Early postmortem studies suggested that brains from schizophrenic patients were smaller in weight and had a smaller cortical volume relative to normal brains and to brains from patients with affective disorder
(4,
5). Likewise, initial structural analyses of schizophrenic patients with magnetic resonance imaging (MRI) uncovered global deficits in cortical gray matter
(6,
7). More recently, some MRI analyses have found selective or more pronounced deficits of frontal gray matter in schizophrenic brains
(8–
12). However, smaller frontal lobe gray matter volume has not been reported consistently in MRI studies of schizophrenic patients
(13,
14). This inconsistency may be partly attributed to difficulty in defining the posterior border of the frontal lobe. Arbitrary landmarks, such as the genu of the corpus callosum or level of the optic chiasm, have often been used to demarcate the posterior extent of the frontal or prefrontal cortex, although the relationship of these structures to the frontal cortex is highly variable
(15). In addition, MRI analyses frequently rely on automated segmentation programs to define the border between gray and white matter, creating a boundary that is dependent on scanner settings.
Method
A total of 47 brains were examined in this study. Thirty-seven brains were obtained from the Harvard Brain Tissue Resource Center, Belmont, Mass.; 10 brains were obtained from the Section on Neuropathology, Clinical Brain Disorders Branch, Intramural Research Program, National Institute of Mental Health (NIMH), Bethesda, Md. All brains were procured in accordance with procedures established by the institutional review boards of the respective institutes and were screened for neuropathology both macro- and microscopically. Fourteen brains were eliminated from the study for the following reasons: insufficient medical records for a definitive diagnosis (N=7), final diagnosis other than schizophrenia (N=5), secondary diagnosis of multiple sclerosis (N=1), and family history of schizophrenia in a comparison subject (N=1). Demographic data for the remaining 19 normal comparison subjects and 14 schizophrenic patients are shown in
Table 1.
The diagnosis of schizophrenia was ascertained via postmortem, retrospective review of the subjects’ medical records and application of either the Feighner criteria, for the brains from the Harvard Brain Tissue Resource Center (F. Benes, personal communication, 2001), or DSM-IV, for the brains provided by NIMH (J. Kleinman, personal communication, 2001). All subjects with schizophrenia in this study had a chronic illness of at least 6 months’ duration without subsequent return to premorbid levels of psychosomatic adjustment, did not show signs of depression or mania consistent with affective disorder, and exhibited either delusions and/or hallucinations, significant communication deficits related to lack of logical thinking, or both. Under the Feighner criteria, “probable schizophrenia” is distinguished from “schizophrenia” by meeting only two of five of the following demographic criteria, whereas three or more must be met for a diagnosis of schizophrenia: 1) single, 2) poor premorbid social adjustment or work history, 3) family history of schizophrenia, 4) absence of alcoholism or drug abuse within 1 year of onset of psychosis, and 5) onset of illness before age 40. One schizophrenic patient (subject 21) had a secondary diagnosis of alcohol abuse. The brains of two patients were neuroleptic-free (subjects 23 and 27), and the brain of one other patient (subject 20) was essentially neuroleptic-naive, as this subject had received medication for only 1 day.
