Modern psychiatry has consistently stressed the seminal role of early childhood experiences in shaping adult behavior. It must, however, be emphasized that a child does not develop in a psychological vacuum. Freud’s Oedipal conflict
(1), Bowlby’s concepts regarding attachment
(2), and Klein’s notion of infant splitting of the mother
(3,
4) all suggest that parent-child interaction is critical to understanding the developing human psyche. The ongoing social debate regarding the adequacy of single parenting further underscores our societal conviction that good parenting, however defined, is crucial to optimal child development.
The children of depressed parents unfortunately are often ill-adapted to meet the myriad challenges posed by the modern world. They are more likely to experience psychiatric illness
(5–
9), with depressed adolescents being three times more likely than nondepressed adolescents to have had a depressed parent
(10). Furthermore, the children of depressed parents experienced greater social, educational, behavioral, and vocational difficulties
(7,
11,
12). Although this familial risk is in part genetically transmitted, other potential pathways of transmission include: 1) exposure to prenatal maternal neuroendocrine aberrations associated with depression during pregnancy, 2) exposure to heightened intrafamilial stress when a parent is depressed, and 3) exposure to the affective, behavioral, and cognitive alterations of the depressed parent
(13).
Considerable attention has been focused on the impact of depression on parenting behavior. For example, a recent meta-analysis of 46 observational studies of depressed mothers demonstrated a moderate association of maternal depression with negative (i.e., hostile, coercive) parenting behaviors and disengaged parenting behaviors
(14). Despite social changes leading to increased paternal participation in child rearing, there has been little investigation of the impact of depression on the parenting behaviors of fathers. One recent study did indicate that families with a depressed father are less likely to sustain positive communication (e.g., approval, assent, humor) than families with a depressed mother
(15). Conversely, another study suggested that depressed fathers are less likely than depressed mothers to engage in negative parenting behaviors; however, it remains unclear whether this is a consequence of gender-specific differences in the manner of coping with depressive illness or the less taxing demands of parenting placed on fathers, who more commonly serve as secondary caregivers
(16).
Alterations in the parenting behaviors of depressed parents may in turn adversely impact the psychosocial and neurobiological development of their children. A meta-analysis of the association between maternal mental health and the security of infant attachment, encompassing 35 studies and more than 200 mother-infant dyads, concluded that maternal depression, maternal stress, and an unsupportive partner are all associated with a greater risk of insecure infant attachment
(17). The authors noted, however, that the heterogeneous distribution of the observed effects suggested that other as yet unidentified contextual factors help determine the probability that the child of a depressed mother develops an insecure attachment.
A few studies have demonstrated that neurobiological alterations are also evident in the children of depressed parents. For example, in two recent studies the preschool-age children of depressed mothers exhibited electroencephalographic alterations in frontal lobe activity that correlated with diminished empathy and other behavioral problems
(18,
19). Neuroendocrine function may also be altered, as one study demonstrated higher serum cortisol concentration in the children of depressed mothers that was correlated with the severity of maternal depression
(20).
Maternal depression during pregnancy also has potentially deleterious consequences for infant development. The newborns of mothers depressed during pregnancy have exhibited poorer motor skills during neurological examination
(21). In addition, depression-associated changes in maternal hypothalamic-pituitary-adrenal (HPA) axis function during pregnancy have been shown to be predictive of similar biological alterations in newborn offspring
(22). At 6 months of age, the children of euthymic mothers who had been depressed during pregnancy continued to exhibit heightened salivary cortisol responses to a standardized psychosocial stressor
(23).
In summary, parental depression may represent a significant early adverse life event for the developing child. Improving our understanding of the mechanisms whereby parental depression conveys risk to the child is critical to the development of therapeutic guidelines for depressed parents. The progress of this research is hampered by the understandable ethical and logistical complexities of conducting neurobiological research involving at-risk children, but the extant preclinical literature contains a wealth of information, unfamiliar to many clinical psychiatrists, that may hasten our understanding of these processes. A variety of animal research models have been devised to investigate the impact of early life stress on development. Although early animal models of stress have most often been cited in the psychiatric literature for their similarities to the impact of childhood trauma (e.g., child abuse, exposure to war), a subset of these research models conferred stress on the young animal by interfering with parental care. Such animal models consequently may be relevant to a consideration of human conditions such as depression that hinder parenting efforts. This review summarizes the laboratory animal data paralleling the common clinical dilemma of parental depression and its potentially deleterious impact on child development.
Defining and Validating Animal Models of Human Behavior
The continuum of animal models used in psychiatric research ranges from animal assays to homologous models
(24,
25). Animal assays provide an expeditious preclinical means to screen potential psychotropic medicines by instituting specific behavioral assessments after the administration of a candidate compound. Because the target animal behaviors need not be similar to any human parallel, animal assays have limited utility for the study of disease etiopathogenesis. Homologous models are instead predicated on the conviction that certain aspects of animal behavior and physiology mirror their human counterparts by virtue of a common evolutionary ancestry. The animal behaviors of interest are not simply analogous (i.e., mimicking a human behavior) but homologous (i.e., resembling a human behavior because they serve a similar function and are activated by similar biological processes). Homologous models therefore permit testing of hypotheses regarding human behavior at various levels within the phylogenetic order.
In a homologous model, the test animal is exposed to a laboratory stimulus that reliably induces a state approximating one or more attributes of some human condition. The stimulus that induces the homologous state may be either a biological intervention (e.g., pharmacological probe, surgical procedure) or an environmental manipulation (e.g., thermal extreme, psychosocial deprivation). When an environmental stressor is used as the laboratory stimulus, the similarity of both the stressor and its outcomes are critical to any valid application of the model to an understanding of human behavior.
Numerous sets of criteria have been developed for evaluating animal models. In one of the simplest and most widely referenced, McKinney and Bunney
(26) contended that the validity of an animal model is contingent on its reproducibility (i.e., reliability) and its accuracy in portraying the phenomenology and treatment response of a corresponding human illness. In the context of psychiatric research, in which the nosology is predicated not on disease causality but on a descriptive phenomenology of illness, this emphasis on empirical evidence has proven most welcome.
