“Shyness,” as used in developmental psychology, refers to a pattern of behaviors and emotional states that includes inhibition of approach behaviors and discomfort on exposure to unfamiliar people and situations, a constricted social life, and physiological markers such as high heart rate and cortisol levels
(1). The term may encompass a broad range of severity, from normal traits such as mild social awkwardness to clinical disorders such as totally inhibiting social phobia. Shyness is often pervasive, in that it is manifest at school, at home (with strangers), and in other social situations. Various authors have suggested that its temperamental antecedents can be seen as early as the first 6 months of life
(2) and that it is an important risk factor for clinical anxiety disorders in childhood and later
(3).
Numerous studies have examined genetic factors involved in shyness, and socially related fears and heritability have been shown to be significant
(4). However, to our knowledge, association of a specific gene to shyness has been shown in only one study of a phenotype—behavioral inhibition
(5). In that study, four candidate genes first flagged in mouse models of emotionality were examined in a selected group of behaviorally inhibited children, and a modest association with glutamic acid decarboxylase was observed.
We assessed shyness after recruitment of unscreened second-grade children using a composite questionnaire administered to the children, parents, and teachers. Children were initially genotyped for the serotonin transporter promoter region 44 base pair deletion/insertion polymorphism (5-HTTLPR), which has been demonstrated to moderate transcriptional efficiency and was originally associated (reviewed in reference
6) with anxiety-related personality traits
(7). Other studies have shown an association between the 5-HTTLPR polymorphism and obsessive-compulsive disorder (OCD)
(8–
13) and autism
(14–
16).
Results
The children were rated on shyness by themselves, their parents, and their teachers. Significant correlations were observed between parents’ and teachers’ ratings and by self-ratings of shyness (child and teacher: r=0.38, N=116, p<0.001; child and parent: r=0.72, N=118, p<0.001; parent and teacher: r=0.35, N=116, p<0.001). Cronbach’s alpha reliability, which measures internal consistency of test items, was also high for all three rating scales as well as for the combined scale (eight parent items: alpha=0.89, N=118; eight teacher items: alpha=0.88, N=116; six child self-rating items: alpha=0.70, N=118; 22 total items: alpha=0.90, N=116). For the genetic analysis, we used the combined rating scale, arrived at by summing the parent, teacher, and child questionnaire ratings.
