A range of family and twin studies have found that anxiety disorder (AD), among the most common of psychiatric conditions (
1), is familial and moderately heritable (
2,
3). Most, but not all twin studies, suggest that familial resemblance for AD arises solely from genetic effects (
4–
8). Genetic influences on AD have also been detected using genome wide association designs (
3,
9). Multivariate analyses have suggested a substantial sharing of genetic risk factors across various specific anxiety disorders (
10,
11).
However, to date, no adoption study of AD has been reported. While twin studies examine genetic and environmental influences within a generation, adoption studies address a different and equally important question: the magnitude and causes of cross-generational transmission.
We therefore report an expanded National Swedish adoption study of clinically diagnosed AD examining six family types: 1) intact nuclear families, 2) families with not-lived-with (NLW) biological fathers, 3) families with NLW biological mothers, 4) families with stepfathers, 5) families with stepmothers, and 6) biological and adoptive parents from adoption families. We address four major questions: first, what is the magnitude of the transmission of AD from parents to their offspring and to what degree does it result from genetic versus rearing effects? Second, what are the sources of the familial cross-generational relationship between AD and MD? Third, to what degree are the cross-generational genetic and rearing effects which contribute to liability to AD and to MD correlated? Fourth, how do results differ when we examine AD-subtypes or impose a hierarchy on MD and AD diagnoses?
Methods
Information for this study was collected from nationwide Swedish registers (as outlined in appendix Table 1 in the
online supplement). Each person’s unique identification number, having been replaced with serial numbers for confidentiality, were used for registry linkages. Ethical approval was secured from the regional ethical review board in Lund, Sweden. Using the Swedish Hospital Discharge Register, Outpatient Care Register and nationwide primary care data, we diagnosed AD and MD using ICD-8, 9, and 10 codes and phobia, panic disorder, generalized anxiety disorder, and AD-not otherwise specified (NOS), using only ICD-10 codes as seen in appendix Table 2 in the
online supplement. Individuals registered with bipolar disorder or schizophrenia (see appendix Table 2 in the
online supplement) were censored. This study population included all individuals born in Sweden between 1960 and 1992, who were alive and resided in Sweden at least until the age of 20. For these individuals, here called offspring, we included the number of years, during ages 0–15, that they resided in the same household and geographic area as their biological mother, biological father, and possible stepfather, stepmother, adoptive mother, and adoptive father. Study end was on December 31, 2018 due to availability of medical register data. From 1960 to 1985 (every fifth year), we used household identification numbers from the Population and Housing Census to define family types. The household identification includes all individuals registered at the same residence. From 1986 onward (every year), we defined family type using the family identification from the Total Population Register. The family identification is defined by individuals registered at the same property who are also related, adopted, married, or had common children. Therefore, from 1986 we could not detect all stepparents, e.g., stepparents not married to a biological parent, nor having a common child with a biological parent. For years with no information on whether offspring and parents resided together, we used information from the closest year. Geographical areas called Small Areas for Market Statistics (SAMS) are defined by Statistics Sweden, the Swedish government-owned statistics bureau. There are approximately 9,200 SAMS throughout Sweden, and are characterized by homogeneous building types and limited by natural boundaries.
We defined the following types of families: intact families that included offspring residing from ages 0 to 15 years in the same household with both their biological mother and father; families with NLW (not-lived-with) father/mother, that included offspring who never resided in the same household or SAMS area as the biological father/mother; two types of stepfamilies (stepfather/stepmother) that included offspring who did not reside the entire period between ages 0 and 15 with the biological father/mother and resided at least 10 of these years with a biologically unrelated man/woman 18–50 years older; adoptive families that included offspring adopted at younger than 5 years, with information available on both adoptive parents and at least one biological parent. The adoptive parents had to be biologically unrelated to the offspring and the offspring had to reside with each adoptive parent for at least 10 years between the ages 0 and 15. As domestic adoptions declined substantially in Sweden in recent decades, we expanded modestly our adoption cohort to maximize sample size, including offspring born from 1955 to 1992. The NLW and stepparents were defined so that their relationships with their offspring resembled those of an adoptee and his/her biological and adoptive parents.