More stringent criteria proposed by Willner
(27) require that animal models exhibit face, predictive, and construct validity. Face validity refers to the phenomenological similarity between an animal model and a human syndrome. It should be remembered that behaviors may be analogous without being homologous. For example, certain avian species commonly kill their young for brood reduction during periods of nutritional stress. Although this avian behavior bears certain empirical similarities to human child abuse, few would argue that it is a relevant model for studying human infanticide. Predictive validity refers to the accuracy of a model in forecasting the course and outcome of a human syndrome. It has been argued that predictive validity is the only scientifically meaningful criterion by which to evaluate an animal model
(25). Finally, construct validity represents the degree to which both the human syndrome and the animal model are unambiguously defined such that a rational theory can be constructed to explain and test theories regarding the etiopathogenesis of the disorder. It is ultimately the presence of homology that constitutes the construct validity of a purportedly homologous animal model.
Although there is widespread agreement that construct validity constitutes the defining property of a test, neither psychosocial nor neurobiological research has successfully met Willner’s call for construct validity. Because the pathophysiology of mental disorders remains obscure, the homology of an animal model to a human psychiatric condition cannot be absolutely demonstrated. An animal model’s homology can nevertheless be inferred, in the absence of a conclusive determination of construct validity, when evidence from the model’s face and predictive validity is consistent with a rational theoretical construct explaining how the model reflects a human condition.
Parenting and the Diathesis-Stress Model
Stress plays a pivotal role in the pathogenesis of many psychiatric
(28) and medical illnesses
(29–
32). Individuals, however, display a heterogeneous susceptibility to the effects of stress. Whereas some are especially resilient, others demonstrate an endogenous vulnerability toward stress-induced illness. It is this predisposition to illness, or diathesis, that provides the foundation for the diathesis-stress model
(33–
37).
The diathesis-stress model has been applied to a host of psychiatric and medical illnesses. This model has also been applied to our understanding of parenting styles
(38) and to the development of aberrant parenting behaviors
(39). But what is the source of the diathesis? One contentious area of deliberation has been the relative contribution of inheritance and environment, the so-called nature-versus-nurture debate. Such arguments have historically been couched in absolute terms, but the diathesis-stress model permits a more balanced consideration. Because inherited and acquired factors contribute to vulnerability, this model becomes a useful structure for organizing concepts regarding the mechanisms by which parents convey the risk for illness to their offspring. The parental contribution to the vulnerability of offspring to psychiatric illness may occur through numerous pathways, most notably genetic transmission and altered parenting. Theoretically, the predisposition to illness should be demonstrable by using both psychological and biological measures. It is this conviction that the diathesis for illness may be measurable that underlies many applications of animal models to the study of human behavior.
Despite the absence of a definitive conclusion regarding their validity, homologous animal models do offer a means to explore the diathesis-stress model. Both inherited and acquired vulnerability to experimentally induced stress have been demonstrated by using animal models. The genetic diathesis has largely been demonstrated not by design but inadvertently as researchers have endeavored through selective breeding to develop animal strains that either more reliably or more robustly exhibit the target signs and symptoms resembling a human syndrome. The success, for example, in breeding animals that are more susceptible to stress-related behavioral alterations illustrates the contribution of genetic inheritance to the vulnerability to stress-related illness.
Animal models have also contributed to our understanding of environmentally acquired vulnerability to disease. Stress sensitization models have been used to demonstrate that exposure to one stressor can alter the response to subsequent milder stress exposures. For example, in a predator exposure model, rats placed briefly in a small enclosure with a cat later demonstrated increased anxiety-like behaviors that were detectable long after the initial exposure
(40,
41). Similarly, animals exposed to a foot shock and then repeatedly introduced to the setting of that initial stressor developed an exaggerated acoustic startle
(42). One frequent application of animal models relevant to the diathesis-stress model has been to study the biobehavioral sequelae of early life stress. In some instances, these models simply expose young animals to nonspecific noxious stimuli such as foot shock or restraint. Models of early adverse experiences have recently evolved to utilize psychosocial stressors. These models typically stress the young animal by modulating parent-offspring interaction and thereby reflect not only early life stress but more specifically conditions such as parental depression that hinder parenting.
In assessing the validity of such animal models, it is critical to establish an operational definition of the comparative human experience. As mentioned previously, the homology applies not only to parallels in early life stress but also to long-term outcome. The most extensively studied psychiatric vulnerability arising in the children of depressed parents has been for subsequent depression. Hence, the predictive validity of homologous animal models of parental depression ultimately depends on the accuracy of these models in recreating the persistent biobehavioral changes underlying a diathesis for depression.
Regarding the neurobiology of depression, existing research has emphasized monoamine neurotransmitter systems and the HPA axis. The classic HPA axis stress response commences with increases in hypothalamic corticotropin-releasing hormone (CRH) secretion from nerve terminals in the median eminence that in turn induce pituitary release of ACTH and ultimately increased adrenal secretion of cortisol. The role of HPA axis alterations in depression has been studied for more than 40 years, since it was first reported that depressed patients have increased concentrations of circulating adrenocortical hormones
(43,
44). Cortisol nonsuppression in the dexamethasone suppression test reliably demonstrates an association between severity of depression and HPA axis hyperactivity, although clinical utility of such findings is limited because of poor sensitivity and specificity
(45–
48). Numerous lines of evidence, including elevated concentrations of CRH in CSF
(49–
53), blunted ACTH responses to administration of exogenous CRH
(54–
58), and heightened CRH mRNA expression in the hypothalamus
(59), have indicated that CRH hypersecretion likely underlies the pathogenesis of HPA axis derangements in depression. More illuminating is the finding that CRH neurons reside in extrahypothalamic regions of the central nervous system (CNS) and may mediate the neurovegetative symptoms of depression via limbic projections
(60). Though the neurocircuitry of central CRH alterations during depression remains obscure, it can now be stated with confidence that depression is commonly associated with both CRH hypersecretion and hypercortisolemia, with the added aberration of dysfunctional HPA axis negative feedback
(36,
60–63).
Monoamine neurotransmitters, particularly norepinephrine and serotonin (5-HT), have likewise garnered considerable scrutiny in studies of the neurobiology of depression. Noradrenergic systems, once postulated to be central to the pathophysiology of depression
(64), have produced largely inconsistent results in the intervening years. Long-standing psychophysiological research has nonetheless repeatedly demonstrated physical responses to stress suggestive of sympathetic activation, including tachycardia, hypertension, diaphoresis, dizziness, heightened skin temperature and skin conductance, and increased electromyographic activity. It now appears unlikely that alterations in central norepinephrine activity are necessary to the pathophysiology of depression; however, there remains substantial evidence that dysregulation of locus ceruleus norepinephrine systems contributes to the elaboration of numerous depressive symptoms
(65). Some theories have implicated a central role for 5-HT dysfunction in depression
(66). The widespread utility of serotonergic antidepressants lends considerable credence to such hypotheses. The identification of numerous anatomical and functional interfaces between CRH and 5-HT systems in the CNS
(36,
67) has provided a theoretical framework for understanding the neurobiology of depression that is only now beginning to be explored.
