Schizophrenia is commonly a chronic, debilitating disorder with lifelong consequences for affected individuals and families. Schizophrenia also is a heterogeneous disorder with respect to illness presentation, clinical course, and outcome. There has been renewed interest in investigation of inflammatory abnormalities in schizophrenia, stimulated in part by our increased understanding of brain-immune interactions in other chronic medical disorders. Recent genome-wide association studies have led to the identification of associations between genes involved in the regulation of the immune system and schizophrenia risk, perhaps most notably complement protein C4 (
1–
3). Prenatal maternal infection with bacterial, viral, and parasitic infectious agents is a replicated risk factor for schizophrenia (
4), and the inflammatory response to infection is a potential common pathway underlying this association. There is a bidirectional association between autoimmune disorders and severe infections, which are associated with increased inflammation and schizophrenia (
5,
6). There is evidence that patients with schizophrenia have inflammatory abnormalities in the blood, cerebrospinal fluid, and central nervous system (CNS), including levels of inflammatory markers, number of immune cell numbers, and antibody titers (
7–
10). Patients with schizophrenia may have an increased prevalence of certain comorbid infections and associated inflammation (
11). Several trials have found that adjunctive treatment with nonsteroidal anti-inflammatory drugs (NSAIDs) or other anti-inflammatory agents may be associated with improvements in psychopathology among some patients with schizophrenia (
12,
13), and that baseline blood inflammatory markers may predict response to these agents (
14,
15). Taken together, these findings support the plausibility that inflammation may be involved in the etiopathophysiology of schizophrenia, at least for some patients.
It is important to emphasize, however, that the evidence for inflammation in schizophrenia has been fraught with substantial heterogeneity, including negative studies and small magnitude of effect sizes for many associations. There are many potential explanations for such discrepancies, including small sample sizes of individual studies, different phases of illness studied, and potential confounding factors (e.g., smoking, obesity, medication effects, and medical comorbidities). Nevertheless, another important potential explanation is that inflammation may be present in only a subset of patients with schizophrenia. For example, many psychiatric hospitalizations in the late 19th and early 20th centuries were caused by psychotic symptoms in the context of advanced neurosyphilis. Subsequently, the recognition of an infectious cause of psychosis and the discovery and development of penicillin were game changers in the treatment of these patients. The identification and characterization of a potential subgroup of patients with schizophrenia and inflammation may thus hold promise of novel disease- and life-altering treatment approaches.
In that regard, two case reports provide compelling evidence for potential mechanistic associations between the immune system and schizophrenia. Sommer and colleagues (
16) described a 67-year-old man with chronic lymphocytic leukemia and no past psychiatric history who developed new-onset psychosis status after receiving an allogenic peripheral blood stem cell transplant from a brother with chronic schizophrenia. Conversely, Miyaoka and colleagues (
17) reported on a 24-year-old man who experienced prolonged remission of treatment-resistant psychosis—without any antipsychotic medication—after receiving a bone marrow transplant for acute myeloid leukemia. Of course, is it challenging to establish causality, but the bidirectional nature of these case reports provides support for the plausibility of such an association.
In this article, we present a selected review of studies informing on schizophrenia as a potential inflammatory disorder, with emphasis on replicated findings. Because a review of all aspects of immune function in schizophrenia is beyond the scope of the present work, the reader is also referred to related comprehensive reviews (
18–
22). We present a theoretical framework in an attempt to integrate these findings, and we suggest potential mechanisms whereby inflammation may play an etiopathophysiological role in a subset of patients with schizophrenia (
Table 1).