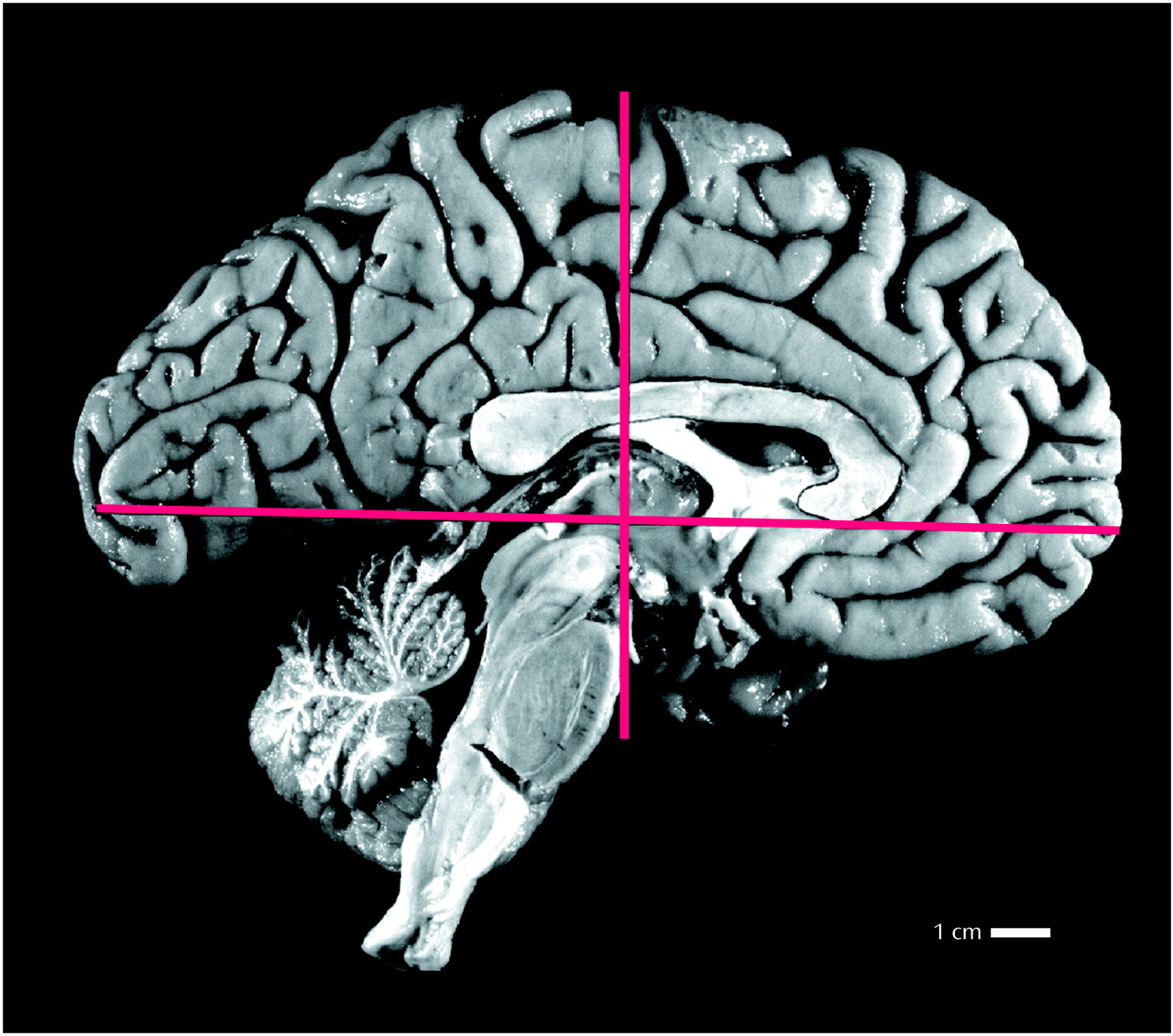
Whole, or nearly whole, formalin-fixed left hemispheres were shipped to the Department of Neurobiology at Yale University School of Medicine. Lateral and medial views of the hemispheres were photographed, and the brains were then dissected into 2-cm-thick coronal slabs by using a custom-made brain slicing apparatus. The first coronal cut was made on a plane perpendicular to the line connecting the anterior and posterior commissures and was positioned at the midpoint of this line (
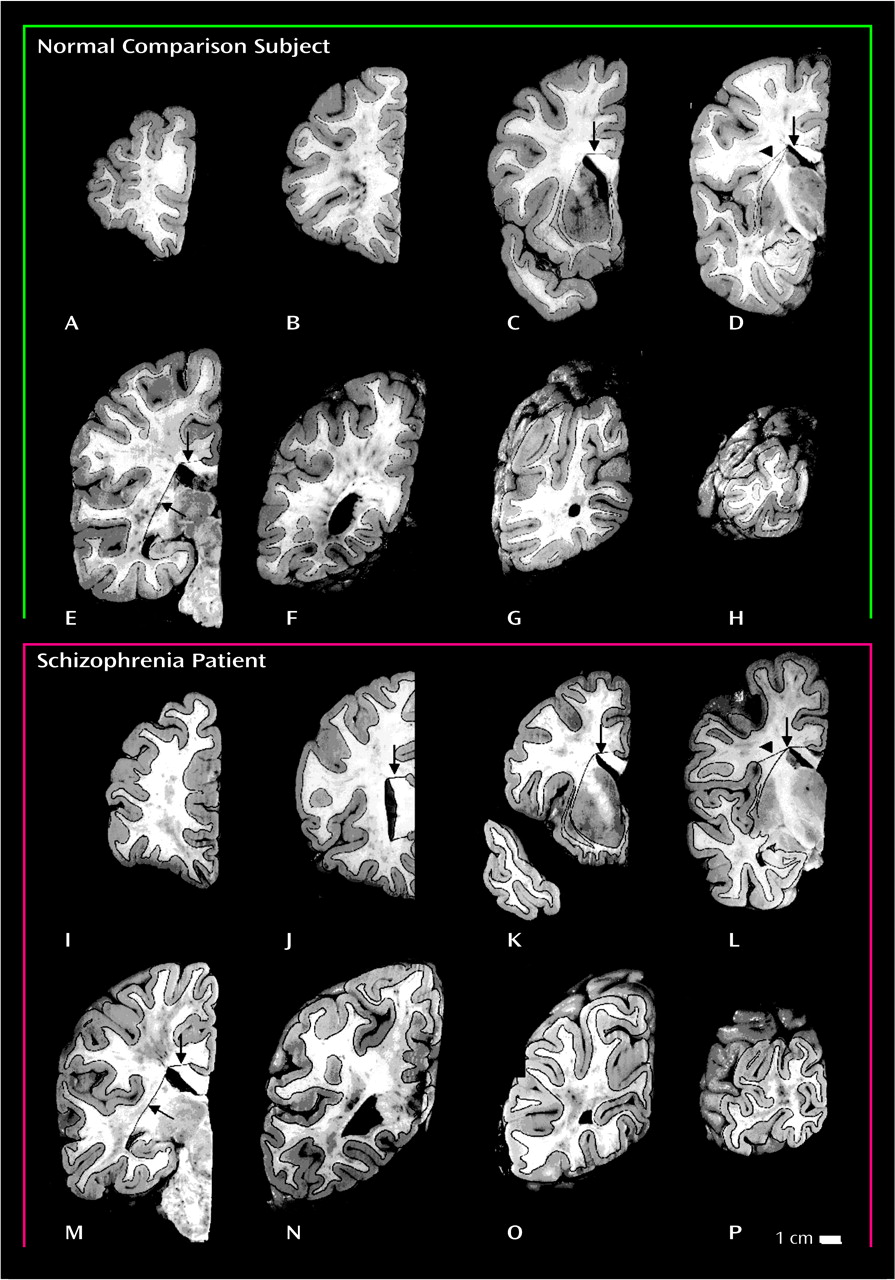
Figure 1). Subsequent cuts were made anterior and posterior to this initial cut at 2-cm intervals. This dissection yielded 8–10 coronal brain slabs. As the posteriormost slab did not have a posterior cut surface, 7–9 slab faces per brain were analyzed. The posterior faces of these slabs were photographed and printed at approximately 2× magnification; gray and white matter compartments were easily distinguished and outlined with a fine black marker (
Figure 2). All photographs were coded so that the investigators who performed the measurements were blind to the subject’s diagnosis. A point-counting grid was placed over each photograph, and the number of points on the grid (spacing=1 cm) falling on gray and white matter was recorded. The Cavalieri method
(19) was used to determine the total gray matter volume and the total white matter volume. Frontal gray matter volume and frontal white matter volume were estimated by using the same procedure but only for slabs that included the mid-commissural cut (
Figure 2, panels D and L) and tissue anterior to the mid-commissural cut. Five brains (one from a comparison subject and four from schizophrenic patients) were missing small areas of the medial temporal cortices; therefore, only measurements of frontal gray matter volume and frontal white matter volume were performed for those brains. The coefficient of error, calculated by dividing the standard error of the mean by the mean
(19), for all measurements in this study ranged from 0.032 to 0.043.
