrose have been shown to activate different groups of neurons in the primary taste cortex in nonhuman primates (
23). Counter to our hypothesis, we did not detect differences in response to water and sucrose. However, fMRI studies may not have the resolution to distinguish between response to water and sucrose in subregions of the primary taste cortex or related regions (
12,
24). Furthermore, water and sucrose similarly activate taste circuitry and regions associated with emotion and motivation (
7,
12). Because we did not detect response differences between water and sucrose, we collapsed responses across tastants to examine group and condition effects.
Reduced activation to tastants when hungry in the ventral caudal putamen and reduced functional connectivity between the right and left ventral caudal putamen are consistent with other studies showing disruption in limbic striatal circuitry in individuals with current or remitted anorexia nervosa (
16,
25,
26). Nonhuman primate studies have shown that the ventral caudal putamen receives sensory-limbic inputs from the insula, as well as from the amygdala, orbitofrontal cortex, and temporal lobe, and is distinct from the sensorimotor dorsal putamen, which is afferently regulated by the motor cortex (
9). Human neuroimaging studies have shown that it is functionally (
22) and structurally (
27) connected to prefrontal limbic and premotor cortex regions (
Figure 4B). This circuitry is integral to reward processing and the preparation for and control of actions triggered by external stimuli (
28). Reduced recruitment of the striatal aspects of this circuitry in response to tastants when hungry may impair the translation of taste reward value to motivated eating behavior in anorexia nervosa. However, the molecular mechanisms contributing to the selective sensitivity of this region to hunger signaling in anorexia nervosa require further study.
At a lower threshold, the remitted anorexia nervosa group showed reduced activation to tastants when hungry in the anterior insula (left anterior short gyrus and middle short gyrus) and reduced functional connectivity between the right anterior (middle short gyrus and posterior short gyrus) and mid-dorsal insula and ventral caudal putamen. The insula is a hub for interoception (the awareness and integration of internal body signals to regulate behavior) (
10), and accumulating evidence suggests disturbances in anterior and mid-dorsal insular function in individuals with current or remitted anorexia nervosa (
14,
15,
29–
31). The human gustatory cortex maps to the anterior and mid-insula (
7). Moreover, the mid-insula is a chemosensory region involved in relaying various somato- and viscerosensory signals to the anterior insula and striatum (
7,
29). Specifically, the mid-dorsal insula integrates gustatory information with information about the body’s homeostatic needs (hunger and satiety signaling) to modulate feeding behavior. The anterior insula projects along the ventromedial axis of the striatum, including the rostral ventral striatum and ventral caudal putamen (
9), to mediate and regulate goal-directed behavior, such as the consumption of palatable food (
7). Together, these findings raise the question of whether there is a disconnect between the insula and striatum in anorexia nervosa, resulting in a failure to integrate taste information with motivational and homeostatic drives (
10).
Our findings here are, to our knowledge, the first to suggest that individuals with remitted anorexia nervosa have an altered insular and striatal response to taste that is moderated by metabolic state. Our earlier studies of tastant responses in remitted anorexia nervosa similarly showed hypoactive response in the anterior insula, ventral putamen, and other striatal regions (
14,
15). There was less supervision of prestudy eating in these early studies, raising the possibility of reduced food intake prior to scanning. Comparison of the present findings with those of our earlier studies is also confounded by updates in neuroimaging statistical methods (
32) and circuit identification (our earlier studies averaged activation across an anatomically defined region of interest rather than performing voxel-wise analyses within an anatomical search region).
Few studies have compared hungry and fed states in anorexia nervosa. Using a different design, Cowdrey et al. (
33) also found altered insular and striatal function during taste processing in remitted anorexia nervosa. Holsen et al. (
5) showed similar hungry-state hypoactivation in the anterior insula and limbic regions in response to pictures of food compared with objects in anorexia nervosa independent of illness state. Holsen et al. (
5) also noted that earlier studies showed less consistent results, possibly because of methodological differences. Our study only assessed neural response to taste, but other studies suggest altered mid-insular and striatal responses to other interoceptive stimuli (
30,
31), reward prediction error (
25,
34), and food images (
35). We previously showed reduced ventral striatal response to monetary rewards when hungry (
16), suggesting that a dysfunction of homeostatic influences on neural processing of salient stimuli or reward is not restricted to food in anorexia nervosa but may generalize to secondary reinforcers (e.g., money) as well. Evidence that nutritional state strongly affects reward and interoceptive processing in anorexia nervosa has significant implications for assessing these constructs and may explain the mixed findings in previous studies that did not manipulate or control for metabolic state.