The Immune System and Inflammation
The immune system encompasses two major divisions, the innate and adaptive immune systems. Innate (nonspecific) immunity is the first-line defense against infection and/or cellular injury. Innate immunity provides an immediate, maximal response against a potential pathogen. Whereas it was previously thought that the innate immune system did not elicit immunological memory, increasingly it has been shown that this arm of the immune system may have immunological memory, in that cells may be trained via metabolic and epigenetic modifications, hence referred to as “trained immunity” (
23). Important cells of the innate immune system include mononuclear phagocytes (neutrophils, monocytes, and macrophages), which can engulf and destroy pathogens, and natural killer cells, which can destroy virus- or tumor-infected cells. By contrast, the adaptive immune system responds to specific pathogens or antigens (i.e., a foreign particle that triggers antibody production). In response to a foreign antigen, B-lymphocytes are stimulated to produce antibodies to neutralize the antigen, and to create immunologic memory of that antigen, which can produce a rapid response in the case of re-exposure. Antigen presentation also activates T-lymphocytes. CD8 T-lymphocytes can destroy infected or damaged cells. CD4 T-helper (Th) lymphocytes coordinate and maximize adaptive immune responses.
Cytokines are key regulators of inflammation—the complex response of blood vessels to injury, whether caused by infection or cell damage and dysfunction—that coordinate both arms of the immune system. Cytokines are key immune signaling molecules that exert effects by binding specific receptors on a variety of target cells in both the periphery and the brain. Soluble cytokine receptors can either inhibit (e.g., soluble interleukin-2 receptor [sIL-2R]) or enhance (e.g., sIL-6R) the biological activity of cytokines. Endogenous cytokine receptor antagonists (e.g., IL-1 receptor antagonist [IL-1RA]) compete with cytokines for membrane-bound receptors. Cytokines are also key regulators of the acute phase response.
Activated immune cells have characteristic cytokine profiles. In response to an antigen and the local milieu, naïve CD4 Th lymphocytes can differentiate into a variety of effector cells that direct diverse immune functions. Th1 lymphocytes, which secrete interferon (IFN)-γ, IL-2, and IL-12, promote proliferation of CD8 T-lymphocytes and maximize the killing efficacy of macrophages. Th2 lymphocytes secrete IL-4, IL-6, and IL-10, which stimulate proliferation of B-lymphocytes and neutralizing antibody production. Th3 lymphocytes produce transforming growth factor (TGF)-β and IL-10 and are involved in mucosal immunity and protection. Th17 lymphocytes produce IL-17, which mediates inflammation and has been implicated as an effector of autoimmune and other inflammatory disorders. IL-6 induces Th17 cell differentiation and inhibits differentiation of regulatory T cells. Regulatory T cells, which express the transcription factor forkhead box P3, suppress adaptive T-cell responses and prevent autoimmunity. Activated monocytes and/or macrophages enhance the inflammatory response via production of IL-1β, IL-6, IL-8, and tumor necrosis factor (TNF)-α. Concomitant with increased production of pro-inflammatory cytokines, the compensatory anti-inflammatory response syndrome is a counter-regulatory mechanism that inhibits the primary inflammatory response and involves adaptive reprogramming of leukocytes (
24). Mediators of the compensatory anti-inflammatory response syndrome include IL-1RA, the soluble TNF receptor (sTNF-R), and TGF-β.
There is significant complexity in regard to how cytokines transmit inflammatory signals from the peripheral immune system to the CNS to drive inflammation in the brain (
25). Historically, the brain has largely been considered an immune-privileged organ, although recent evidence has revealed a lymphatic vasculature in the meninges that suggests a link between the peripheral immune system and the CNS, allowing for central immune surveillance (
26). Prior to this important finding, the field’s understanding of peripheral-central communication focused on two pathways: humoral and neural. One primary challenge regarding immune signaling to the brain is that inflammatory cytokines are relatively large molecules, which have difficulty crossing the blood-brain barrier (
27). The humoral pathway involves cytokines passing through leaky regions of the blood-brain barrier (such as the circumventricular organs) or cytokine binding to transport molecules on the blood-brain barrier. This pathway via the blood-brain barrier may be of specific interest in regard to patients with schizophrenia, who have been shown to have dysfunction of blood-brain barrier integrity and function (
28). Alternatively, the neural pathway involves the binding of inflammatory cytokines to peripheral afferent nerve fibers, which, in turn, translates signals into central inflammatory signals. Another more recent finding is that of a third pathway, the cellular pathway, where activated immune cells, primarily monocytes, are trafficked to the brain via chemokines, such as monocyte chemoattractant protein 1, that are produced by microglia, the resident immune cells of the brain (
29). Future work to understand relationships between peripheral and central inflammatory markers in schizophrenia will further elucidate the relevance of these mechanisms. Neuroimaging of microglial activation (see below) from patients with schizophrenia, and modeling of microglial signaling from induced pluripotent stem cells from patients with schizophrenia, will further advance our understanding of peripheral-CNS communication.
