The anterior cingulate cortex (ACC) lies in a unique position in the brain, with connections to both the “emotional” limbic system and the “cognitive” prefrontal cortex. Thus, the ACC likely has an important role in integration of neuronal circuitry for affect regulation and can be identified as a distinctive region in understanding psychopathology. Affect-regulation, the ability to control and manage uncomfortable emotions, is a primary goal for mental health clinicians in treating psychopathology.
7 Avoidance of painful emotions is often the motivating force in negative behaviors such as substance abuse, binge eating, and suicide. These actions are taken as part of maladaptive approaches to control, avoid, or regulate painful emotions. Clinicians often treat patients by helping them to develop more adaptive coping mechanisms in regulating their emotions. Understanding the processes by which ACC contributes to regulation of emotions may assist clinicians in their therapeutic work.
Anatomy
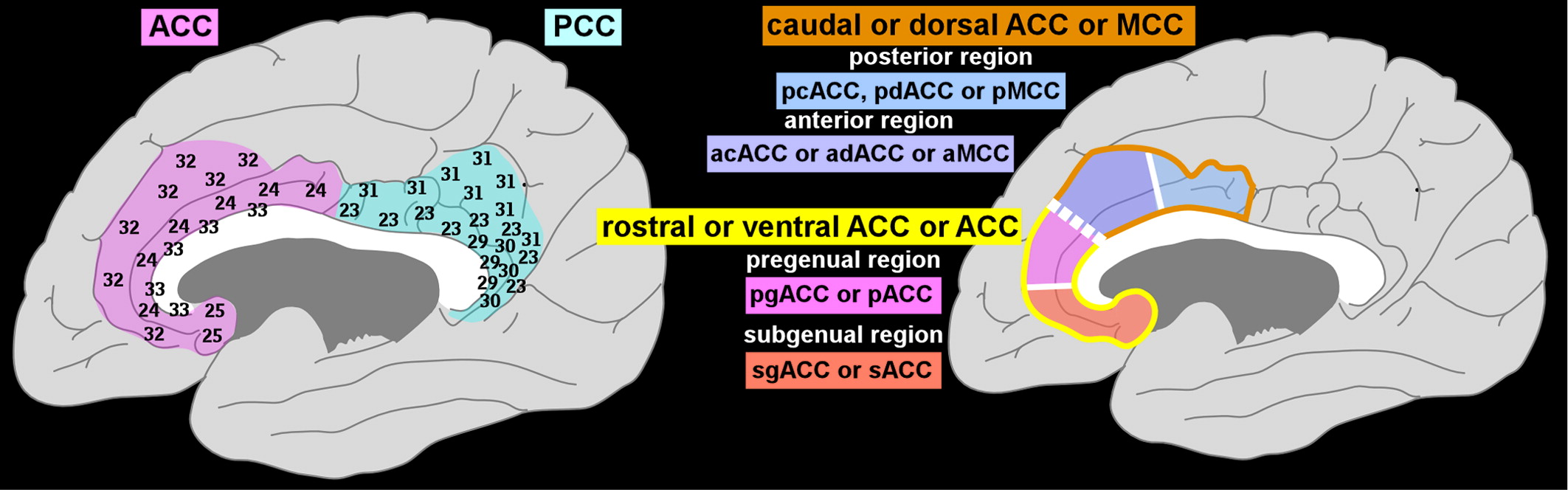
The cingulate cortex lies in the medial wall of each cerebral hemisphere, above and adjacent to the corpus callosum (
Figure 1). This area was originally divided by Brodmann into precingulate (Areas 24, 25, 32, and 33; anterior cingulate cortex [ACC]) and postcingulate (Areas 23, 29, 30, and 31; posterior cingulate cortex [PCC]) regions.
1 There are multiple nomenclatures for this region (
Figure 1). The more-anterior portion surrounding the genu of the corpus callosum has been referred to as rostral or ventral ACC, with the portion adjacent to the PCC considered caudal or dorsal ACC.