It must be acknowledged that the psychophysiological and peripheral neuroendocrine correlates of depression provide no direct explanation of disease etiopathogenesis. Such measures have nonetheless been useful because they use minimally invasive procedures to obtain a peripheral window into higher cortical functioning
(63,
68). Consequently, studies of HPA axis dysfunction do not directly explicate the pathophysiology of depression but nevertheless offer important insights leading the elucidation of comprehensive theories regarding the primary role of the dysfunction of cortical CRH systems
(60,
69,
70).
Comparing the biobehavioral similarities of animal studies to parallel indices in human depression research thus becomes the basis for assessing the valid application of animal investigations to the consequences of parental depression on child development. Our focus is therefore on identifying animal behaviors and peripheral neurobiological alterations that resemble the current clinical findings in depression and the children of depressed parents.
Homologous Models of Parental Depression and Child Development
Neonatal Stress Models
Noxious stimuli
One approach to studying early life stress by using animal models has been to implement stressors akin to those experienced by adult animals. In so doing, researchers expose young animals to noxious stimuli, including thermal extremes, pinprick, foot shock, or surgical procedures.
Despite clear evidence of the presence of CRH-containing neurons in the fetal rat
(71), noxious stimuli such as these evoke only a subnormal HPA axis response during the first 2 weeks of life
(72,
73). During this so-called stress hyporesponsive period, baseline plasma corticosterone concentrations are lower than normal
(74) and are only minimally increased by exposure to a noxious stressor
(74–
76). Stress-induced CRH and ACTH responses are also attenuated during the stress hyporesponsive period
(77–
79). The HPA axis is not a static but a dynamic system that at least in the rat continues to undergo maturational adaptations postnatally. The failure to mount a full stress response during early development may increase the young rat’s vulnerability, but this may serve as an adaptive evolutionary trade-off, protecting the growing animal from the catabolic effects of glucocorticoid hypersecretion. There is no evidence that a similar stress hyporesponsive period exists in the human neonate. This may, of course, be related to the fact that the CNS of the newborn rat pup is developmentally similar to that of a 24-week human fetus
(80). Thus, if a stress hyporesponsive period exists in humans, it may occur prenatally.
Although studies utilizing the exposure of neonates to noxious stimuli have limited face validity in addressing the issue of parenting, and in fact even fail to provide predictive validity for the expected neurobiological stress responses, these studies do illustrate the dynamic nature of the stress response. If HPA axis responsivity during childhood is modulated by programmed maturational processes, then this stress-responsive biological system may serve as a template for the environmental contribution of early life stress on the diathesis for subsequent illness.
Brief neonatal handling in rodents
Another applicable line of animal research has used brief neonatal handling of rodents as the initial stressor. In this paradigm, rodent pups are removed daily from their cages and handled briefly by laboratory personnel. The biobehavioral responses of handled and nonhandled pups to postweaning stress are then examined to determine the persistent effects of the handling exposure.
The expectation that neonatal handling would increase anxiety and biological measures of stress responsivity in future novel situations has not been realized. In fact, neonatal handling has been found paradoxically to decrease anxiety-like behaviors and biological stress responsivity. Handled pups are resistant to stress and aging decrements in learning ability
(81,
82), exhibit higher levels of exploratory behavior in novel environments
(83,
84), and demonstrate an attenuated activation of neuronal fear circuitry in response to stress during adulthood
(85).
Neonatal handling produces transient norepinephrine circuit hyperactivity
(86) but surprisingly leads to a decreased norepinephrine response to stress during adulthood that appears to be a consequence of an up-regulation of α
2 autoreceptors in the locus ceruleus
(87). This paradigm also produces persistent increases in the CNS activity of γ-aminobutyric acid (GABA)
(88), 5-HT
(89,
90), and endogenous opioids
(91–
94). Undoubtedly, however, the most intriguing findings are decreases in HPA axis activity induced by neonatal handling. Early studies demonstrated lower ACTH
(95) and corticosterone
(96,
97) responses to subsequent stressors. Neonatal handling also produces reductions in basal circulating levels of ACTH and corticosterone
(98). These diminished peripheral measures of HPA axis activity are, at least in part, a consequence of decreased CRH secretion as evidenced by lower median eminence concentrations of CRH, decreased expression of hypothalamic CRH mRNA
(95,
99), and diminished CRH release in response to subsequent stress
(99,
100).
Two lines of evidence indicate that brief neonatal handling may also induce an increased sensitivity to HPA axis negative feedback. Specifically, handled pups demonstrated enhanced dexamethasone suppression of corticosterone and a more rapid return of corticosterone to baseline measures after stress exposure
(101). This enhanced sensitivity to negative feedback is likely a consequence of glucocorticoid receptor up-regulation at the hippocampus and other sites
(101–
105).
Neonatal handling studies have indicated that subtle environmental variations can affect the ontogeny of biological systems but on first review may not be directly applicable to a study of the impact of stress on parenting. How are we to understand the effects of neonatal handling? In fact, neonatal handling so improves the rat’s adaptability to subsequent stress that one study implemented “nonhandling” as an early life stressor
(106). One potential explanation is that rodent pups do not experience handling as a particularly unpleasant stimulus. Although handling introduces an environmental change, it is not empirically noxious. Early exposure to novel nonnoxious environmental events such as handling may instead improve the animal’s adaptability to subsequent environmental changes. These studies may simply represent a variant of other early life stress paradigms and are otherwise irrelevant to our consideration of parental depression.
However, it has been observed that neonatal handling of rodent pups stimulates maternal grooming of pups
(97,
107–110). The biobehavioral changes induced by neonatal handling may thus be mediated indirectly through changes in maternal care. On closer inspection, neonatal handling studies may in fact be relevant to the study of parenting in humans. This hypothesis has been supported by work that has documented that increased maternal grooming (apart from a handling paradigm) produces similar down-regulation of HPA axis responsivity to stress
(111) and, in fact, increases hippocampal synaptogenesis
(112). However, although increased maternal care may be one mechanism by which handling reduces HPA axis responsivity, it may not be the only or even a necessary mechanism for the oft-reported handling effect
(113). In summary, with respect to parenting, the rodent neonatal handling paradigm bears face and predictive validity not for parental depression but for the beneficial effects of attentive parenting.