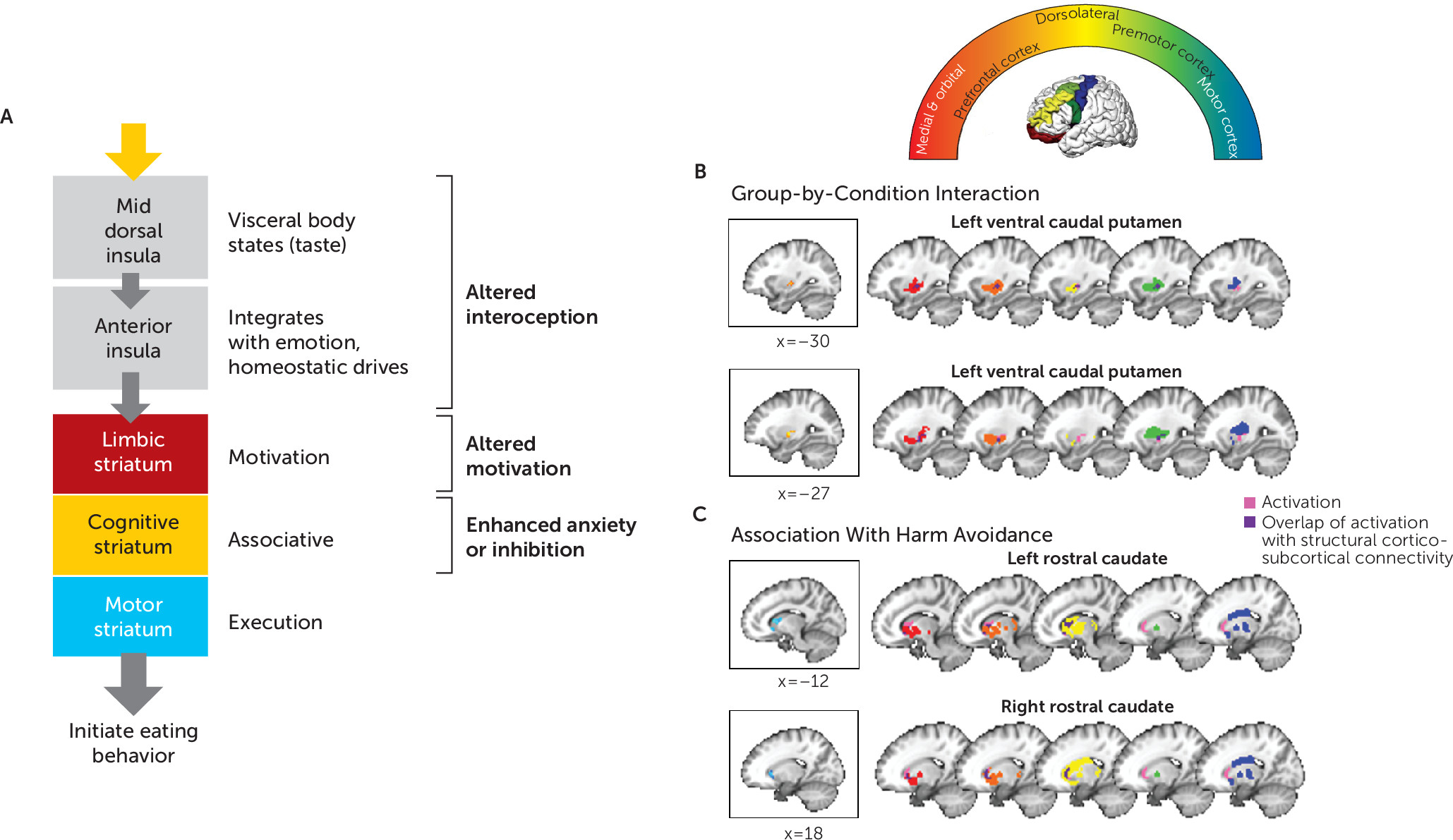
Participants in the remitted anorexia nervosa group who were more harm avoidant showed less activation in response to taste when hungry in regions of the rostral caudate known to receive input from orbitofrontal and medial prefrontal cortices as well as the dorsolateral prefrontal cortex (
Figures 2B and
4C). This raises the possibility that high harm avoidance is associated with reduced engagement of the striatal limbic and associative control circuits that translate food reward to motivated eating, and could be related to altered dopaminergic function. Harm avoidance—a construct of anxiety, inhibition, and inflexibility (
36)—and anxiety alone have been associated with caudate dopamine availability in healthy human subjects (
37) and rodents (
38). Altered dopamine metabolism (
39,
40) is found in remitted anorexia nervosa, and dorsal striatum function measured by BOLD response or dopamine metabolism has been linked to elevated anxiety, harm avoidance, and sensitivity to punishment in anorexia nervosa (
25,
40,
41). Dorsal striatum dopamine signaling also plays a role in feeding (
11). Dopamine-depleted mice, which do not initiate feeding behavior, will resume normal eating after restoration of dopamine selectively in the dorsal striatum, but not the ventral striatum (
42). Thus, dorsal striatum dopamine signaling may serve as a permissive “action initiation” signal, promoting nutritive food retrieval and consumption in response to metabolic demand (
42). In the context of these previous findings, we speculate that a lower dorsal striatum response to taste when hungry among participants with the most pronounced behavioral inhibition in the remitted anorexia nervosa group may reflect a reduced eating action initiation signal. More research using tasks that include behavioral responses is needed to test this hypothesis. Other potential explanations of dorsal caudate dysfunction in anorexia nervosa include efficient cognitive control (
43), development of habitual eating behavior (
44), or anxiety associated with eating (
41).