2 It has been proposed that it is more appropriate, based on anatomic criteria (e.g., cytoarchitecture, receptor mapping, connections), to split the dorsal ACC off as the middle cingulate cortex (MCC).
1,8 Anatomic studies support further subdivision of both the ACC and MCC.
1,2,8 The MCC is divided into anterior (aMCC) and posterior (pMCC) regions (
Figure 1). The ACC is divided into pregenual (pACC) and subgenual (sACC) regions (
Figure 1). There is also evidence suggesting that Brodmann's Area 25 within the sACC is unique and could be considered a separate region.
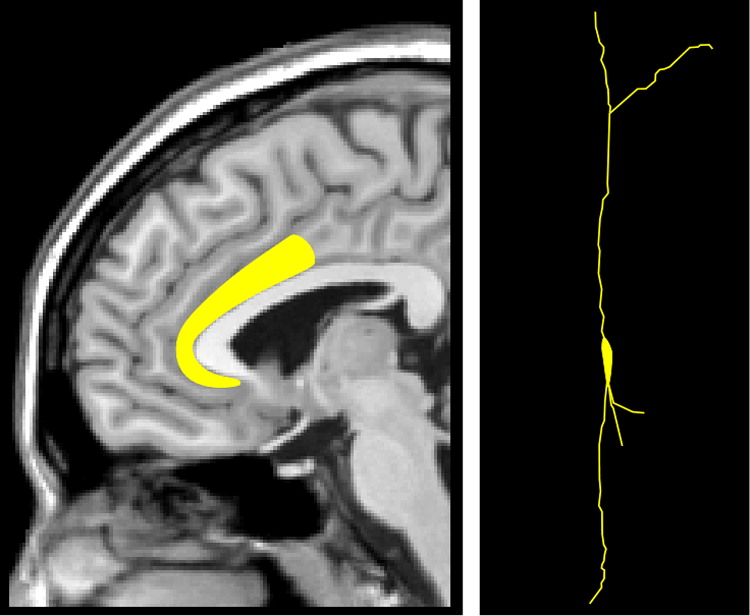
8 Area 25 differs in its cell structure and receptor mapping (e.g., the greatest density of serotonin receptors) from the rest of the ACC and is actually more similar to the MCC in this regard. For clarity, we will use the ACC-MCC terminology proposed by Vogt in this article. An unusual aspect of this region is the presence of Von Economo neurons (spindle neurons), found only in cingulate (pACC and MCC) and insular cortices (
Figure 2).
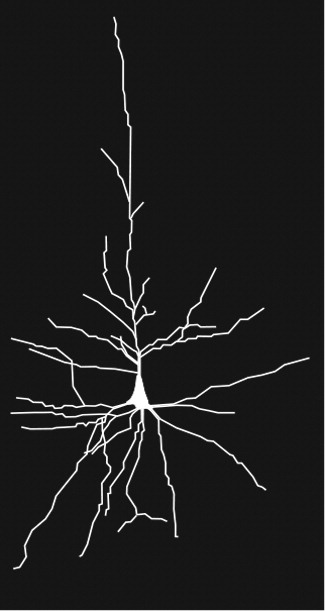
4,5,9–11 Von Economo neurons are present in great apes and humans, but in no other primates. They are more numerous in humans, possibly representing an evolutionary advantage. Von Economo neurons differ from pyramidal neurons in both size and shape (
Figure 2). They are much larger than pyramidal neurons, suggesting faster transmission of information between brain regions, and possibly more connections. They are long, straight neurons, with single, long, apical and basal dendrites, which may indicate that they receive and integrate input from fewer neurons than pyramidal neurons.
5 It has been suggested that Von Economo neurons perform an adaptive function by helping humans and great apes act quickly on an instinctual/intuitive level in social situations.
9 Others have speculated that they may provide fast communication with the anterior insula as part of a salience network.