The brain-gut axis and the microbiome as it relates to the immune system has also been a topic of interest to psychiatry. Microbes that reside in the mucosa of the gastrointestinal tract often have a symbiotic relationship with their host and play a significant role in immune system regulation (
30). The microbiome is sensitive to a number of environmental factors and exposures, including diet, infection, stress, and medications. Various genetic factors, including those related to immune genes, place individuals with altered microbiota at risk for immune dysfunction. This altered gut flora, or dysbiosis, can activate the immune system via binding to pattern-recognition receptors associated with microbe-associated molecular patterns, which in turn leads to production of inflammatory cytokines via activation of the nuclear factor kappa-B pathways and the inflammasome (
31,
32). Gut microbiota may also communicate with the brain via interactions with the endocrine system (e.g., hypothalamic-pituitary-adrenal axis), the autonomic nervous system, the enteric nervous system, as well as signaling via the vagus nerve (
33). Studies of the microbiome in patients with schizophrenia have demonstrated a lack of diversity in gut microbiota, with some evidence for specific bacteria having a relationship with symptom severity (
34–
36), and there is increasing evidence that altered gut microbiota in patients may be related to immune dysfunction (
37,
38). Future directions in this area may involve clinical trials targeting the microbiome to improve dysbiosis and immune alterations in the gastrointestinal system.
Autoimmunity in Schizophrenia
There is increased prevalence of several autoimmune diseases—which are associated with inflammation—among patients with schizophrenia (
62,
63) and other nonaffective psychoses (
64), as well as among their first-degree relatives (
63,
64), compared with control subjects. These autoimmune disorders include acquired hemolytic anemia, celiac disease, interstitial cystitis, Sjogren’s syndrome, and thyrotoxicosis (
63,
64). The association between schizophrenia and autoimmune disorders appears to be bidirectional. Both a personal (
63) as well as a family history (
64) of any autoimmunity is associated with a significantly increased risk of schizophrenia. Conversely, patients with schizophrenia have an increased risk of incident autoimmune disease, and a history of hospitalization for infection acts synergistically on this risk (
6).
A meta-analysis of 81 studies (
19) found significantly increased prevalence of positive titers for 20 different autoantibodies among patients with schizophrenia compared with control subjects. Absolute titers of anticardiolipin immunoglobulin G (IgG) and M (IgM), and nerve growth factor antibodies were significantly increased among the patients with schizophrenia. Lachance and McKenzie (
65) performed a meta-analysis of 12 studies of biomarkers of gluten sensitivity among patients with schizophrenia, finding a significant increase in antigliadin IgG, anti-tissue transglutaminase 2 IgA, and anti-wheat antibodies, suggesting an immune response that differs from patients with celiac disease. A recent study of patients with FEP from China (
66) found higher levels of anticytokine antibodies, including anti-IL-1β, anti-IL-6, and anti-IL-8 IgG. Other meta-analyses (
67) have found an increase in anti-NMDA receptor antibodies among patients with schizophrenia compared with control subjects that is based on high- but not low-specificity seropositivity thresholds. Indeed, these cases of autoimmune encephalitis caused by anti-NMDA receptor antibodies may masquerade as new-onset psychosis. These findings lead to the question of what is the pathophysiologic significance of such autoantibodies among patients with schizophrenia, particularly in the absence of other signs and symptoms of autoimmune disease, and whether these autoantibodies might be targets for novel immunotherapy approaches (e.g., corticosteroids or plasmapheresis).