For all family types, parents had to be alive throughout 1975 and had to reside in Sweden during some period of time from 1976 and onwards. There are overlaps in the parent-offspring relationships between the NLW families and the stepfamilies. When doing joint statistical analyses across family types, parent-offspring relationships included in, for example, a stepfather family was removed from the NLW father family group, so that each relationship was only calculated once.
In our parent-offspring pairs, we calculated tetrachoric correlations—which represent the correlation in relatives for a latent underlying normally distributed liability to illness (
16)—for AD transmission, MD transmission, AD to MD cross-transmission and MD to AD cross-transmission. We used this measure because of its ease of interpretability (
17) and its insensitivity to changes in base rates (
18), given the differing rates of AD and MD across our family types. To calculate weighted tetrachoric correlations and for testing for heterogeneity across families, we use a meta-analysis fixed-effects model (
19). The model is fixed as samples of data come from the same population. For the heterogeneity tests, a significance level of 0.05 was utilized. We also calculated the genetic correlation and the rearing correlation between AD and MD (as outlined in appendix Table 3 in the
online supplement).
In cases with both lifetime MD and AD diagnoses, we assigned a single diagnosis based on the relative number of the two diagnoses, and if those are close, giving greater emphasis to recent diagnoses (see appendix Table 4 in the
online supplement). Data analysis was conducted from August 23, 2021, to February 11, 2022. Statistical analyses were performed using R, version 4.0.3 (
20) (see appendix Table 5 in the
online supplement) and SAS, version 9.4 (
21).
Discussion
We have conducted an extended adoption study of AD and MD using Swedish national samples. We review, in turn, our eight key findings.
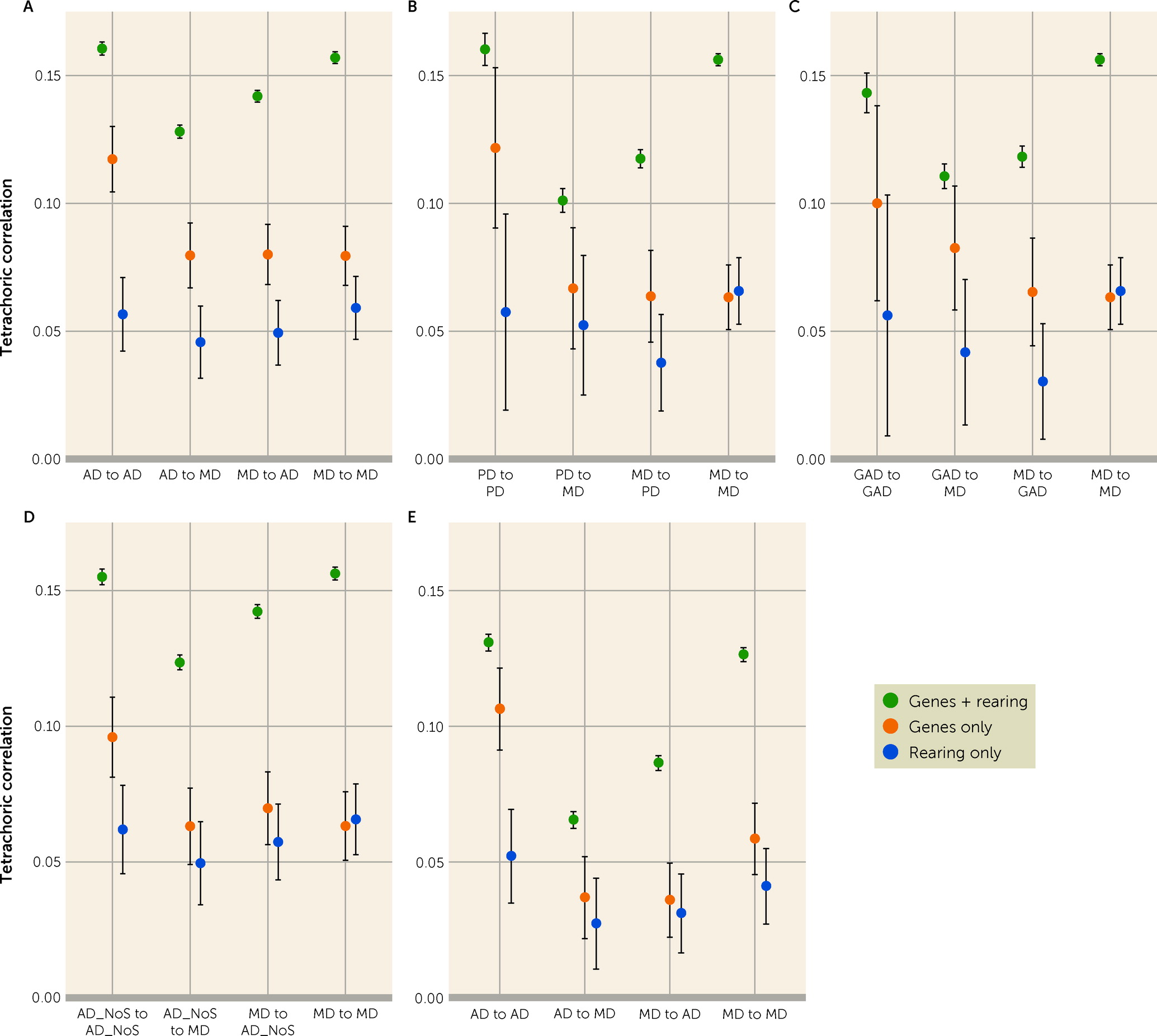
First, assessed with no diagnostic hierarchy, AD is moderately transmitted from parents to their children with a correlation in intact families estimated at +0.16. From our extended adoption design, we are able, for the first time, to determine the sources of this transmission. Approximately 70% of the parent-offspring resemblance for AD results from genetic transmission—estimated at +0.12 and 30% from rearing effects—estimated at +0.06. Our findings are noteworthy as twin studies of anxiety disorders have shown, with near uniformity, that twin resemblance for AD results solely from genetic factors (
4–
8). These results are not necessarily contradictory, as twin studies are poorly powered to detect the modest sibling resemblance that arises from rearing effects (
22).