12,13Multiple areas of the brain (e.g., Brodmann Area 9, medial dorsal nucleus of thalamus, and brainstem monoamine nuclei) have connections with most or all of the cingulate cortex, as indicated by tract-tracing in nonhuman primates. Although important functionally, these connections do not provide clues to specializations within this region. A primary reason for separating the MCC from the ACC is that the connections are quite different, indicating probable differences in functions, as recently reviewed in detail.
14 In brief, the MCC has extensive connections with cognitive (e.g., lateral prefrontal) and motor-related (e.g., premotor and primary motor) areas of cortex and with both pain- and motor-related thalamic nuclei. It also contains the cingulate motor areas, which project to the spinal cord. In contrast, ACC has extensive connections with areas known to be important for emotion (e.g., amygdala), autonomic (e.g., lateral hypothalamus, brainstem centers), memory (e.g., hippocampal region), and reward- (e.g., orbitofrontal cortex, ventral striatum) related functions. As noted above, both MCC and ACC are further subdivided on anatomic grounds (
Figure 1). Compared with the anterior portion of MCC (aMCC), the posterior portion (pMCC) receives stronger projections from inferior parietal cortex, but weaker projections from pain-related thalamic nuclei. In contrast, the aMCC receives strong projections from the pain-related thalamic nuclei and also has strong connections with the amygdala.
15,16 Compared with the subgenual portion of ACC (sACC), the pregenual portion (pACC) has more widespread connections with the lateral prefrontal cortex and much fewer with the amygdala. It also has fewer projections to the ventral striatum.
16,17 The pACC receives projections from the pain-related thalamic nuclei, although less than the aMCC.
15 The sACC has strong reciprocal connections with the amygdala and projects strongly to the ventral striatum and autonomic centers in the hypothalamus and brainstem.
16–18Functional Neuroimaging in Healthy Individuals
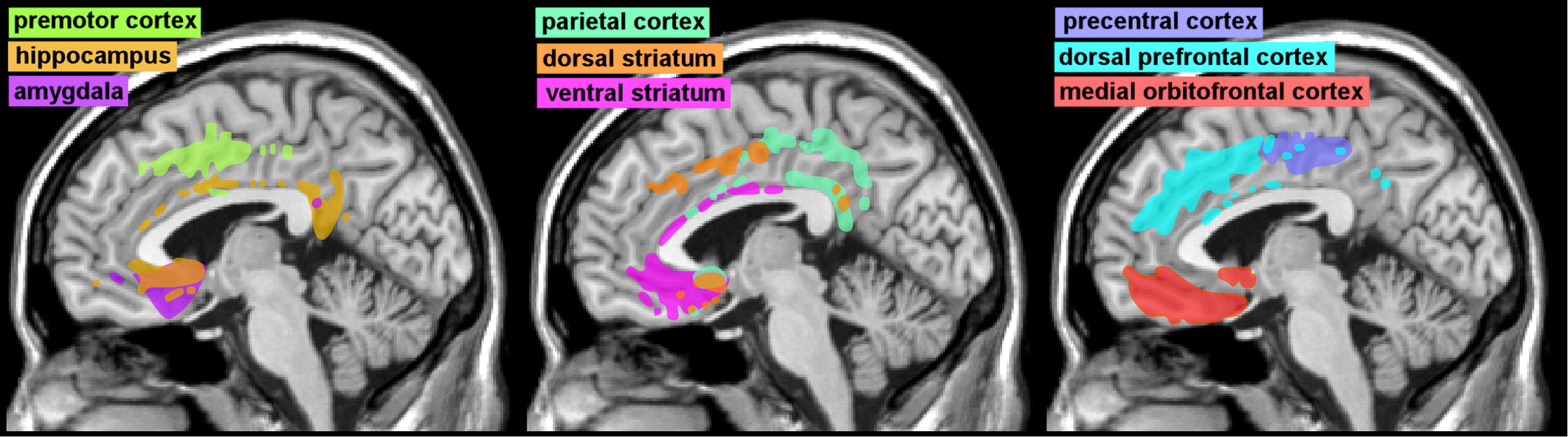
Functional neuroimaging studies support the presence of similar regional differences in connections. Diffusion tensor imaging-based measures of structural connectivity have identified multiple functional regions within cingulate cortex based on patterns and strengths of connections that are very similar to those derived from anatomic studies (Cover and
Figure 3).