Maternal deprivation in rodents
In contrast to the previous paradigms, maternal deprivation protocols represent overt models of disrupted parenting. In these models, rat pups are separated from their mother for prescribed interval(s) berfore weaning. The pups are not only deprived of maternal care during the separation, but maternal behavior typically remains aberrant even after reunion
(114), thereby exhibiting greater face validity in relation to the behavior of depressed parents than the earlier paradigms. This parallel to the human condition is underscored by Bowlby’s early application of the term “maternal deprivation” to the impact of mental illness on maternal behavior
(115).
Maternal separation in the rat potentiated behavioral changes resembling depression and anxiety
(116,
117). Maternally separated female offspring were also more likely to display aberrant maternal care with their own litters
(118,
119), although not all studies have demonstrated this finding
(120). Furthermore, maternal separation induced acute changes in HPA axis activity, as indicated by increases in serum corticosterone
(121,
122) and ACTH concentrations
(79). CRH secretion was also altered by maternal deprivation, but the pattern of alteration depended to some extent on the timing of the separation. For example, maternal separation at postnatal day 10 resulted in decreased CRH concentrations in the median eminence but no change in the density of pituitary CRH receptors
(123). The decrease in median eminence CRH was presumably due to depletion of CRH stores from nerve terminals at that site, and the absence of any alteration in pituitary CRH receptors could be attributable to the immaturity of that system. By postnatal day 18, maternal separation produced a down-regulation of pituitary CRH receptors (presumably a consequence of CRH hypersecretion) associated with no changes in CRH concentrations in the median eminence, presumably a consequence of the further maturation of the system resulting in an increased rate of CRH synthesis
(123).
The neurobiological changes induced by maternal deprivation are also long lasting, but the timing of the separation appears to modify the specific biological sequelae. For example, repeated separation of 4–6 hours per day on postnatal days 6–20 produced increased ACTH responses but normal corticosterone responses to subsequent stress in adulthood
(124). By contrast, a single 24-hour separation beginning on postnatal day 3 produced a normal ACTH response but an increased corticosterone response to subsequent stress. Separation beginning on postnatal day 11 induced a normal corticosterone but a blunted ACTH response to stress
(125,
126). Taken in isolation, the specific significance of these findings may be unclear, but overall they demonstrate that interruption in maternal care during a period of rapid CNS development may produce lasting neurobiological alterations.
CRH hypersecretion likely underlies the persistent HPA axis alterations associated with early maternal deprivation. Maternally deprived adult rats exhibited increases in hypothalamic expression of CRH mRNA in the paraventricular nucleus, increased CRH concentrations in the median eminence, and dexamethasone nonsuppression
(124–
127). Diminished elaboration of glucocorticoid receptors in the CNS of maternally deprived rats may underlie alterations not only within the HPA axis but also within noradrenergic systems emanating from the locus ceruleus
(128).
One rat study compared the impact of maternal deprivation on two rat strains: one bred for susceptibility to the inescapable shock (“learned helplessness”) paradigm and another bred for resistance to inescapable shock
(129). When the two rat strains were exposed to maternal deprivation and a subsequent stressor, HPA axis stress responses in the offspring were markedly dissimilar. The stress-susceptible group exhibited an increased ACTH response, but a normal cortisol response, to stress at postnatal day 21, a pattern similar to the HPA axis changes associated with posttraumatic stress disorder (PTSD)
(130–
132). The stress-resistant line did not demonstrate the exaggerated ACTH response to stress. This study provided an excellent example of the potential relative genetic and acquired contributions to the vulnerability for illness.
The genesis of the effects of maternal deprivation on HPA axis activity has been the subject of some debate. The stress experienced by the maternally deprived rat pup may result from either the absence of maternal tactile stimulation or secondary food deprivation
(133). A study designed to address this issue isolated the effects of food intake and tactile stimulation
(134). Providing surrogate maternal care (e.g., stroking the anogenital region with a warm brush) reversed the effects of maternal separation on CRH type 2 receptor mRNA expression in the hypothalamus. However, neither food supplementation nor contact with a sedated or nonlactating mother reversed this effect, suggesting that alterations in maternal care underlie the biological changes induced by maternal separation in the neonate.
Maternally separated rat pups also demonstrated alterations in the function of endogenous opioids. This is likely a byproduct of hypothalamic CRH hypersecretion that induces pituitary secretion of proopiomelanocortin, a prohormone that is not only a precursor of ACTH but also of melatonin and certain endogenous opioids. In studies that used a hotplate for nociception testing, maternally separated rat pups demonstrated increased response latencies believed to be a consequence of the analgesic effects of elevated concentrations of endogenous opioids
(135). This effect was potentiated by morphine and blocked by pretreatment with the opioid antagonist naloxone
(135). A recent study indicated that adult male rats subjected to repeated maternal separation during the first 2 weeks of life exhibit diminished morphine antinociception during testing with a hotplate
(136). Since the effect was not witnessed in adult female rats, this study raises interesting questions regarding the gender-specific effects of diminished maternal care on the maturation of opioid systems.
A limited subset of rodent studies has investigated the effects of maternal deprivation on other neurotransmitter systems. Although there is some inconsistency in the data, numerous studies have suggested that the activity of the principal inhibitory CNS neurotransmitters is diminished in adult rats previously exposed to maternal separation. For example, maternally deprived adult rats in novel situations exhibited heightened anxiety-like behaviors and showed evidence of reduced GABA activity, including decreased GABA
A binding, and decreased levels of CNS benzodiazepine type I receptors
(137). These findings contrast with earlier findings that the offspring of attentive female rats who more frequently groomed their young showed less fearfulness in novel situations and increased CNS levels of benzodiazepine type 1 receptors
(138). Surprisingly little attention has been devoted to the effect of rodent maternal deprivation on the ontogeny of central 5-HT systems. Maternal deprivation has been associated with decreased 5-HT concentrations in the dorsal hippocampus and prefrontal cortex of offspring
(139), but this finding is in contrast to other results demonstrating no alterations in the density of 5-HT fibers in the prelimbic, anterior cingulate, and precentral medial cortices of maternally deprived rats
(140). Although only tangentially relevant to animal modeling of parenting, postweaning social isolation of young rats has been shown to decrease the serotonergic innervation of the hippocampus
(141) but to have no impact on 5-HT concentrations in the hedonic circuitry innervating the nucleus accumbens
(142). To our knowledge, only one rodent maternal deprivation study has examined the function of neuropeptide Y, another transmitter with putative inhibitory activity at the locus ceruleus
(143). In that study, pups demonstrated increased expression of preproneuropeptide Y mRNA immediately after a 24-hour separation. Because the pups were also deprived of food and water during the 24-hour separation, it is difficult to ascertain whether the result was primarily an effect of nutritional deprivation or of disrupted maternal care.