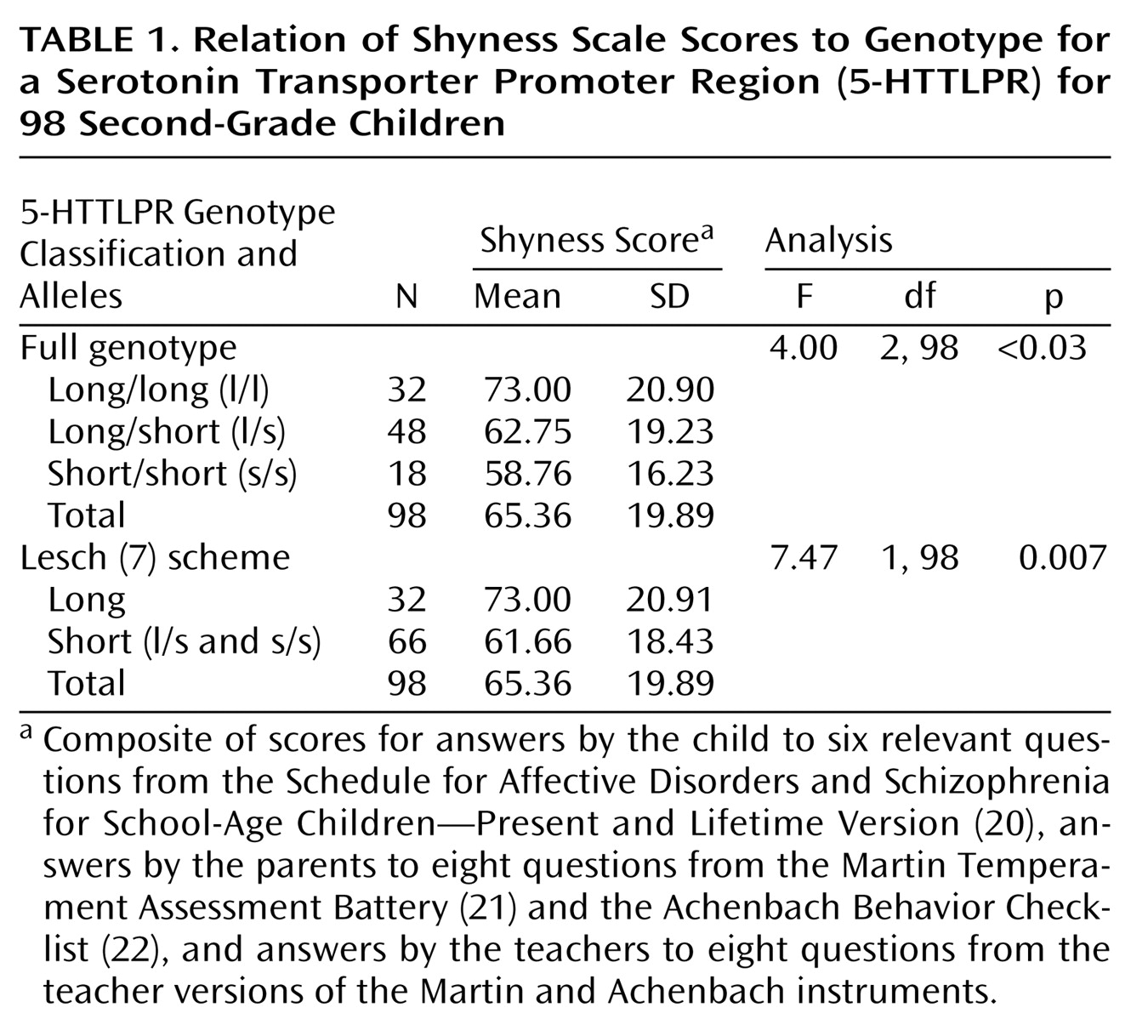
Shyness scores were grouped by all three (long/long homozygotes, short/long heterozygotes, and short/short homozygotes) genotypes and also by a genotype classification (long/long versus short/short and short/long) first suggested by Lesch et al.
(7), which reflects the effect of the polymorphism on transcription (
Table 1). A significant association was observed between the long 5-HTTLPR polymorphism and shyness, both by the functional classification of Lesch et al. (ANOVA: F=7.47, df=1, 98, p=0.007, N=98; eta squared [a measure of effect size]=0.072, observed power=0.77) as well as by consideration of all three genotypes (ANOVA: F=4.00, df=2, 98, p<0.03, N=98; eta squared=0.078; observed power=0.70). Note that after Bonferroni correction a significant difference at the p<0.05 level was observed between the long/long versus the short/short genotypes. The Bonferroni correction provided by ANOVA performed by SPSS is applicable only with more than two independent variables (in this instance, the long/long, long/short, and short/short genotypes). Of interest, the boys were the main contributors to the association (Lesch scheme [ANOVA], only boys: F=12.53, df=1, 64, p=0.001, N=65; only girls: F=0.04, df=1, 32, p=0.85, N=33). Genotype frequencies were distributed according to the Hardy-Weinberg equilibrium.
Another way of estimating the effect size of the 5-HTTLPR genotype on shyness scores is from the correlation coefficient derived from a linear regression analysis of our data. For example, in our data set, Pearson’s correlation coefficient was 0.269 (p=0.004, one-tailed) for this analysis, which was obtained by regressing composite shyness scores on the Lesch genotype classification scheme. Such a correlation is similar to an odds ratio of ∼1.2–1.9, which, by Cohen’s definition
(27), is a small effect.
Since population admixture and stratification may confound interpretation of association studies, we examined the distribution of the 5-HTTLPR genotype in a large sample that represents the two principal ethnic groups examined in the present study. These subjects were previously recruited in our studies of normal personality
(6). No significant difference in genotype frequency (long/long, long/short, or short/short) for the 5-HTTLPR genotype was observed between the Ashkenazi (N=575) and non-Ashkenazi (N=383) Jewish populations (χ
2=0.01, df=2, p=1.00). Genotype frequencies were in Hardy-Weinberg equilibrium.