6,19 Thus, sACC has the highest connectivity with limbic- (amygdala, ventral striatum, hippocampus) and autonomic- (hypothalamus) related areas, whereas MCC has the highest connectivity with cognitive- (dorsal prefrontal cortex) and motor- (premotor and motor cortex) related areas. Functional neuroimaging-based measures have also been developed to assess functional connectivity. Brain areas in which activity is changing in the same way at the same time (positively correlated) are thought to be functionally connected. Resting-state functional MRI (fMRI) studies indicate that the ACC is most functionally connected with areas implicated in affective processing, with pACC having more widespread connections than sACC.
20–22 Both pACC and sACC were anti-correlated with areas involved in cognitive and sensorimotor processing. In contrast, activity in the MCC was strongly correlated with areas involved in sensorimotor processing, with aMCC also functionally connected with areas important for cognition and anti-correlated with areas involved in affective processing.
20–22 A study of developmental influences found that adults have smaller, more focal, areas of brain activity and more long-distance connections than children.
21 Of note, the sACC was the region with the greatest number of long-distance connections in adults.
Differences among regions of the cingulate cortex in task-related activations also contribute to understanding functional roles. Motor-related tasks activate dorsal MCC, consistent with the locations of the cingulate motor areas.
6 MCC and pACC are both responsive to pain, with MCC more likely to be activated by thermal and pACC by visceral stimulation.
3,15 Simple emotions activate ACC, with pACC more responsive to happiness and sACC to sadness.
15,23 Induction of sadness increases subjective ratings of pain and pain-related activation of the MCC.
24 Reward also activates the ACC; sACC activity correlates with the expected value of options.
6,25 Action-selection and expression of learned fear are more likely to activate pMCC, whereas tasks requiring cognitive control, conflict-monitoring, error-detection, or emotion- (including fear) related appraisal (evaluation) are more likely to activate aMCC and perhaps pACC.
2,3,6,15 Activations related to emotional conflict-regulation and fear-inhibition during extinction are more likely in the sACC.
2 Reappraisal activates both aMCC and sACC.
2 Thus, MCC is “cognitive”—involved in conflict-monitoring and response-selection and execution. Within MCC, aMCC is implicated in emotional appraisal, conflict-monitoring, approach–avoidance decisions, and willed control of actions. pMCC is involved in body-orientation and movement-execution. ACC is “affective,”—involved in emotion assessment, emotion-related learning, and autonomic regulation. Within ACC, pACC is implicated in emotional regulation, autonomic integration, and affect related to pain. sACC is implicated in autonomic control, visceral integration, and conditioned learning.
Self-regulation of emotion is a conscious and voluntary process influenced by multiple factors, including mood and competing regulatory demands.
26 The amygdala is a key limbic structure, and it has a central role in emotion.
27 The cingulate cortex has projections to both the amygdala and the prefrontal cortex. Reaction to emotional stimuli is controlled by a “top-down” emotion-regulation process from several areas of frontal cortex.
28 For example, when the pACC is activated by emotional conflict resolution, reduced activity is seen in the amygdala.
29 Top-down control provides the capacity to regulate an over-activated emotional response. However, both over- and under-regulation of emotion can cause problems.
There are many approaches to self-regulation of emotion. Studies indicate that reappraisal, distraction, distancing, and suppression all decrease ratings of emotion.
30–34 However, amygdala activity was decreased by reappraisal, distraction, and distancing, but increased by suppression.
30–33 Similarly, subjects with high suppression scores had greater activation of the amygdala while viewing neutral images, suggesting that this is a less successful emotional-regulation strategy.