Social and maternal deprivation in primates
Scrutiny of the primate social deprivation model has been quite extensive. Initial reports accentuated its value as a homologous model for depression
(144). Experimental refinements in which the monkey’s mother is replaced with a series of incrementally more realistic maternal surrogates
(145) have lent the model utility for investigating the consequences of disrupted parenting.
Most neurobiological research using the social deprivation model has focused on catecholamine systems. Monkeys reared with no attachment object or an inanimate surrogate exhibit depressive-like symptoms during later separations and markedly lower CSF norepinephrine concentrations
(146). In most studies, although not all
(147), peer-raised monkeys have exhibited less pronounced despair on separation and have shown baseline CSF norepinephrine concentrations that are intermediate between those of mother-reared monkeys and total isolates
(148–
150). These findings suggest an inverse relationship between maternal contact, norepinephrine neuronal activity, and later socialization. It is noteworthy that desipramine administration produced elevations in CSF norepinephrine concentrations largely consistent with its known pharmacology in both peer-raised and mother-raised monkeys
(148). Differential rearing may
(147) or may not
(148) produce alterations in CSF concentrations of 5-HT metabolites. One intriguing study did indicate that maternally deprived monkeys with the short rhesus macaque serotonin transporter allele (
rh5-HTTLPR) exhibit an exaggerated expression of the attentional and emotional alterations commonly associated with this genotype
(151).
HPA axis function has also been studied in socially deprived primates. A study comparing socially isolated to peer-reared rhesus monkeys demonstrated higher basal cortisol levels in the isolates but no differences between groups in cortisol responses to a novel situation
(152). Another study comparing peer-reared to mother-reared rhesus monkeys surprisingly reported higher basal concentrations of ACTH and more pronounced stress-induced increases in both ACTH and cortisol in the mother-reared offspring
(153) that was not appreciably altered by antidepressant administration
(154). These surprising results contrast with more recent findings of higher cortisol increases during subsequent social separation in peer-raised than in mother-raised monkeys that were correlated with higher self-administration of alcohol under typical living conditions
(155).
The magnitude of social deprivation in these models extends beyond the loss of parent-child interaction. In some models, animals are deprived of an entire range of social interaction when they are removed not only from their mothers but from the social group as a whole. Therefore, findings from such research bear limited face validity to the study of parental depression
(156). In designs that recognize this limitation, other studies have evaluated the acute sequelae of brief maternal separation. For example, one study examined effects of 24-hour separation by comparing four conditions: total isolation, remaining with the mother, being placed with a peer of the same group, or being placed with a peer from another group
(157). Thirty minutes after separation, the isolated infants and the infants placed with a strange peer had markedly elevated cortisol concentrations; 24 hours later only the isolated infants exhibited hypercortisolemia. Thus, the presence of peer-aged conspecifics modulated the acute biological stress response.
Other studies have modified the manner of separation from the infant’s mother. A study of 24-hour separation placed infants in one of three conditions: total isolation, remaining with the mother, or separated from the mother but able to see her
(158). In this study, the totally isolated infants were the least likely to vocalize but had the highest plasma cortisol concentrations; there were no detectable differences in the behavioral measures or cortisol concentrations of the other two groups. In a similar study in which infants were totally isolated, left with the mother, or placed in an adjacent cage with the mother in full view, elevations in plasma cortisol and CSF catecholamine metabolites (i.e., 3-methoxy-4-hydroxyphenylglycol, homovanillic acid) were greatest in the total isolates and at an intermediate level in the infants that were separated from but able to see the mother
(159). In yet another primate study, monkeys were subjected to a 2-hour separation from their social group at the ages of 6 months and 1 year to investigate the impact of aging on the HPA axis response to separation
(160). In this investigation, there were considerable individual differences in both the behavioral and cortisol responses to separation at both intervals, although the cortisol response was attenuated at 1 year of age. A recent study suggested that these alterations in HPA axis responsivity may be a consequence of changes in negative feedback mediated by the glucocorticoid receptor
(161).
Variable foraging in primates
Although separation paradigms that interfere with the quality of parenting reliably induce neurobiological and behavioral alterations in both rodents and primates, these strategies depend on an experimental intervention that is disparate from normal human experience. The face validity of the research protocol may be improved by instead altering a naturalistic stressor that indirectly stresses the infant by contesting the parent’s ability to provide adequate care.
All mammalian species face the challenge of feeding their offspring. The difficulty of this task depends both on the availability of a food source and the proficiency of the parent in procuring food from that source. During periods of nutritional stress, mammalian parents may struggle heroically to feed their young
(162), but they may also abandon their young
(163). Incorporating this reality of survival into an experimental paradigm provides a technique for modulating parental stress and thereby investigating the adequacy of parental care.
The objective in these studies is not to subject the animals to malnutrition but to increase the effort of parenting while permitting adequate nutrition. If the model achieves this goal, then the effects of parental stress on offspring development can be ascertained without the confounding effects of aberrant nutrition. A suitable primate model has been implemented by using three foraging conditions: low foraging demand, high foraging demand, and variable foraging demand. In the low foraging demand condition, food is readily available and requires no maternal effort for procurement. By contrast, in the high foraging demand condition, mothers are required to perform a task such as digging through wood chip bedding to obtain food. In the variable foraging demand condition, the requirements for food procurement are unpredictable. In this paradigm, the high foraging demand condition serves both as a comparator state and a nutritional control.
Infants raised in the variable foraging demand condition exhibited behavioral alterations suggestive of insecure attachment that resemble anxiety in humans
(164–
168). As juveniles and adults, primates raised under variable foraging demand conditions exhibited persistently elevated CSF CRH
(169,
170). The CRH elevations, however, were not accompanied as anticipated by elevations in cortisol. Instead, these offspring demonstrated lower CSF, and presumably plasma, cortisol concentrations than the comparator groups
(169). This finding is strikingly similar to the profile of HPA axis activity reported in patients with PTSD
(130–
132). A follow-up study demonstrated that CSF concentrations of somatostatin, a peptide whose release has been reported to be potentiated by CRH but that itself suppresses HPA axis activity, was also elevated in adult primates raised under variable foraging demand conditions
(171). Thus, hypersecretion of somatostatin is one potential explanation for the conundrum of decreased cortisol release in the face of CRH hypersecretion. In primates raised in variable foraging demand conditions, the growth hormone response to clonidine was inversely correlated with CSF CRH concentrations
(172), suggesting a potential mechanism for this often reported finding in depressed patients.