We cannot, in our sample, easily clarify the mechanisms of the rearing effect of AD. Social learning may contribute. In humans and primates, the fear of objects or situations can be learned from observing fear responses in others (
23). Parental anxiety disorder might be associated with suboptimal parenting (e.g., by causing overprotectiveness) and/or resulting in parental strife or separation which could predispose to AD in offspring. Indeed, high trait neuroticism—associated genetically with anxiety disorders (
24,
25)—in parents is predictive of overprotective parenting (
26) which predisposes offspring to anxiety disorder (
22,
27–
29).
Second, we observed parent-offspring cross-transmission from AD in parents to MD in children as well as from MD in parents to AD in children that were quantitatively only moderately lower than the parallel within-disorder effects. The magnitudes of these transmissions in intact families (+0.13 vs. +0.14) were very similar, as were the transmissions in genes-only (both +0.08) and rearing-only relationships (both +0.05). In Sweden, assuming no diagnostic hierarchy, MD and AD have a symmetrical cross-disorder cross-generation transmission of liability.
Third, we were able to calculate the correlation of the cross-generational genetic influences on MD and AD as equaling +0.83. This result is consistent with a number of twin studies, especially those that have examined generalized anxiety disorder, which report quite high genetic correlations, suggesting a substantial sharing of genetic risk factors between MD and AD (
12–
14). A prior twin study of lifetime MD and generalized anxiety disorder in Sweden, modelled separately by sex, reported a genetic correlation of the two disorders equal to 1.00 in females and +0.74 in males (
13). The mean of these two figures would produce a figure close to that observed here. Of note, this twin study utilized interview-based DSM-IV diagnoses, while our study relied on registry diagnoses, yet produced very similar findings.
Fourth, we also calculated, for the first time to our knowledge, a rearing environmental correlation for MD and AD which was estimated at +0.83. Our findings suggest that the familial environmental factors that are both related to parental diagnoses of MD or AD and that influence risk for MD and AD in offspring are highly inter-correlated, albeit not identical. This high inter-correlation stands in contrast to the considerably lower resemblance of depressogenic and anxiogenic environmental exposures in adulthood as revealed both by twin modeling (
12) and the assessment of specific classes of stressful life events (e.g., loss/humiliation versus danger) (
30–
32). The anxiogenic and/or depressogenic aspects of the home environment and enduring parent-child relationships that we capture in this study likely differ qualitatively from the acute environmental exposures in adulthood that predispose to MD or AD.