Taken together, these findings highlight circuitry that may play a key role in pathological eating in anorexia nervosa (
Figure 4A). Of note, this circuitry has also been implicated in appetitive changes associated with major depression: increased appetite is associated with greater response to food stimuli in limbic reward circuitry, whereas appetite loss is associated with hypoactivation within the anterior and mid-insula (
45). The pathophysiology driving these disturbances in anorexia nervosa remains to be determined. It is possible that top-down processes inhibit these signals or that there is altered homeostatic system modulation (
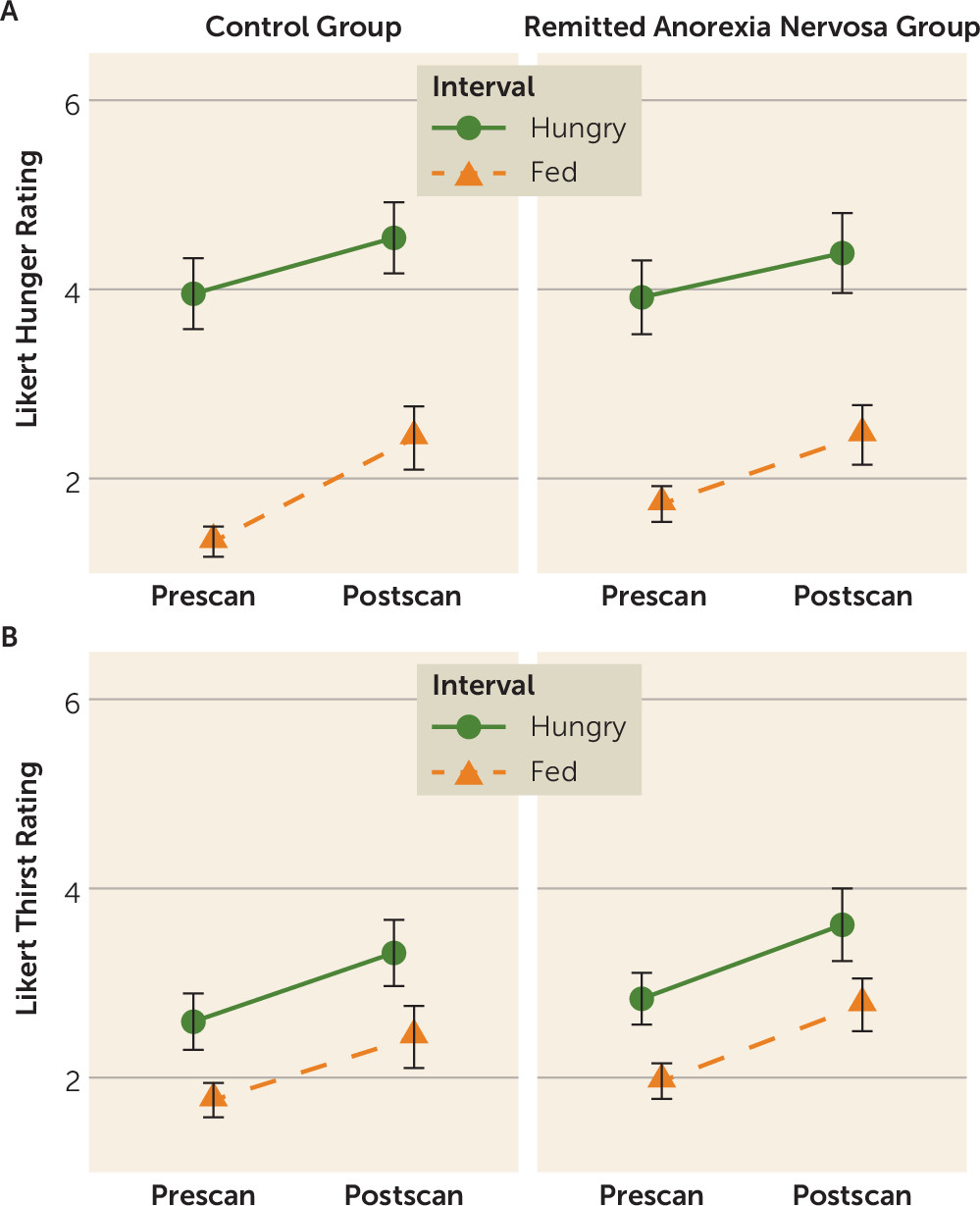
4). Interestingly, our participants with remitted anorexia nervosa reported hunger and fullness levels similar to those of the control participants (
Figure 1), raising the possibility that bottom-up hunger signaling is intact but not accurately translated within the insula-striatal network that motivates action. We propose (
Figure 4A) that disturbance in the mid or anterior insula may result in a distorted signal about hunger or feedback about energy balance, the disturbance in the ventral caudal putamen may result in diminished motivation, and a lower caudate response may inhibit initiation among the most harm-avoidant individuals, each serving to maintain food avoidance. Whether developing anorexia nervosa requires disturbances in all three of these processes (interoception, motivation, inhibition) or whether having any one disturbance is sufficient to impair the signal through this network remains uncertain. Factors such as a distorted body image may initially drive restricted eating. If individuals are also vulnerable to failing to respond to homeostatic and reward signals that stimulate eating when starved, they thus may be able to maintain food avoidance.