Inflammation in First-Episode Psychosis
In a meta-analysis of patients with FEP and chronic psychosis (
68), many inflammatory markers were elevated, including IFN-γ, IL-1β, IL-1RA, IL-6, IL-8, IL-10, IL-12, sIL-2R, TGF-β, and TNF-α. There was similar directionality between patients with FEP and chronic schizophrenia for IL-1β, IL-6, sIL-2R, and TNF-α. In another meta-analysis of studies of antipsychotic-naïve patients with FEP (
69), IL-1β, sIL-2R, IL-6, and TNF-α were elevated compared with healthy control subjects, suggesting associations with inflammation that were unrelated to antipsychotic medications. Ultimately more longitudinal data will be necessary to appreciate changes in inflammatory markers over the course of illness in psychosis.
Imaging of Neuroinflammation in Schizophrenia
There is evidence of associations between peripheral blood inflammation, namely IL-6, and smaller hippocampal volumes among patients with chronic schizophrenia (
97,
98) and FEP (
99,
100). A recent study (
101) found that among outpatients with schizophrenia, increased blood levels of IL-6, IL-8, and IL-10 were significantly associated with cortical thickness measurements in the orbital frontal cortex, frontotemporal gyrus, and cingulate cortex. In diffusion tensor imaging, higher blood IL-6 levels have also been negatively correlated with fractional anisotropy and positively correlated with radial diffusion in patients with schizophrenia (
102). In a study of patients with FEP (
103), the percentage of whole-brain gray matter was inversely correlated with IFN-γ and IL-12 levels. Another small study of outpatients with schizophrenia and elevated cytokine mRNA levels (
104) showed a significant reduction in the left pars opercularis (Broca’s area), as well as a negative relationship between IL-1β mRNA levels and left pars opercularis volume. In the North American Prodrome Longitudinal Study (
105), the rate of prefrontal cortical thinning was significantly associated with higher baseline levels of proinflammatory cytokines, and this inverse correlation was significantly greater among subjects who developed a psychotic disorder than among those who did not and among control subjects. Two studies (
106,
107) have also found significantly increased IL-6 mRNA expression in the prefrontal cortex of patients with schizophrenia. A third postmortem transcriptional study (
108) found upregulated gene expression of the IL-6 signaling pathway in the hippocampus, prefrontal cortex, and striatum among patients with schizophrenia compared with control subjects.
Whereas the investigation of inflammation in schizophrenia has largely focused on measurement of peripheral blood markers, there is also significant evidence for increased density and activation of microglia in the brain, reflective of neuroinflammation, that has been demonstrated in both neuropathological and neuroimaging studies (
109–
111). Imaging of microglial activation using positron emission tomography (PET) is a burgeoning field with an increasing number of studies. As the resident immune cells of the brain, microglia release cytokines and other molecular mediators to promote an inflammatory response (M1 phenotype) while simultaneously downgrading the anti-inflammatory response (M2 phenotype) (
112). When activated, microglia express translocator protein (TSPO), and TSPO radioligands have been developed for PET imaging (
113). The findings from these TSPO studies have been mixed. In an individual participant meta-analysis of five studies of second-generation TSPO radioligands, Plavén-Sigray and colleagues (
114) showed that individuals with FEP and schizophrenia had lower levels of TSPO (as measured by total distribution volume [V
T]), compared with healthy control subjects, in the frontal cortex, temporal cortex, and hippocampus. A second, recent meta-analysis of 12 TSPO PET studies showed that binding potential was elevated among patients with schizophrenia compared with control subjects, despite no difference in V
T (
115). The discrepancies between these meta-analyses highlight the distinct challenges in interpreting these studies of neuroinflammation.