Fifth, our analyses permit us to compare the magnitude of the genetic and rearing influences for AD and MD. In most twin samples examining MD and one or more anxiety disorders, MD is more heritable (
13,
33). That is not what we see here. Narrow-sense heritability in adoption designs equals twice the tetrachoric correlation in gene-only relationships which would be estimated as +0.24 for AD and +0.16 for MD, and this difference is statistically significant (p<0.001). These heritability estimates are lower than those seen in twin studies, but this occurs commonly (
34–
37), likely, at least in part, because adoption studies examine genetic effects across different generations, while relative pairs in twin analyses are always of the same age.
Sixth, our expanded adoption design assumes that 1) NLW fathers and mothers are good proxies for biological parents in an adoption design and 2) stepfathers and stepmothers are reasonable proxies for adoptive parents in an adoption design. We could test these assumptions. Of the 16 relevant comparisons of the weighted parent-offspring correlations provided in
Table 3 (that is NLW with biological and step- with adoptive-parents), none were significant at chance-corrected levels, results which support the validity of our expanded adoption design.
Seventh, our sample contained three AD subtypes with sufficient sample size to examine individually with MD. Consistent with prior results showing high genetic correlations across different ADs (
10,
11) and recent molecular genetic work which has focused on an aggregate AD diagnosis (
9), the pattern of results was broadly similar across these three forms of AD, and the aggregate results for all AD. In accord with prior twin studies, generalized anxiety disorder and MD had a modestly stronger genetic relationship than did panic disorder and MD (
33).
Finally, as seen previously (
33,
38–
40), we observed high levels of comorbidity between MD and AD. Previous twin studies of these syndromes have not typically taken this into account, in part because of uncertainty about an optimal approach. To attempt to address this knotty problem, we used a Procrustean approach by imposing a diagnostic hierarchy on comorbid cases based on number and recency of diagnoses so that all comorbid MD and AD cases were assigned a single diagnosis of either MD or AD. We then repeated our analyses. The major change in findings were reductions in the genetic cross-disorder parent-offspring correlations and, as a result thereof, a sizable drop in the MD-AD genetic correlation. Not surprisingly, these findings suggest that the substantial MD-AD comorbidity contributes to their high genetic correlation. Further conceptual and empirical work will be needed to determine an optimal approach to “correcting” for MD-AD comorbidity in genetically informed analyses.
Limitations
These results should be interpreted in the context of two potentially important methodological limitations. First, this study is restricted to subjects with physician-diagnosed MD and/or AD in Sweden. We could not evaluate affected individuals who never presented for treatment. Our treated cases were likely, on average, more severely affected than those who would be detected by personal interviews in community samples. It is difficult to determine whether such incomplete ascertainment is more likely to attenuate or upwardly bias estimates of parent-child resemblance. Obtaining our subjects as we did from hospital, specialist, and primary care registries, however, means that we did not need to rely on subject cooperation or accurate recall. Such data can produce false negative and false positive diagnoses, the nature of which is difficult to estimate. Sweden has no large-scale interview-based psychiatric epidemiological study with which to compare our treated prevalence rates. However, an epidemiologic survey in near-by Norway provided a lifetime estimate of MD of 17.8% and estimates for phobias, panic disorder, and generalized anxiety disorder ranging from 4.5% to 14.4% (
41). 42,161 interviewed twins from the birth-certificate based Swedish twin registry produced an estimated lifetime prevalence of MD of 19.5% and generalized anxiety disorder of 6.8%. These compare to our estimates in our intact families for AD and MD of 13.5% and 14.3% respectively. The validity of both the AD and MD diagnosis from primary care data has been supported by its prevalence, sex ratio, sibling and twin correlations and associations with well-documented psychosocial risk factors (
42,
43).
Second, to maximize power, we analyzed offspring born across a long 32-year period. To investigate cohort effects, we repeated our analyses for all AD cases in the older and younger half of our cohort. These results are summarized in Appendix Figures 2 and 3 in the
online supplement and, respectively, Tables 17–20 and 21–24 in the
online supplement. While the broad pattern of findings is quite similar across the two subcohorts, parent-offspring correlations from intact families (genes plus rearing) are significantly higher in our younger cohort. Of the eight correlations for rearing or genes only correlations across subcohorts, none were significantly different, correcting for multiple testing (see appendix Table 25 in the
online supplement).