Psychotherapy’s efficacy is well established for a wide range of emotional disorders. Despite this, no psychotherapeutic intervention works equally well for all patients, and the mechanisms through which psychotherapy reduces symptoms and enhances functioning remain difficult to specify. With the advent of neuroimaging technologies, researchers have new tools with which to identify clinically meaningful markers of brain function that are associated with treatment response. Two kinds of associations have been examined. Treatment outcome prediction studies seek to identify those patterns of brain function that confer a higher likelihood that a treatment will work. Treatment mechanism studies examine changes in the brain as a result of the intervention in question, in order to help understand how treatments are exerting their effects. Both types of study hold promise for developing a more complete understanding of the neural mechanisms involved in successful therapy, and both may guide future treatment refinement, novel mechanistic treatment development, and personalized treatment prescriptions tailored to individual patients.
Below, a brief overview of the general neural architecture believed to be relevant for anxiety, depression, and psychotherapeutic interventions for these disorders is provided. Anxiety and depression are the focus of this review because these are the two disorders for which the most evidence has accrued, and also because there is good reason to believe that anxiety and depression share at least some underlying neural mechanisms. Functional neuroimaging studies of psychotherapy are reviewed, and recent advances toward improving the methodology and clinical relevance of research in this area are highlighted. Ideally, work will continue to progress toward greater relevance and import for the practicing clinician.
Neural Circuitry of Anxiety and Depression
Contemporary neurobiological models of anxiety and depression include both distinct (anxiety and depression specific) and overlapping networks of brain regions. As displayed in
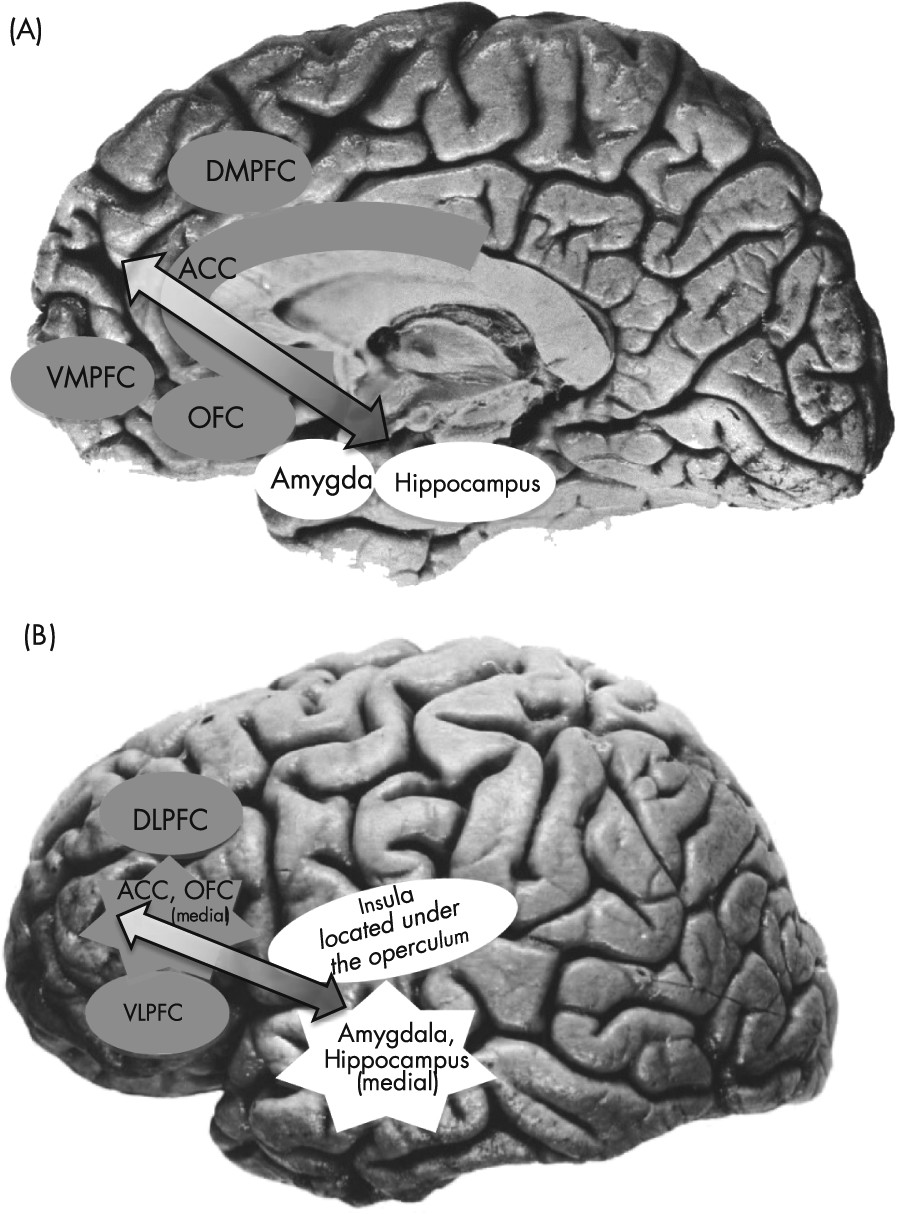
Figure 1, the neural circuitry of general emotion dysregulation and high negative affect, which is implicated in both types of disorders, includes an interconnected set of brain regions involved in the generation and regulation of emotion (
1–
3). Limbic structures (such as the amygdala, hippocampus, and insula) react to emotional information. Activity from these regions feeds forward through the anterior cingulate cortex ([ACC] involved in the appraisal and encoding of emotion), orbitofrontal cortex ([OFC] involved in the integration of affective and sensory information and reward processing), and finally to the dorsomedial and ventromedial prefrontal cortices ([DMPFC, VMPFC] involved in self-referential processing and in moderating emotional reactions). The initial activity in the limbic regions can be regulated by regions located within the prefrontal cortex (PFC). Lateral prefrontal regions, including the dorsolateral and ventrolateral prefrontal cortex ([DLPFC, VLPFC] both of which subserve higher-order cognitive functions), interact with the other frontal systems noted above, including the DMPFC, VMPFC, and ACC. These frontal systems are functionally interconnected with the amygdala and other limbic regions (
3) and can modulate limbic activity during controlled processing of emotional stimuli (
4).