Measurement reliability was assessed by using a subset of four brains (two normal, two schizophrenic) for total gray matter volume, total white matter volume, frontal gray matter volume, and frontal white matter volume. The Pearson correlation coefficients for the four volumes were 0.99, 1.00, 0.97, and 0.78, respectively. Two-tailed significance values for the correlations (N=4) were <0.02, 0.003, <0.03, and 0.22, respectively. Similar results were obtained with Kendall’s tau-B and Spearman’s rho.
A Student’s t test was performed after Pearson correlation to determine whether the normal comparison group and the schizophrenic group differed in age, postmortem interval, brain storage time in formalin, or brain weight. In addition, regression analyses were performed to assess possible correlations between measured parameters and age, postmortem interval, storage time in formalin, or brain weight. Significant correlations were found between brain weight and total gray matter volume (r=0.69, t=–3.93, df=27, p=0.001), total white matter volume (r=0.57, t=2.80, df=27, p=0.01), frontal gray matter volume (r=0.58, t=3.70, df=32, p=0.001), and frontal white matter volume (r=0.61, t=3.84, df=32, p=0.001), and between storage time in formalin and total gray matter volume (r=–0.61, t=–3.93, df=27, p=0.001), total white matter volume (r=–0.52, df=27, p=0.01), frontal gray matter volume (r=–0.45, t=–3.08, df=32, p=0.005), and frontal white matter volume (r=–0.39, t=–2.65, df=32, p<0.02). Accordingly, an analysis of covariance (ANCOVA) was performed on these volumes, with diagnosis as the factor and brain weight and storage time in formalin as covariates. Possible gender differences in brain weight, total gray matter volume, total white matter volume, frontal gray matter volume, and frontal white matter volume were analyzed with ANCOVA, with storage time in formalin as the covariate. Since the study group included some brain samples with incomplete hemispheres, volumes for each region were analyzed by independent ANCOVAs. As an additional control, volumes for each region for the study group subset with complete hemispheres were analyzed by repeated measures ANCOVA. Data were initially analyzed with gender as a factor. However, observed power estimates were low. Moreover, nearly all of the gender-based variance was accounted for by using brain weight as a covariate. As a consequence, except for the gross overview noted below, gender differences were not considered in this study. For all statistical analyses, p<0.01 was considered significant, although 0.1<p<0.01 are also reported.