35 Reappraisal, distraction, and distancing were all associated with increased activation of areas in lateral and medial prefrontal cortex, sometimes extending into the MCC.
31–33 Activation of a similar area of medial prefrontal cortex was present when individuals created a negative interpretation of neutral images.
36 In one study, habitual use of reappraisal rather than suppression was associated with the lowest levels of psychiatric symptoms, whereas infrequent and ineffective self-regulation of emotion was associated with the highest level.
37 Thus, increased emotional awareness and attention to when one feels negative emotions may help the individual identify events that contribute to the negative state. Another study reported the idea that regional cerebral blood flow (rCBF), measured by arterial spin-labeling, was elevated in a region of ventromedial prefrontal cortex that included part of the sACC in those with high suppression scores.
38 The authors suggest that this may indicate overactive monitoring of internal state. A study in which male subjects were asked to inhibit their sexual arousal while watching erotic film-clips reported activations in the superior frontal gyrus and pACC but not in limbic-system areas usually activated by such stimuli.
39Thus, activation of the pACC contributes to suppression of the initial limbic-system response, an example of a top-down process. Studies in normal individuals with very different abilities to identify and communicate emotions are also informative. A greater awareness of emotion, or emotional intelligence, has been related to higher overall psychosocial functioning.
40 A series of studies looked at the influence of individual differences in emotional awareness on rCBF (as measured by positron emission tomography) during viewing of emotional films or pictures. They found a positive correlation between emotional awareness and activity in the MCC, but only when the emotional stimuli were highly physiologically arousing (as indicted by skin conductance).
41,42 As noted by the authors, the greater activation of this area in individuals with higher emotional awareness may indicate greater deployment of attentional resources to emotional processing. An fMRI study comparing activation during viewing of emotional pictures in individuals who scored either high or low on a measure of emotional unawareness also reported a positive correlation between the subjects' arousal ratings of the pictures and activation in the MCC.
43 The area of activation was much greater in the High group (low emotional awareness), extending into pACC and sACC. The authors suggested that this may indicate greater effort expended to down-regulate arousal in this group, and noted that suppression is a common emotional-regulation strategy for such individuals.
Functional Neuroimaging in Psychiatric Disorders
Alterations in task-related activations in individuals with psychiatric disorders (as compared with healthy individuals) can inform understanding of psychopathology. Lower-than-normal activations within cingulate cortex have been reported for several conditions in which emotional numbing and/or suppression of emotions are common symptoms. For example, a metaanalysis of functional neuroimaging in posttraumatic stress disorder (PTSD) found reduced activation in both sACC and aMCC during fear conditioning.
44 The authors proposed that reduced activation in sACC may indicate impairment in emotion-regulation and fear-extinction, whereas the reduced activation in aMCC may relate to reduced experience of negative emotion. Patients with schizophrenia, which often includes a flattening of affect, show less aMCC activation during executive tasks.
45–48 A metaanalysis of changes in brain activation in depression reported that more studies found reduced rather than increased activation in response to emotional stimuli in the ACC and MCC.
49 A study of fear-conditioning reported absence of the normal activations (anterior cingulate, insular, and orbitofrontal corticies) in individuals with psychopathy, whereas individuals with social phobia showed an overactivation of these regions.
50 Although results are not entirely consistent across all studies, most anxiety disorders other than PTSD show excessive activation in both the pACC and MCC.
44,51 One possible reason for this is the difference in what is feared. For example, in PTSD, there can be a fear of personal harm, whereas, in social phobia (social anxiety disorder), there is more likely a fear of public embarrassment. Greater-than-normal activation of both pACC and MCC have been reported for individuals with generalized anxiety disorders for tasks involving aversive stimuli, and, for individuals with obsessive-compulsive disorder, during tasks involving errors.
52–56 Longitudinal studies of treatment (e.g., cognitive-behavioral therapy, medication) have found at least partial normalization of brain activation in treatment responders.
49,57,58 Comparison of different treatment approaches indicates both similar and unique alterations in brain activity.