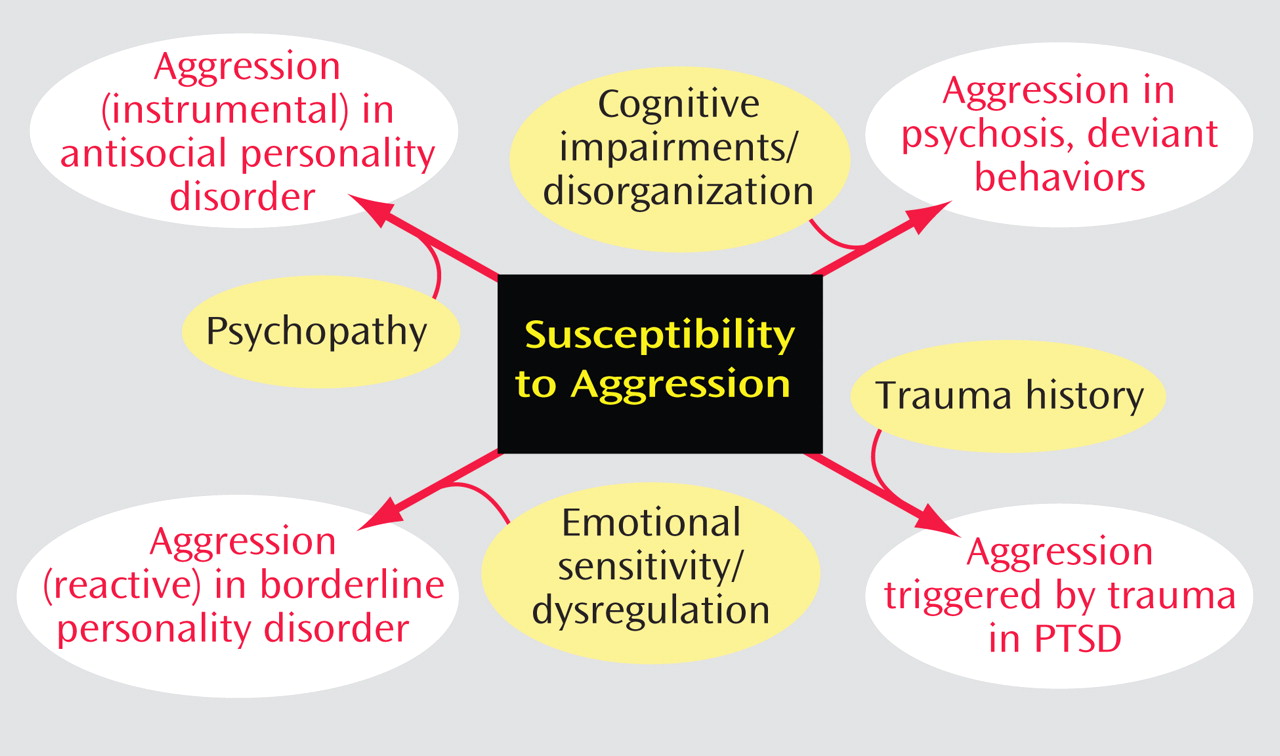
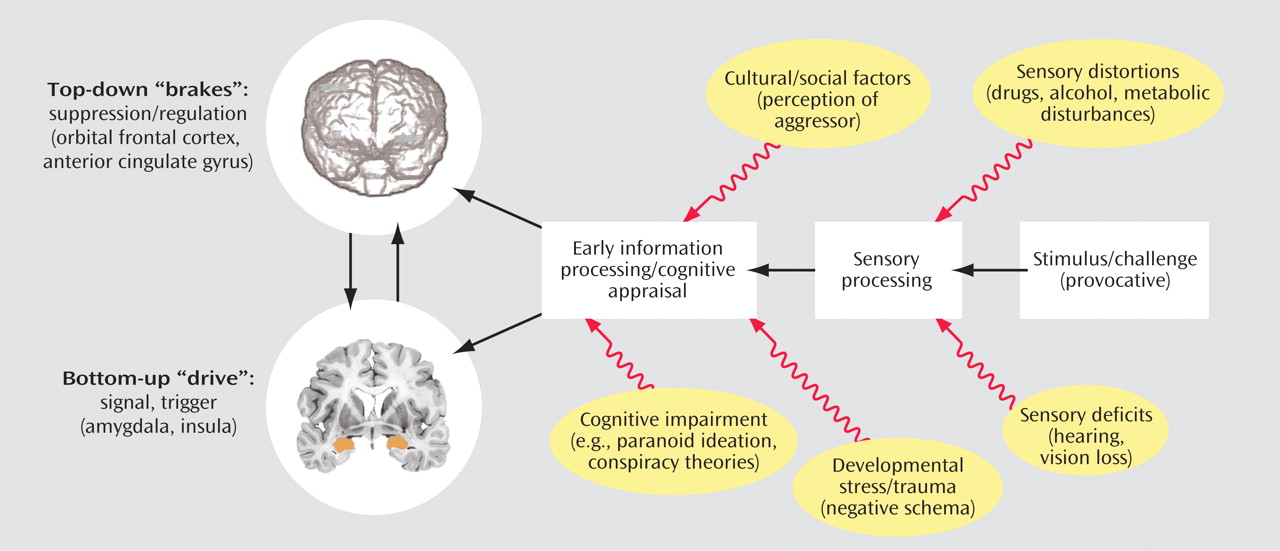
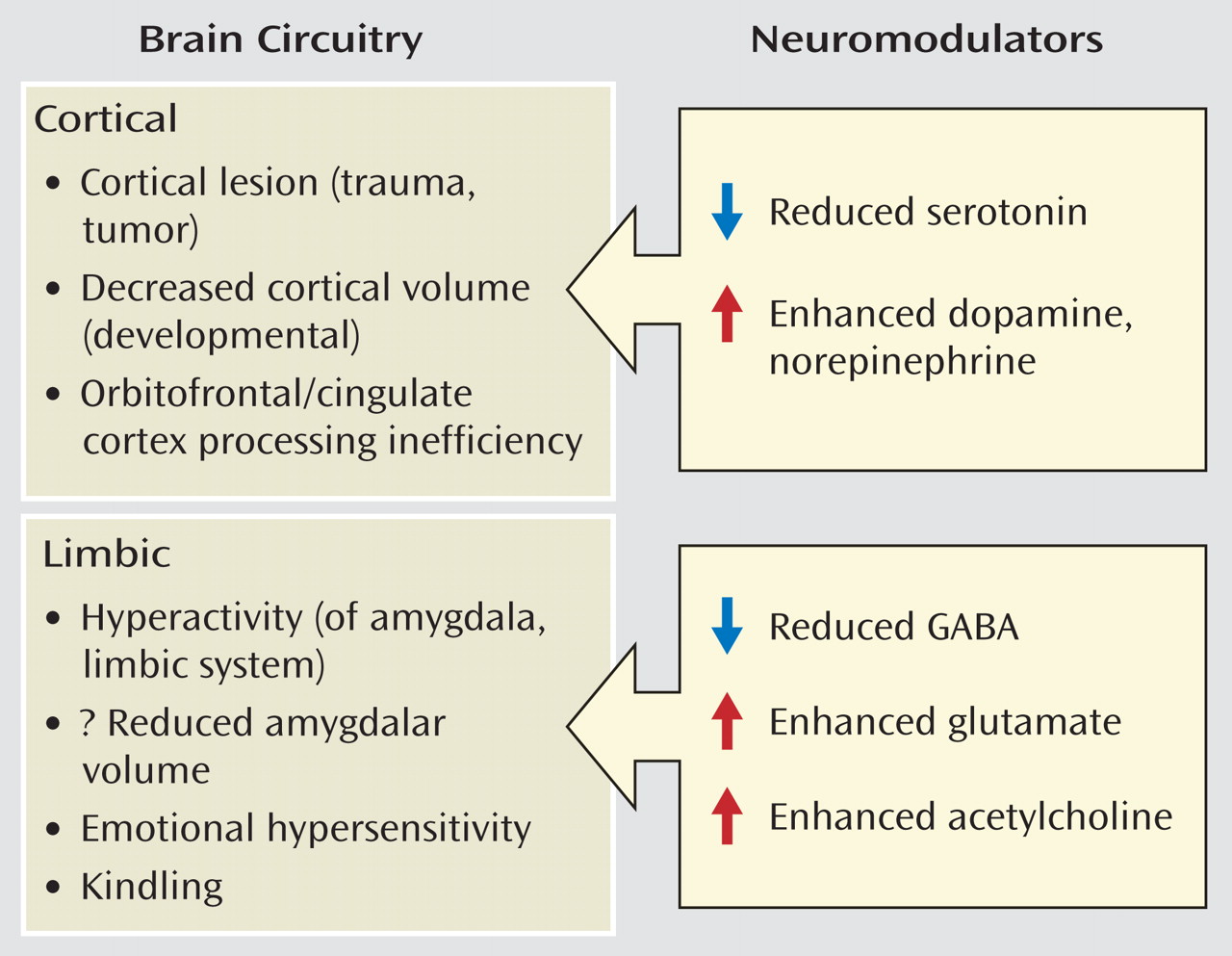
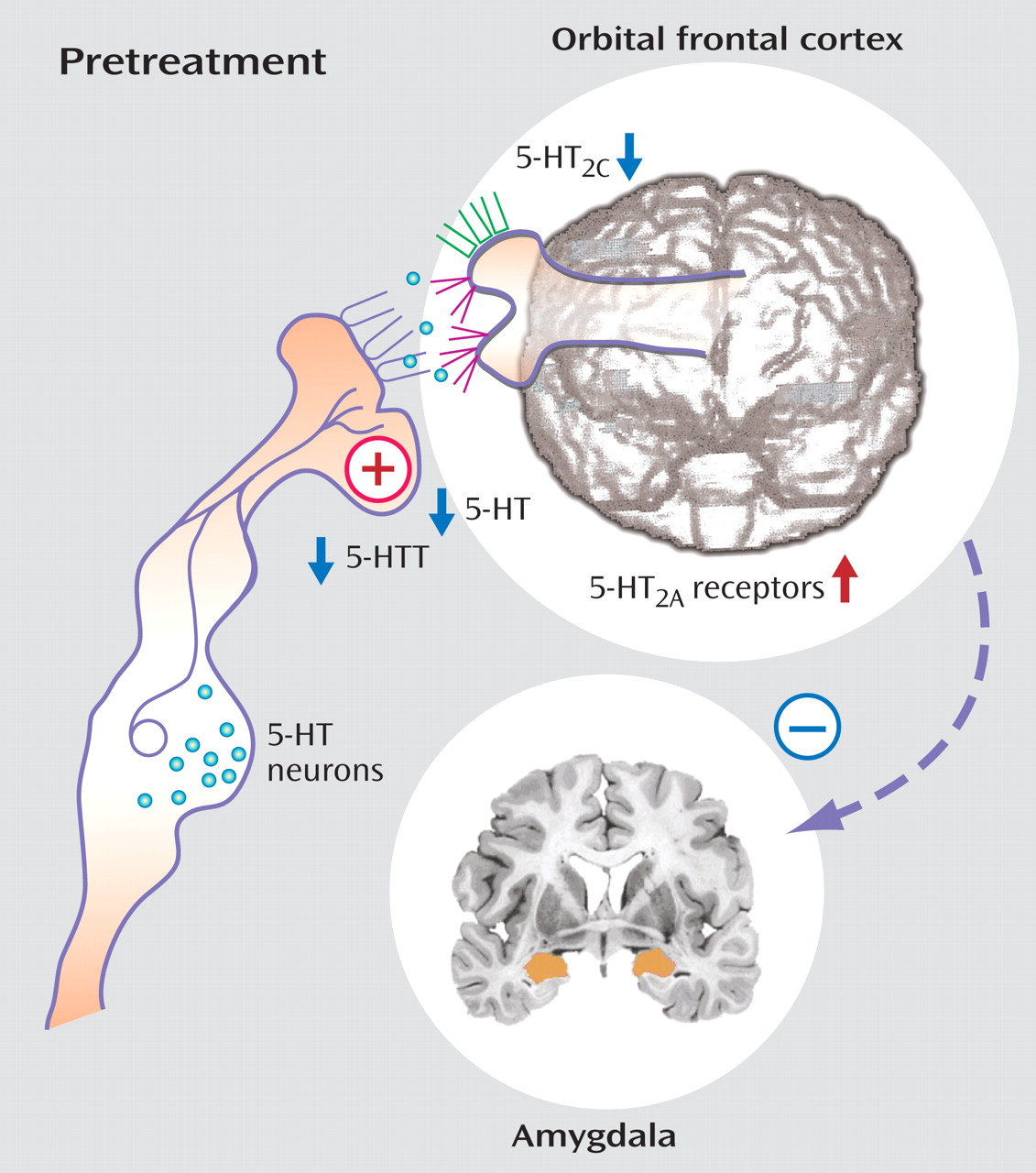
Serotonin
Serotonin facilitates prefrontal cortical regions, such as the orbital frontal cortex and anterior cingulate cortex, that are involved in modulating and often suppressing the emergence of aggressive behaviors primarily by acting on serotonin 5-HT
2 receptors in these regions. Thus, deficiencies in serotonergic innervation of these regions could be expected to result in disinhibited aggression upon provocation. This model is supported by studies that have shown that selective serotonin reuptake inhibitors (SSRIs) reduce impulsive aggression
(57) as well as neurobiological studies that have implicated reduced concentrations of the serotonergic metabolite 5-hydroxyindoleacetic acid (5-HIAA) and reduced neuroendocrine responses to serotonergic probes
(58 –
60) in aggressive personality disorder patients or individuals who have had violent suicide attempts
(61,
62) . Animal models in rhesus and macaque monkeys support an association between lower serotonergic activity and aggression in freely moving primates
(63) . Interestingly, serotonin depletion is also implicated in decreased learning of cooperation and diminished perception of trustworthiness
(64) .