Discussion
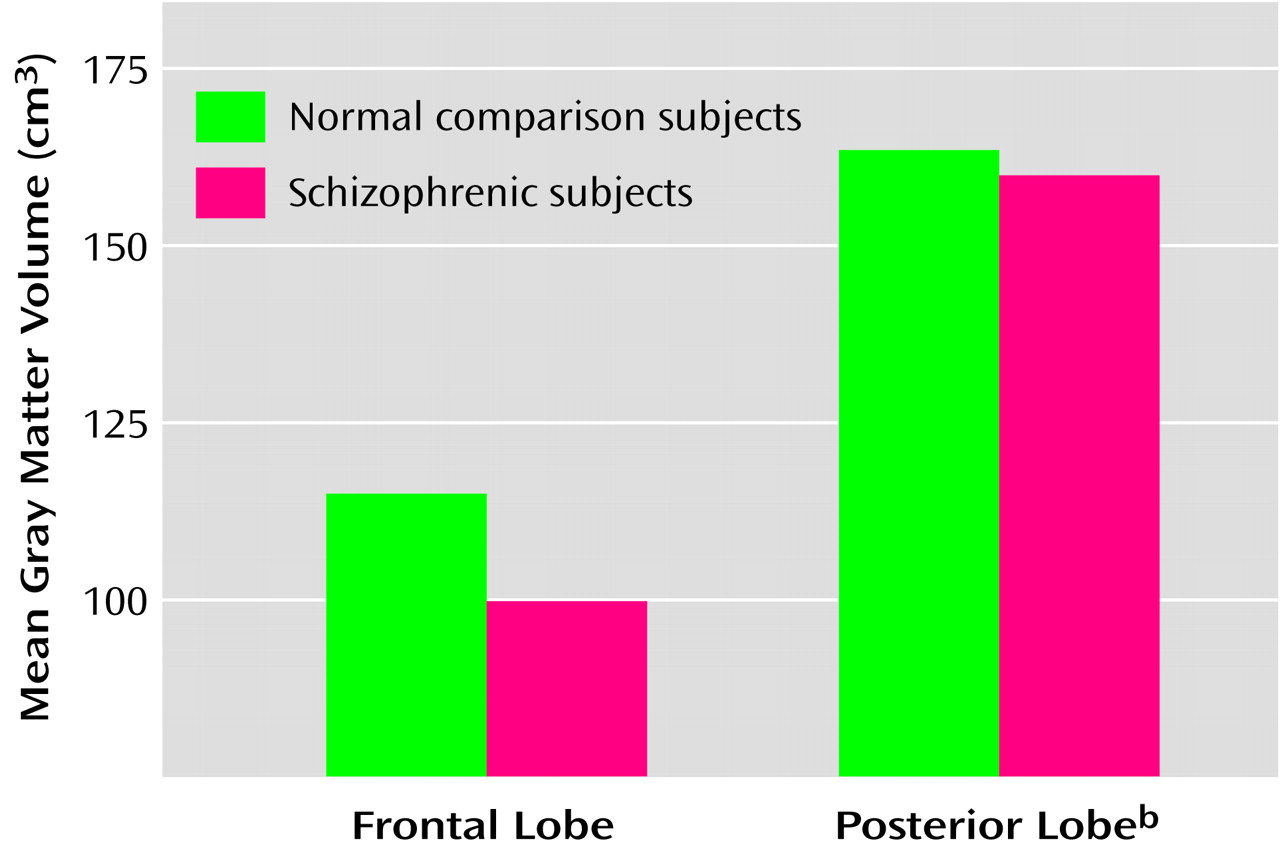
In the full group of brains in this study, only the frontal gray matter volume was significantly smaller (12% smaller) in the schizophrenic subjects than in the comparison subjects. Total gray matter volume of the schizophrenic brains was 8% smaller than that of the comparison brains, but this difference was not significant. However, with a larger number of subjects, a significant difference in whole gray matter volume may have been detected. These observations are consistent with the findings of several MRI studies indicating a preferential or selective deficit in gray matter volume in the frontal lobes
(8–
12) and with those of recent MRI analyses focusing specifically on subdivisions of the prefrontal cortex
(20–
22). However, smaller frontal gray matter volume is far from a universal finding in schizophrenia research. Indeed, only about half the MRI studies have reported smaller frontal gray matter volume in schizophrenia
(13,
14). McCarley et al.
(14) suggested that the 8% smaller prefrontal cortical width we previously described
(23) is near the limit that can be detected with the resolution of current MRI technology, resulting in both positive and negative reports in the literature. As schizophrenia is a heterogeneous disease, cohort differences certainly might also contribute to the inconsistency between studies. Finally, imprecision in the definition of the frontal lobe may account for some of the variability in findings. Arbitrary anatomical landmarks, such as the corpus callosum or optic chiasm, have often been used to determine the posterior border of the frontal lobe. In this study, we elected to use the midpoint of the anterior commissure–posterior commissure line to define the posterior margin of the frontal lobe, a delineation that has been shown to represent an accurate approximation of the central sulcus
(15). In addition, the border between gray and white matter, a boundary that is dependent on scanner settings with MRI, is readily discernible on slabs of formalin-fixed brain tissue. Thus, accurate discrimination of the frontal lobe gray matter compartment in a cohort that did not include elderly subjects (>76 years old) may have enabled us to detect relatively small volumetric differences in schizophrenic and comparison brains.