The impact of variable foraging demand on biogenic amine systems has had limited investigation. CSF concentrations of 5-HT and dopamine, but not norepinephrine, metabolites were elevated in adults reared in conditions of variable foraging demand
(171). The 5-HT results are consistent with previous findings of hyposensitivity to a pharmacological challenge with the 5-HT agonist
m-chlorophenylpiperazine in subjects reared in variable foraging demand conditions; however, the norepinephrine findings are surprising given the behavioral sensitivity to the α
2 antagonist yohimbine exhibited by variable foraging demand subjects
(173).
Prenatal Stress Models
It is not surprising that many view parenting as beginning not only before birth but even before conception. For example, avian species diligently build nests before laying their eggs. Likewise, agencies such as Planned Parenthood encourage prospective parents to prepare for parenting before conception. This is an important concept because there is growing evidence that the quality of prenatal parenting has a significant impact on the well-being of the offspring.
Mental illness during pregnancy may well represent the first adverse life event for the developing child
(174). For example, maternal depression during pregnancy may slow fetal growth
(175,
176), increase the risk of obstetrical and postnatal complications
(177–
180), and precipitate long-term behavioral changes in the offspring
(181,
182). These observations have tremendous implications for perinatal psychiatric practice.
A long line of animal research has investigated the effects of prenatal stress on the developing offspring. Consistent with the limited human data, data from animal studies have suggested that stress during pregnancy can adversely affect offspring growth
(183–
187), learning ability
(188–
191), and attainment of developmental milestones
(192). Prenatal stress may also induce persistent behavioral aberrations. In particular, adult rats exposed to prenatal stress continued to exhibit anxiety-like behaviors in novel situations
(193–
198), depression-like behaviors
(199), and exaggerated behavioral responses to stress
(200–
202). Primate studies have also demonstrated decreased exploratory behavior in prenatally stressed offspring
(203,
204).
As expected, these behavioral changes have been associated with long-term neurobiological changes. Pregnant rats exposed to an uncontrollable stressor exhibited increases in basal corticosterone concentrations and decreases in corticosteroid-binding globulin (further elevating the free fraction of corticosterone)
(205). Fetuses of these stressed pregnant dams also demonstrated increased concentrations of plasma corticosterone
(205). Initial evidence suggested that fetal hypersecretion of CRH may underlie the increased HPA axis activity. For example, repeated but brief exposure to prenatal stress increased the expression of CRH mRNA in the paraventricular nucleus of the fetal hypothalamus; however, prolonged stress exposure induced neuronal apoptosis
(206).
Prenatally induced alterations in HPA axis activity may persist into adulthood. For example, prenatally stressed adult rats demonstrated exaggerated corticosterone responses to subsequent mild stressors
(202,
207–212) and showed a phase advance in the circadian rhythm of corticosterone secretion
(213). Basal concentrations and stress-induced elevations of ACTH have been reported to be higher in prenatally stressed rats than in comparison animals
(208,
214). These changes are accompanied by a down-regulation of glucocorticoid receptors at the hippocampus
(209). The elevations in ACTH concentrations are paralleled by heightened concentrations of β-endorphin in prenatally stressed rats at 10 days of age
(215). The effects of prenatal stress on the ontogeny of opioid systems may have consequences both for the behavioral responses to novelty
(216) and the differentiation of sexually dimorphic behaviors
(217). However, measures of CRH secretion have been inconsistent. One study of prenatally stressed adult rats did not detect any alterations in median eminence CRH concentrations
(100), but another reported increased concentrations of CRH in the amygdala
(218).
The mechanism by which maternal stress during pregnancy modulates the fetal HPA axis is unclear. Placental passage of maternal corticosterone during prenatal stress exposure may be one mechanism by which stress exposure during pregnancy affects the offspring. For example, eradicating maternal corticosterone via bilateral adrenalectomy before stress exposure obviated the impact of prenatal stress on neonatal hippocampal glucocorticoid receptors
(219). However, the finding that neonatal adoption reversed the impact of prenatal stress on hippocampal glucocorticoid receptors suggests that the effect may be mediated through changes in postnatal maternal care
(209). Consistent with the latter hypothesis is evidence that stress during pregnancy adversely affected postnatal efforts at maternal care
(220,
221).
There has been relatively little study of the impact of prenatal stress on primate HPA axis activity. In one study, prenatally stressed juvenile rhesus monkeys exhibited increases in basal and stress-responsive ACTH and cortisol, compared with juveniles whose mothers were not stressed
(222). Another study in which the administration of ACTH to pregnant rhesus monkeys produced developmental changes similar to other stress paradigms during pregnancy lends additional credence to the theory that HPA axis hyperactivation mediates the long-term sequelae of prenatal stress
(223).
Preclinical studies have also indicated that prenatal stress affects biogenic amine systems. For example, prenatally stressed animals have exhibited alterations in the plasma concentrations of norepinephrine, dopamine, and their metabolites
(212,
224–229) that were consistent with increases in the turnover of these catecholamines. Furthermore, prenatal stress has been associated with alterations in both norepinephrine
(230) and dopamine receptors
(231). Changes in the asymmetric patterns of dopamine neural circuitry in the CNS of prenatally stressed adult rats have also been reported
(232,
233).
Prenatal stress has also been associated with long-term alterations in serotonergic activity. Decreased plasma and CNS concentrations of 5-HT and its metabolites in prenatally stressed animals have been reported
(212,
234,
235). 5-HT receptor profiles in the hippocampus
(234,
236) and behavioral responses to 5-HT challenge
(237,
238) were also changed in adult rats subjected to prenatal stress. These changes in serotonergic function may in part be programmed by increased glucocorticoid exposure during pregnancy
(239). Moreover, the effect of prenatal stress on cholinergic neurotransmission appears to be mediated by CRH activity at the hippocampus
(240).