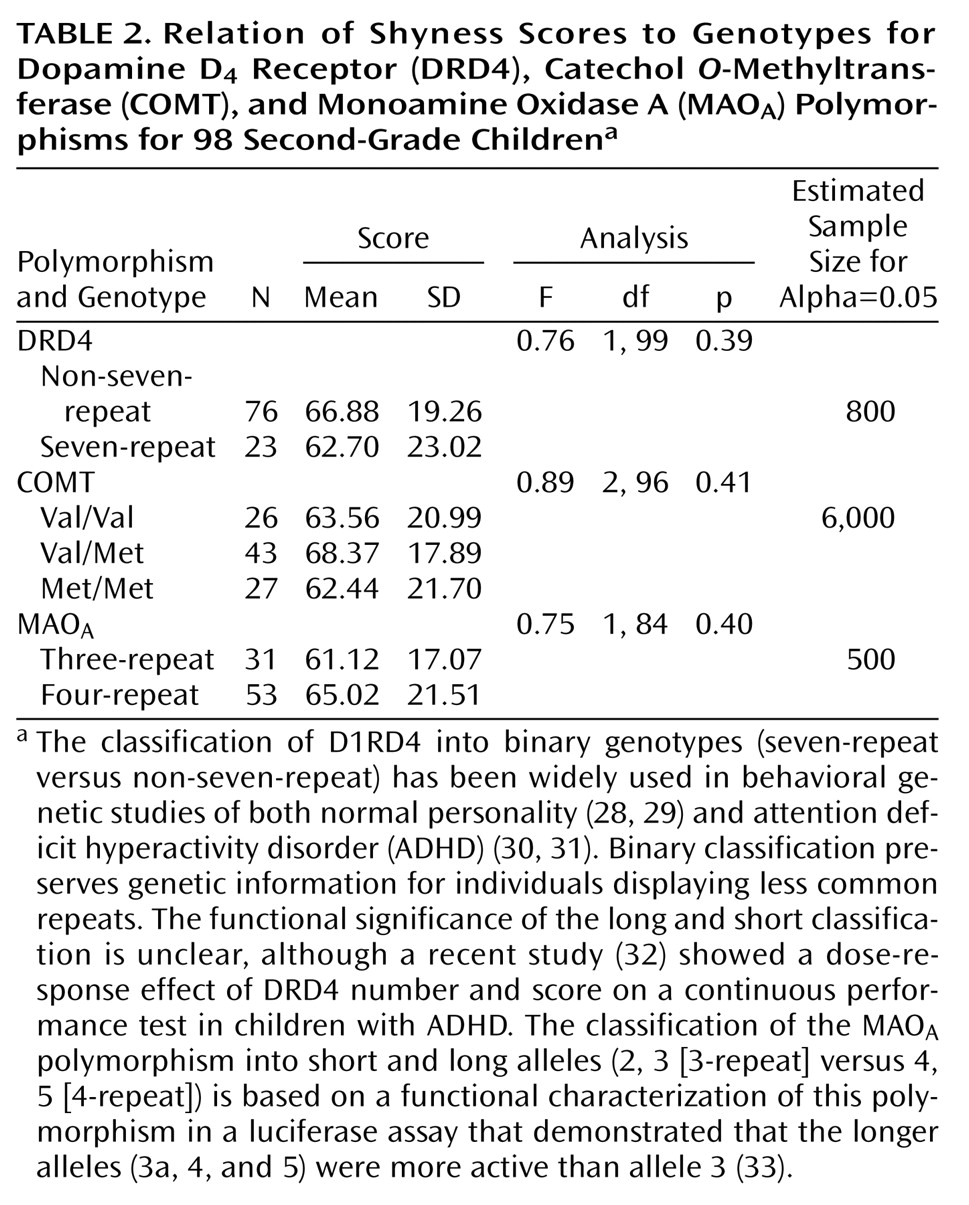
We also examined three additional polymorphisms for possible association with composite shyness scores: the DRD4, COMT, and MAO
A (
Table 2). No association was observed for any of these genes with shyness (DRD4: F=0.76, df=1, 99, p=0.39, observed power=0.14; COMT: F=0.89, df=2, 96, p=0.41, observed power=0.20; MAO
A: F=0.75, df=1, 84, p=0.40, observed power=0.14). We included in
Table 2 the estimated sample size for a significant effect for each of these genes at alpha=0.05, based on observed differences in mean values and standard deviations in the current sample.
A Bonferroni correction for multiple testing (four genes tested) showed that the association with the 5-HTTLPR polymorphism was still significant (4×0.007=0.028).
Discussion
The current study shows an association between shyness measured in a group of second-grade schoolchildren by using a composite rating scale (of self-reports, parent reports, and teacher reports) and the long allele of the 5-HTTLPR polymorphism. Shyness, a phenotype akin to behavioral inhibition
(34)—a well-described childhood temperament—has been suggested to be an antecedent of later anxiety disorders in adolescents and adults, especially social phobia
(35). Although there is considerable evidence that genetic factors significantly contribute to the etiology of social phobia
(4), the current study is one of a very few investigations
(5,
36) that has examined the role of a specific and common gene variant in conferring risk for this temperament construct. Stein et al.
(36) examined 17 multiplex families comprising 76 probands (from ages 18 to 65) who were diagnosed with social phobia and failed to observe linkage to 5-HTTLPR. However, as the authors noted, their study had power to observe only a major gene effect. In contrast to these two previous studies, we examined a group of unscreened schoolchildren in the second grade. Although using subjects who exhibit extremes of a phenotype in genetic studies is generally considered the most efficient strategy, a recent genome-wide linkage study on human stature (height)
(37) suggested otherwise. Indeed, the authors reached the conclusion that selective ascertainment of families may provide only modest gains in efficiency.