Underlying Mechanisms
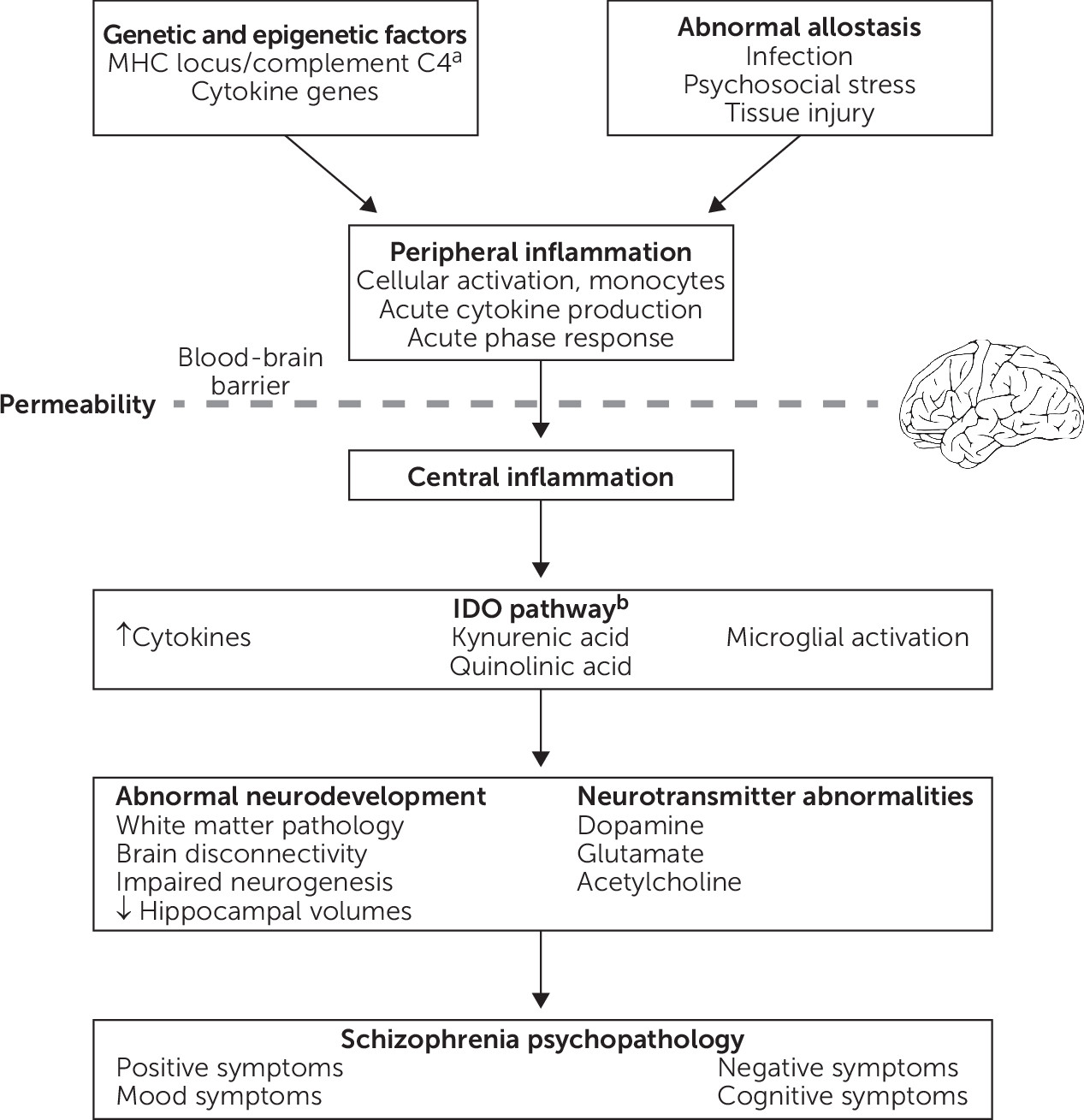
Figure 1 presents a theoretical framework that integrates findings regarding potential mechanistic associations between inflammation and schizophrenia. Briefly, abnormal allostasis interacts with genetic and/or epigenetic factors, which results in peripheral inflammation, characterized by cellular activation, cytokine production, and the acute phase response. In the setting of increased blood-brain barrier permeability, there is also central inflammation, including cytokine production, microglial activation, and activation of the tryptophan catabolic pathway. As a result, inflammation can directly or indirectly affect a variety of processes, including brain connectivity, neurodevelopment, neurogenesis, neurotransmitter function, and white matter function, thereby contributing to schizophrenia psychopathology. For example, increased blood IL-6 in rodents directly modulates dopamine turnover and sensitization to amphetamine-induced locomotion (
125). By contrast, the activity of indoleamine 2,3-dioxygenase (IDO), the rate-limiting enzyme in tryptophan catabolism that is expressed in microglia, can be modulated by cytokines. IDO induction results in increased production of kynurenine, which is converted into the NMDA receptor antagonist kynurenic acid, and this pathway has been implicated in schizophrenia (
126). Inflammation may also play a role in specific symptom domains, such as negative symptoms and cognitive impairment.
Conclusions and Future Research
The decades-long literature on inflammation in schizophrenia is fraught with significant heterogeneity, including negative studies. Many associations between inflammation and schizophrenia have had small-to-moderate effect sizes. Possible explanations for heterogeneous findings include minor inflammation-associated changes that may not be clinically significant, study-related factors (e.g., small sample size and inadequate consideration of potential confounding and/or moderating factors), and/or inflammation that occurs among only a subset of patients with schizophrenia. Regarding the latter, a recent meta-analysis of the variability and distribution of cytokines in FEP did not find evidence for an immune subgroup (