Maternal stress during pregnancy is associated with persistent behavioral and cognitive changes in both rodents and nonhuman primates, paralleling concerns arising from clinical psychiatric experience. The long-term neurobiological effects may be mediated in large part through organizational effects of maternal HPA axis activation on fetal CNS maturation
(241). Although it remains unclear whether the placental passage of maternal glucocorticoids or the impact of inadequate postnatal care mediates the alterations in offspring HPA axis function, the ultimate clinical importance of these data is the same. It would seem prudent to treat perinatal maternal mental illness aggressively to circumvent the potential adverse impact of maternal illness on the developing child.
Two-Parent Models
Despite the considerable breadth of animal research investigating parental care, the study of paternal care is woefully inadequate. Throughout most modern cultures, humans are monogamous (albeit in many cases serially so) and provide biparental care (although mothers continue to provide a disproportionate share of child care). However, single-parent households are increasingly prevalent, and we have made little progress in understanding the behavioral and neurobiological consequences of single parenting. Without this knowledge, our efforts to educate and guide single parents are not evidence-based and may therefore be seriously deficient.
Two-parent animal models that incorporate paternal care and consider the impact of its absence are sorely needed. Unfortunately, nonhuman primate research has provided little insight. Group-living primates typically are not monogamous; therefore, male primates have little assurance of paternity and from an evolutionary standpoint little incentive to provide parental care. In fact, adult male primates of several species interact no more often with their own offspring than with other infants
(242–
244). Adult male primates appear to initiate interaction with infants, not in an effort to provide care for their own offspring or even to curry favor with potential mates, but instead as part of a triadic interaction to buffer tensions with other adult males
(243). Numerous avian species mate for life and provide biparental care, but the complex neurobiological and behavioral issues requiring study are not likely to be addressed adequately in a nonmammalian species.
Fortunately, a small number of monogamous mammalian species that provide biparental care are being studied. In particular, rodent species such as the prairie and meadow voles, Siberian and Djungarian dwarf hamsters, and California mice have been studied. Most of this research has scrutinized the neurobiology underlying the affiliative behaviors of these species. There is clear evidence that oxytocin stimulates maternal behavior, mating activity in males, and infant attachment behaviors in these highly social creatures
(245,
246). A limited subset of these investigations have studied the determinants of paternal care and the impact on the offspring when paternal care is unavailable.
Research with voles has demonstrated that the father and the mother demonstrate distinct patterns of interaction with their young that suggest unique parental roles
(247). Furthermore, the presence of the father during infancy has a positive effect on the subsequent paternal behavior of male offspring. Juvenile male voles allowed to remain in the nest when a subsequent litter is delivered were more likely to provide alloparental care if their fathers were present during upbringing
(248). Similarly, paternal care was more readily initiated by adult male voles reared as neonates with their fathers present rather than with unfamiliar males
(249). However, exposure to pheromones of the mate also facilitated the initiation of paternal care
(250). In addition, continued exposure to pups eventually induced paternal care even in voles raised without a father
(249,
250).
The success of another species, the Djungarian dwarf hamster, appears to be particularly dependent on the participation of both parents in rearing the offspring. Pup survival was compromised by the removal of the father
(251). The introduction of a second unmated female to the nest did not compensate for the father’s absence
(252).
The biological substrate for paternal behavior in these species remains unclear. It is unlikely to be solely predicated on oxytocin. In males of numerous species including humans, oxytocin has been shown to potentiate sexual behavior and to facilitate the ejaculatory response
(245). A study with the monogamous, biparental California mouse, which provides biparental care, has shown that plasma oxytocin remains elevated in the male after copulation and during early gestation but declines before delivery and remains low throughout lactation
(253). Preliminary evidence indicated instead that vasopressin may potentiate the effects of androgens to induce paternal behavior in these rodent species
(254,
255).
Although the data produced to date are limited, two-parent, monogamous animal models offer numerous advantages. They provide greater face validity for human parenting by affording an opportunity to investigate paternal care. They also furnish a means to study the impact of parenting without a father. Future studies should investigate the neurobiological effects of paternal separation not only on the developing offspring but also on the mother who remains behind to care for her litter. Unfortunately, no two-parent primate models have been identified.
Conclusions
An impressive array of evidence indicates that stressful events in early life contribute significantly to the vulnerability for adulthood psychopathology (
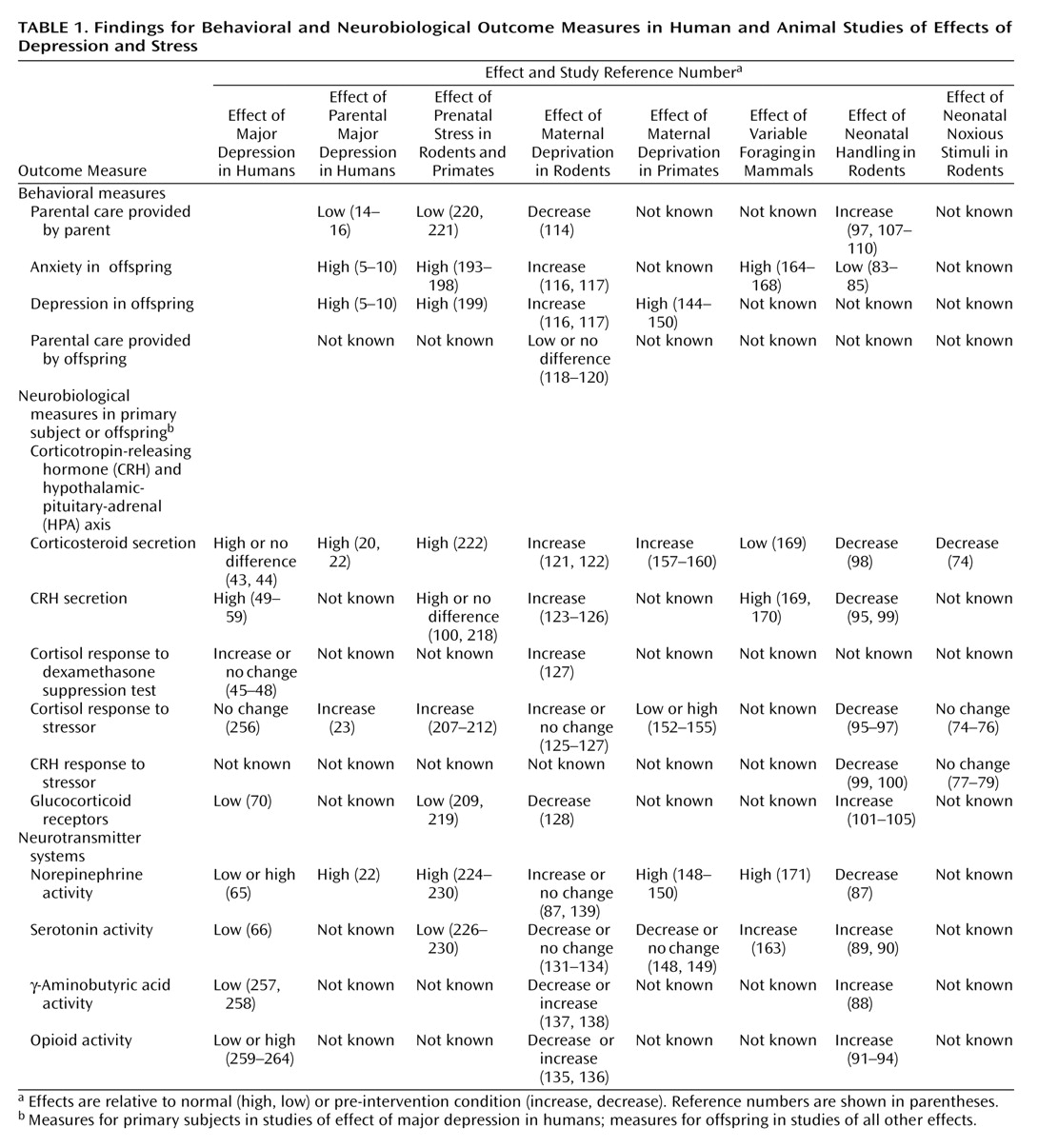
Table 1). Parental nurture helps to foster adaptive responses to early life stressors, thereby protecting the child from this vulnerability to later illness; however, parental depression may undermine this protection and occasionally is itself the source of early life stress. Increasing evidence substantiates the observation that parental depression contributes to the child’s diathesis for subsequent stress-related illness.