A longitudinal study from Australia that measured a broad range of temperament and behavioral problem variables found an association between the long 5-HTTLPR allele and higher anxiety at 13–14 and 15–16 years
(38), which is consistent with the current findings of an association between the same allele and shyness. At earlier developmental stages, no association was observed between temperament traits and the transporter gene. In previous studies
(39–
42), we examined the role of the 5-HTTLPR polymorphism in human temperament from 2 weeks to 1 year. However, in contrast to the present findings and the Australian longitudinal study
(38), it was the short allele (by an interaction with the DRD4 repeat variant) that enhanced avoidant-like behaviors during this early developmental period. It should be considered, however, that despite the apparent face validity of neonatal and infant temperament traits (distress to limitations, negative emotionality) to measures of adult behavioral traits (social and general anxiety), infant temperament is likely to be only weakly correlated with adult traits. Extrapolating gene effects on behavior from infancy to adult personality constructs, therefore, may lead to ambiguous conclusions. It is also worth noting that the original Lesch et al. finding
(7) showed an association between the short 5-HTTLPR alleles that was shown to reduce gene transcription (and presumably make more serotonin available in the synapse) and that adult neuroticism is opposite from what might be expected from the action of selective serotonin reuptake inhibitors (SSRIs).
The currently reported association of the long 5-HTTLPR allele with shyness may be relevant to recent findings in adult anxiety disorder, OCD, and this gene
(8–
13). Both a family-based
(12) and an association
(11) study showed associations between the long 5-HTTLPR polymorphism and OCD. OCD is often comorbid
(43) with social phobia and other anxiety disorders, suggesting the possibility that such disorders share some, but not all, common genetic determinants—for example, the 5-HTTLPR polymorphism. Such a notion is strengthened by the effectiveness of SSRIs across the range of anxiety disorders
(44). Of interest, clinical response to fluvoxamine in OCD patients is related to the 5-HTTLPR genotype
(45,
46), and carriers of the long allele are more responsive to treatment.
Some studies have also shown an association between the long 5-HTTLPR allele and autism
(14–
16). It is worth noting that Cook et al.
(47) found an association between autism and the
short HTTLPR allele. Tordjman et al.
(16) suggested that this promoter region polymorphism by itself does not convey risk for autism but instead influences the behavioral phenotypic expression of autism. A core phenotype in autism is the repetitive behaviors that also characterize OCD. Most intriguing is the epidemiological study of Smalley et al.
(48), showing that the frequency of social phobia—20.2%—in autistic families is approximately 10 times more common than that found among the relatives of comparison probands (2.4%).
Although we observed no differences between 5-HTTLPR genotype frequencies when grouped by the two main ethnic groups examined in this study, population stratification cannot be excluded as a contributing cause to the observed association. Future studies should include the use of more robust family-based designs, such as the transmission disequilibrium test
(49,
50). Additionally, more sophisticated case-control designs
(51) and the use of so-called genomic control subjects
(52) offer alternative strategies for handling population admixture in association studies.
We also examined three additional polymorphisms (DRD4, COMT, and MAOA) that have been studied in many other human behavioral genetic investigations and were therefore likely candidates for contributing to shyness. No association was observed between any of these polymorphisms and shyness in the current study. However, our failure to detect an association between these genes and shyness may reflect the difficulty in observing genes of small effect size in a sample of 98 children.
A word of caution is called for regarding association studies in general and the present report in particular. Ioannidis et al.
(53) noted that “validation of statistical hypotheses in genetic epidemiology is a task of unprecedented scale” because of the increasing number of candidate genes generated by the mapping of the human genome that are being intensively examined in association studies of human disease. Meta-analysis offers a robust solution to making sense of numerous investigations.
Similarly, the provisional association reported here between shyness and the long 5-HTTLPR allele needs to be examined using both case-control and family-based designs in numerous independent study groups before the validity of this observation is either strengthened or weakened. Nevertheless, the role of shyness as a risk factor for subsequent psychopathology and the importance of elucidating its genetic architecture suggest the importance of the current study as a catalyst that will undoubtedly generate additional investigations of this and other genes in contributing to this intriguing phenotype.
Shyness in the children we examined may be considered an intermediate phenotype (a so-called endophenotype
[54]) that is more proximally related to the genetic substrate than is the higher-order construct of a disorder (e.g., social phobia)
(5). To the extent that shyness is an intermediate phenotype, genetic influence on it may be stronger and less complex, making shyness in children an excellent target for genetic dissection that may shed light on adult anxiety disorders.