127). In contrast, associations between baseline blood cytokine levels and response to adjunctive NSAID treatment among patients with schizophrenia (
14,
15) raise the possibility of a subgroup of patients for whom inflammation may be a marker of treatment response. Each of these hypotheses warrants further investigation.
Despite substantial heterogeneity, there is replicated evidence for inflammation in schizophrenia in multiple domains, including risk factors, clinical course of illness, and symptomatology (
Table 1). Thus far, inflammatory markers with replicated positive findings across multiple domains include IL-1β, IL-6, and CRP. Although there is evidence for changes in levels of inflammatory markers with antipsychotic treatment (
68,
120), there has been less investigation of inflammation as a marker of response to treatment in schizophrenia. Studies of specific anti-inflammatory strategies, such as monoclonal antibody immunotherapy, are also in their relative infancy. Furthermore, a small number of studies have afforded evidence for a potential causal association between inflammation and schizophrenia. For example, the study by Sekar et al. (
2) has provided compelling evidence that the C4 gene may increase schizophrenia risk by influencing synaptic pruning during critical periods of brain development. In mice, a single maternal injection of IL-6 during pregnancy caused prepulse and latent inhibition deficits in the adult offspring, suggesting the immune response may mediate robust epidemiologic evidence for the association between prenatal maternal infections and schizophrenia risk in the offspring (
128).
In conclusion, inflammation likely plays an important role in the pathology of schizophrenia for at least some patients, although there is not clear evidence to support the assertion that schizophrenia is truly an inflammatory disorder. This area of study shows promise toward a greater understanding of the etiopathophysiology of this heterogeneous syndrome and toward identification of potential illness subtypes, diagnostics, and therapeutics. These efforts aim to reduce illness risk, alleviate symptoms, advance relapse prevention efforts, inform treatment effectiveness, and affect disease progression, ultimately toward more personalized medicine approaches and improved quality of life for at-risk and patients with established schizophrenia.