Although ideal parenting may be poorly defined, the animal literature indicates that deficits in parental care may lead to persistent adverse sequelae for the offspring. As animal modeling of human parental behavior is further refined, we will gain additional insights into the qualitative impact on psychiatric vulnerability in the offspring of events such as depression that interfere with parental nurture. Animal models already are an important component of psychiatric research offering several distinct advantages over human research. First, some species such as rodents have brief gestational and developmental periods, enabling researchers to study outcomes in an expeditious manner. Second, animal models permit researchers to invoke particular environmental manipulations (e.g., maternal deprivation, food supply limitation, pharmaceutical challenge) that would be impractical in naturalistic human studies. Third, invasive measurement of CNS activity is only feasible in an animal model. Despite these advantages, an animal model with poor homology to the human experience is not particularly useful. Defining and validating homologous animal models is in fact often the most challenging aspect of preclinical psychiatric research.
Homologous studies relevant to our consideration of parental depression have used both rodent and primate models. Among the existing animal paradigms, the variable foraging model demonstrates the greatest face validity for parental depression in humans. This model avoids direct interaction between the researchers and the animal offspring. Furthermore, the variable foraging model does not abolish parental care (even transiently) but introduces instead an environmental stressor that challenges the maternal capacity for caregiving. Although this more closely parallels some of the difficulties encountered by depressed parents, it must be acknowledged that the stress of a variable environmental perturbation is not fully homologous (i.e., does not bear absolute construct validity) to the presence of a stress-related illness (i.e., depression).
Both prenatal and postnatal experimental laboratory stressors that either directly or indirectly impair maternal care produce a persistent deleterious impact on offspring. By contrast, the only early life stressor studied to date that tends to bolster maternal care, i.e., neonatal rat handling, typically produces adaptive behavioral effects and reduces neurobiological sensitivity to subsequent stress in the offspring. Consequently, the neonatal stress studies exhibit predictive validity with regard to the clinical sequelae of parental depression in that they demonstrate that deficits in maternal care are associated with anxiety-like and depression-like behaviors in the offspring that commonly persist into adulthood. The predictive validity is further underscored by the fact that the stress paradigms also uniformly produce acute hyperactivity and long-term hyperresponsivity of HPA axis function. The key event in these HPA axis changes appears to be CRH hypersecretion. This pattern bears striking parallels to findings in human studies of acute stress responses and may also be germane to the pathophysiology of stress-related illnesses, including depression. In addition, CRH not only drives the endocrine stress response but also integrates immunological, autonomic, and behavioral responses. Manageable designs for multisystem stress studies are only now being contemplated.
Once appropriate homologous animal models of parental depression are identified, they should dramatically hasten the progress of this important line of research, which is currently hampered by the inherent limitations of conducting neurobiological research with children at risk. We anticipate that this research will ultimately help clarify the alterations in central CRH function that are believed to exist in those vulnerable to psychiatric illness by virtue of early adverse life events such as parental depression. Moreover, appropriate animal models will ease the implementation of multisystem research. For example, because most norepinephrine-secreting neurons emanate from a single brain stem region, the locus ceruleus, interaction between the norepinephrine system and other neurobiological systems is robust. Future research will undoubtedly investigate the homeostatic interactions between norepinephrine activity and other biological systems. The well-documented stimulatory effects of CRH
(265,
266) and glutamate
(267,
268), in addition to the inhibitor effects of 5-HT
(269–
271), GABA
(272), endogenous opioids
(265,
273), and neuropeptide Y
(274), on locus ceruleus activity provide one potential framework for future cross-system research.
The preclinical findings regarding parenting carry profound clinical implications. First, it is imperative that we better understand the mechanisms whereby parental depression contributes to the developing child’s subsequent psychiatric vulnerability. These animal data may direct the development of pharmacological interventions for a child once an insult from parental depression has occurred. Such treatments may find utility both for children in the acute aftermath of a childhood stressor and for adult offspring with a persistent vulnerability to illness that is, at least in part, a consequence of the persistent effects of depressive illness suffered by their parents.
Second, these animal data pose a seminal clinical question relevant to the psychiatric care of parents with young children. Does parental mental illness pose the first adverse life event for a child
(174)? The preclinical data have, for example, been absent in many of the proposed treatment guidelines for perinatal psychiatric illness
(275–
279). We contend that the clinician should consider the preclinical data regarding the risk to offspring posed by untreated parental mental illness during the child’s formative years when planning treatment for both men and women with children
(280,
281). Data from animal parenting research, for reasons we have described, have helped to delineate the potential adverse consequences of parental depression and must be incorporated into rational clinical decision making in the context of parental mental illness.