Antagonists of 5-HT
2A receptors reduce impulsivity in animal models
(65), and atypical neuroleptics with prominent 5-HT
2A antagonism have antiaggressive efficacy in clinical populations
(66) . Agonists, on the other hand, reduce impulsivity at the 5-HT
2C receptor
(65), suggesting that the two receptor subtypes may have complementary roles in the regulation of aggression consistent with their reciprocal modulation in animal model systems
(67) .
Pharmacological probe studies assess net serotonergic activity by measuring responses of hormones, such as prolactin, to serotonergic releasing agents, including d, l-fenfluramine. Blunted prolactin responses are associated with suicide attempts in depressed and personality disorder patients and impulsive aggression in personality disorder patients, more specifically, borderline personality disorder and those criteria of borderline personality disorder reflecting impulsive aggression
(68) . Metachlorophenylpiperazine, which directly activates the 5-HT
2C receptor, has been associated with blunted prolactin responses and increased impulsivity and hyperactivity
(65 –
67,
69) . Ipsapirone and buspirone, which are agonists for the 5-HT
1A receptor, also produce reduced neuroendocrine responses
(11) . In paradigms assessing receptors on peripheral blood elements, increased 5-HT
2A receptor binding in platelets has been associated with experimentally induced depletions in serotonergic activity (e.g., in tryptophan depletion paradigms). Such increases are also correlated with impulsive aggression in subjects with personality disorders, suggesting increased aggression in the face of reduced serotonergic activity
(70) .
More recently, imaging studies have pointed to reduced orbital and ventral medial prefrontal activation in response to d, l-fenfluramine in impulsive aggressive personality disorder patients
(61), in borderline personality disorder patients (who are often characterized by impulsive aggression)
(71), and in depressed patients with a history of suicide attempts
(72) . Personality disorder patients with intermittent explosive disorder, as determined by integrated research criteria, characterized by periodic episodes of dyscontrol of aggression and violence, demonstrated diminished activity in the orbital frontal cortex and adjacent ventral medial cortex relative to comparison subjects but did not necessarily differ in other regions of interest, including the parietal cortex
(61) . Statistical parameter mapping suggested that this difference was highly localized to a small region of the orbital frontal cortex near the site of the lesion of Phineas Gage. A study of borderline personality disorder patients showed reduced activation to fenfluramine in similar regions
(71) . The glucose metabolic response to the selective 5-HT
2C agonist metachlorophenylpiperazine was also reduced in patients with intermittent explosive disorder, as determined by integrated research criteria, particularly in the left hemisphere and anterior cingulate, as opposed to the posterior cingulate
(37) . Comparison subjects demonstrated an expected positive correlation between the amygdala and prefrontal cortex activity at rest, while this correlation was zero order to negative in borderline personality disorder patients with intermittent explosive disorder, as determined by integrated research criteria
(37) .