Comparison With Previous Postmortem Studies
Previous postmortem studies of the whole frontal lobe
(16), prefrontal gray matter
(17), or frontal gray matter
(18) have not reported significantly smaller volumes in schizophrenic brains, although small numbers of subjects
(16,
17), inclusion of aged subjects
(17,
18), and inclusion of both brain hemispheres
(16,
17) may account for the negative findings. Brain volume has been shown to undergo significant age-related reduction, with the frontal lobes exhibiting more severe volumetric deficits with age than the other cortical lobes
(24–
26). Moreover, aging is not uniform across individuals. Aged rats have been shown to exhibit functional and anatomic heterogeneity. Some aged rats do not differ from young rats with respect to hippocampal morphology or function, while others show deficits in learning capacity and concomitant changes in hippocampal volume
(27). In humans, dendritic patterns in aged subjects show similar variability
(28). Therefore, simple age-matching does not compensate for the changes occurring in elderly subjects. A recent study found that the age-related changes in frontal lobe volume were indistinguishable from the pathology found in schizophrenic subjects
(29). As a result, inclusion of aged comparison subjects is likely to mask volumetric findings in the schizophrenic cohort or, at the very least, to increase the variability in both comparison and schizophrenic groups, making it less likely that differences will be detected.
Despite differences between our findings and earlier findings with respect to schizophrenic patients, our volumetric measurements for the comparison brains are very similar to those found in previous postmortem studies that used sulcal landmarks to define the boundaries of the frontal lobe
(16–
18). For instance, Pakkenberg
(16) reported a frontal lobe volume of 211.6 cm
3, representing gray and white matter volumes combined, in normal comparison subjects. A later study of normal human brains
(30) reported a frontal lobe volume of 213 cm
3. The combined frontal gray and white matter volumes for the comparison subjects in the present study totaled 210.5 cm
3. Although a somewhat smaller frontal lobe volume (174.4 cm
3) was reported by Highley et al.
(18), the cingulate gyrus was not included in their definition of the frontal lobe. Thus, the agreement between the frontal lobe volume measured in our study and that of previous reports using sulcal landmarks is quite good. The volume of comparison subjects’
prefrontal gray matter volume reported by Thune et al.
(17) (62.5 cm
3) is significantly smaller than the volume we found for whole frontal lobe gray matter (114.7 cm
3), but that should be expected.
Technical Considerations
We chose to use the definition of Talairach and Tournoux
(15) for the posterior border of the frontal lobe for two reasons. First, we wanted to dissect the brains as uniformly as possible, and this was best achieved by making the first cut at the midpoint of the anterior commissure–posterior commissure line. In addition to rendering brain slabs that were cut at a standard angle, our protocol also generated a random first cut. If we had begun cutting from the frontal pole, the first cut would not have been random. Second, once the brains were cut, identification of the central sulcus would have been difficult, because the central sulcus, like the plane of dissection, is oriented in the coronal plane and basically traverses the posterior face of the block just anterior to the first cut. It should be noted that the Sylvian fissure was used to define the boundary between the frontal and temporal cortices; therefore, while it might be possible that some parietal cortex was included in delineation of the frontal lobe, our definition of frontal lobe absolutely did not include any anterior temporal cortices.
Possible Confounding Factors
Storage time in formalin was the only factor that was significantly correlated with gray and white matter volumes. These correlations probably reflect differences in storage time between the Harvard Brain Tissue Resource Center and NIMH cohorts, as storage time in formalin was longer in the NIMH brains, averaging a little more than 4 years. Nevertheless, storage time in formalin was included as a covariate in the ANCOVAs for these measures, and the resulting analysis showed that the selective deficit in frontal gray matter volume remained significant.