Limitations and Future Directions
This study has several methodological strengths. We included only participants who had been weight stable (<3 kg weight change) and physiologically and cognitively remitted from anorexia nervosa for at least 1 year, and we systematically monitored and manipulated prescan nutritional status. The study also has some limitations. The passive task was designed specifically to characterize differences in hard-wired circuitry underlying neural sensitivity to gustatory processing, limiting our ability to assess top-down cognitive control, decision making, or learning (
46). Unexpectedly, all participants rated the water as more pleasant-tasting than the sucrose solution and both the water and sucrose solutions as moderately pleasant-tasting. This likely limited our ability to isolate activation associated with sweet taste reward. Additionally, these ratings were made only after the task was completed; future studies should include pre- and postscan pleasantness ratings.
Although signal dropout precluded inclusion of the orbitofrontal cortex and hypothalamus in our gustatory-reward circuit mask, exploratory voxel-wise whole brain analyses (see the
online supplement) were consistent with the region-of-interest results, with extended findings in thalamic, medial prefrontal, and parietal regions. Although we balanced diagnosis across scanners (χ
2=0.37, p=0.60) and participants underwent scanning on the same scanner for both scanning sessions, differences in the magnet hardware may have influenced findings between participants. Since individuals with anorexia nervosa have dysfunctional eating (unlike our participants with remitted anorexia nervosa), it is not clear from our cross-sectional study whether dysfunction within this insular and striatal network in remitted anorexia nervosa is a trait-level alteration or a scar of being underweight. Reward- and anxiety-related alterations associated with anorexia nervosa persist after symptom remission and may even be present in childhood (
4,
47); we therefore hypothesized that our study would detect trait-like alterations in neural activation. Changes in other circuits not studied here (e.g., executive control circuits [
48]) may be required to compensate for these persistent reward- or anxiety-related alterations to promote normalized behavior. In our sample, BOLD response was not associated with duration of illness or remission, providing further support for trait-like alterations; however, longitudinal research among individuals at risk for developing anorexia nervosa is needed to understand alterations that may predispose individuals to anorexia nervosa and persist after remission. Moreover, further research using paradigms that directly assess food reward valuation and approach in currently symptomatic individuals with anorexia nervosa will be necessary to more rigorously test our hypothesized association between reduced hungry-state activation and reduced motivation to eat.
Finally, our results indicate hungry-state differences in the neural response to predictable receipt of uncertain tastants (sucrose or water). These results cannot be generalized to all food stimuli. For example, other studies suggest different response to aversive taste processing (
33,
49). It remains unknown why differences appear specific to hunger and not satiety, and further research is needed to understand the effects of hunger-state differences on unpredictable tastants.
Implications
Treatments for anorexia nervosa are only marginally effective because we lack an understanding of the underlying neural mechanisms of the disorder. If reduced recruitment of the neural circuitry implicated in translating reward signals to motivated behaviors when hungry interferes with food approach and eating initiation after restriction in anorexia nervosa, particularly among individuals with high anxiety, pharmacological and psychotherapeutic strategies that directly target these processes may be beneficial. For example, this circuitry involves dopaminergic projections. Recent findings for olanzapine and aripiprazole (
50–
53) suggest that investigation of medications targeting the dopamine system may improve eating behavior and reduce anxiety in anorexia nervosa. In addition, these findings support investigating behavioral strategies for enhancing initiation to eat or compensating for altered homeostatic drives (
54,
55). Finally, a registry aimed at understanding how individuals with anorexia nervosa recover could be of much benefit in developing new treatment strategies. Such registries have been of use for developing insights into successful weight loss in obesity (
56).