PET imaging studies have illuminated the activity-specific components of the serotonergic system, including the serotonin transporter and 5-HT
2A receptor. Serotonin transporter activity as labeled by [
11 C](McNeil 5652) was reduced in number in the cingulate cortex in patients with aggressive personality disorder relative to comparison subjects
(73), and interim results using a more cortically sensitive probe, [
11 C]DASB, suggest significant reductions in transporter binding in both the anterior cingulate cortex and orbital frontal cortex in aggressive patients (unpublished data of Siever et al., 2007). Receptor binding of 5-HT
2A, which can be measured by [
11 C]MDL100907 or altanserin, is significantly increased in physically aggressive patients with personality disorders
(74) and in female borderline personality disorder patients
(75), consistent with postmortem
(76) and platelet studies
(77) suggesting increased 5-HT
2A receptor binding associated with suicide. These studies are consistent with 5HT
2A antagonism reducing impulsivity
(65), but such antagonists have yet to be studied in relation to impulsive aggression in clinical populations. Pilot data suggest reductions in 5-HT
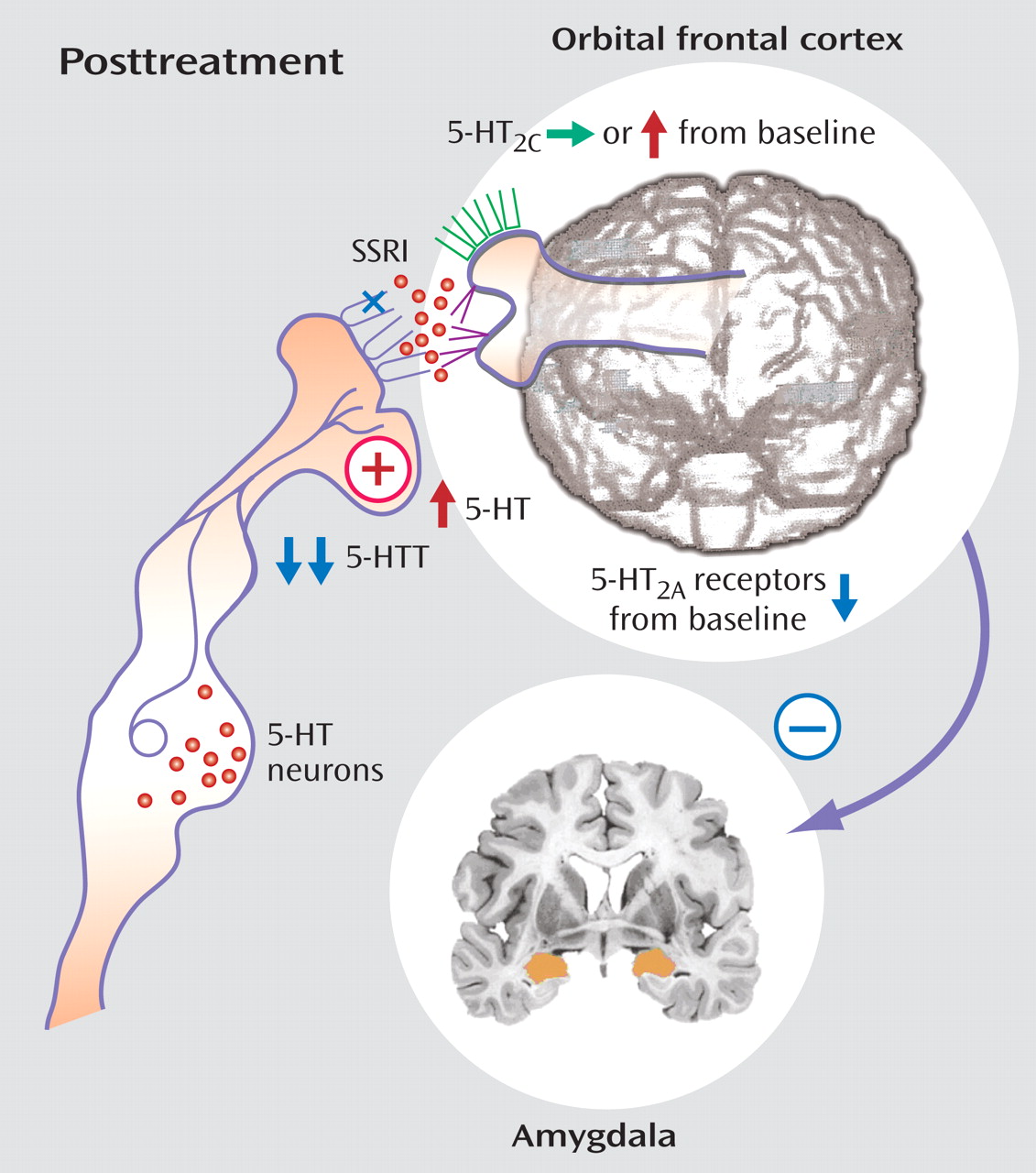
2A binding with treatment with the SSRI fluoxetine, which may parallel reductions in aggression symptoms (unpublished data of Siever et al., 2007), implicating increased 5HT
2A receptor sensitivity associated with aggression and its reduction with efficacious treatment.