The slightly longer postmortem intervals in the schizophrenic group relative to the comparison group also probably cannot account for the differences in frontal gray matter volume observed in this study, because the deficit in the entire schizophrenic group was also found in the smaller subgroup of whole brains, even though the schizophrenic and comparison groups had no difference in postmortem interval. Moreover, postmortem interval did not show a significant correlation with any of the measured parameters in the regression analysis.
Most of the schizophrenic patients in this study had received typical antipsychotic drugs. Changes in the volume of subcortical structures have previously been shown to be related to medication
(31–
33). Therefore, the possibility that long-term exposure to antipsychotic drugs may account for the smaller volume of cortical gray matter must be considered. However, the findings for the three schizophrenic patients in this study who were essentially neuroleptic-free revealed the same selective frontal gray matter volume deficit (12%) that was found for the schizophrenic group as a whole. Likewise, smaller gray matter volumes in specific regions of the prefrontal cortex have been found in neuroleptic-naive patients
(21), indicating that the deficit in gray matter volume is a pathologic correlate of the disease and not a medication effect. Moreover, cortical gray matter was not reduced in thickness in nonhuman primates treated with antipsychotic drugs for a period of 6 months
(34).
Exclusion of the schizophrenic patient with a secondary diagnosis of alcohol abuse did not alter the results of the study: frontal gray matter volume was still significantly smaller in the schizophrenic group without this patient (13% smaller than in the comparison group).
Prominence of Frontal Lobe Pathology
The study results indicate that the difference in gray matter volume between schizophrenic and normal brains was greater in the frontal lobe than in more posterior lobes. While these findings lend further support to several MRI studies that have indicated a preferential or selective deficit in gray matter volume in the frontal lobes
(8–
12), it is noteworthy that circumscribed regions of the mesial temporal cortex
(35–
39) and superior temporal gyrus
(40–
43) in schizophrenic patients have been shown to exhibit smaller volumes that are comparable in extent to the frontal lobe deficit reported here. Thus, a finer-grained analysis of volume in the posterior cortices may have revealed small regional deficits in gray matter volume of the temporal or parietal lobes that would rival the reductions observed in the frontal lobe as a whole. Nonetheless, it is an important observation that the magnitude of the volumetric deficits in gray matter volume mirror the functional deficits to a large extent. For example, cognitive deficits on “prefrontal” tasks that involve abstract thinking, working memory, and contextually dependent behaviors are more pronounced in schizophrenia than are deficits in visual processing
(3), thereby correlating with the greater reduction of frontal than occipital lobe gray matter. Indeed, if we had been able to delineate the prefrontal cortex in our analysis, the deficit in schizophrenic patients might have been more pronounced.
Data from the present study support a growing literature suggesting that the frontal lobe, and the prefrontal cortex in particular, is a focal point for pathology in schizophrenia. Although few in number, postmortem studies that have examined both prefrontal and more posterior cortices in schizophrenic brains have generally found specific prefrontal deficits
(44,
45) or more marked pathology in the prefrontal cortex
(23). However, postmortem assay of excitatory amino acid concentrations and the levels of related enzymes have found abnormalities in hippocampal and prefrontal, but not parietal or cingulate, cortices
(46). Paired temporal and frontal pathology has also been observed with proton magnetic resonance spectroscopy of
N-acetylaspartate concentrations in the schizophrenic brain, revealing bilateral reductions of
N-acetylaspartate in the hippocampus and dorsolateral prefrontal cortex
(47). Thus, the cortical pathology in schizophrenia is nonuniform and seems to selectively involve prefrontal and medial temporal cortices.
Finally, the loss of prefrontal gray matter is consistent with the now substantial evidence for selective changes in prefrontal cell morphology or function rather than a loss of cell number per se
(3,
48). Thus, although we cannot exclude the possibility that small pockets of neuronal degeneration are present in the cerebral cortex, the volumetric deficit in the frontal lobe most likely represents a smaller than normal number of the structures associated with neuronal connectivity, i.e., axonal processes, synaptic terminals, and dendritic arbors and spines.