The functioning of disorder-specific networks is also key to understanding the relationship between psychotherapy and brain function. In addition to the general emotion processing and emotion regulation networks described above, there is a partially overlapping set of regions that shows increased activation to fear-related stimuli. This system forms a “fear network,” and is particularly relevant to the anxious arousal and exaggerated fear responses that characterize anxiety disorders. This fear-responsive circuitry includes limbic regions such as the amygdala, hippocampus, and parahippocampal gyrus, as well as the insula, periaqueductal gray, and medial portions of the PFC (mPFC) including the VMPFC, OFC, and ACC (for a more detailed review, see de Carvalho et al. [
5]). Finally, the functioning of an additional network, the reward circuit, is particularly relevant in the treatment of major depression (
2), as it may play a role in anhedonia. This network of regions includes the ventral striatum, portions of the thalamus, amygdala, OFC, and mPFC (for a more detailed review, see Eshel and Roiser [
6]).
Hypothesized Neural Circuitry of Psychotherapy
One potential cause for many of the core symptoms of depression and anxiety, particularly those associated with negative emotional experiences, could be an inefficiency of top-down cortical control over regions that respond to emotional stimuli (e.g., limbic and fear-network-related regions). Psychotherapy has broadly been hypothesized to remediate these neural abnormalities and reduce symptoms via a strengthening of the cortical, top-down emotion regulatory processes (
Figure 1; see [
7,
8]). Under this theory, improved PFC and cortical function would lead to enhanced regulation over limbic regions, thereby attenuating emotional reactions to negative inputs. Basic research in healthy control subjects suggests that psychotherapy skills, such as problem solving, cognitive reappraisal, extinction learning (the critical learning mechanism in exposure therapy for anxiety), and modification of clients’ self-representations each rely on the function of PFC structures (e.g., DLPFC, VLPFC, VMPFC, ACC; reviewed in [
9]). This model therefore suggests that psychotherapy’s effects should involve improved regulation in PFC regions and corresponding modulation of regions responding to negative or threatening stimuli.
Whereas the neural models described above permit hypotheses regarding which regions of the brain are likely to be relevant in psychotherapy research, it is considerably more difficult to generate hypotheses regarding the expected direction of effects, e.g., increases versus decreases in activity. Following a review of the existing research findings, a discussion of the complexity in making directional hypotheses is provided.
General Discussion
The findings reviewed above are broadly consistent with predictions regarding the neural substrates of psychotherapy. Activity in regions associated with negative emotion, emotion regulation, fear, and reward are associated with a response to psychotherapy, and psychotherapy appears to alter the functioning of these regions. Beyond understanding which regions are involved, however, the state of the field has not yet evolved sufficiently to make many specific conclusions regarding the direction of these effects. Conflicting directional observations may, for example, be due to differences in task states (imaging during a resting state versus symptom provocation versus application of a specific therapy skill). Increased PFC function at rest may, in fact, contribute to decreased capacity for activation of the PFC in response to symptom provocation or skill application, leading to findings in opposing directions depending on the task state that is examined (
7). Furthermore, regional increases and decreases observed in neuroimaging are currently subject to multiple interpretations. Increased activation in a given region might be interpreted as reflecting an improvement in the strength of the region’s function, or as an impairment in the region’s efficiency, reflecting a need for greater activity in order to accomplish the same effect. One goal of future research will be to further clarify the precise nature of associations and to resolve the inconsistencies that have been observed.
The fact that brain function measured pretreatment is associated with the likelihood of response to psychotherapies is important. It suggests that future refinements regarding the precise direction of these effects across multiple kinds of tasks may enable treatments to be selected or individually tailored to the unique needs of the individual. Currently, several studies of psychotherapeutic treatment for anxiety have consistently implicated increased hyperresponsivity of regions reacting to threatening stimuli (e.g., the limbic and visual processing areas) as a marker of a better response to therapy. Such hyperreactivity may represent a clinically useful biomarker conferring a higher chance of a positive outcome from therapy, perhaps due to increased engagement with anxiety-provoking stimuli at baseline.
The pattern of predictive findings from studies of depression suggests that the functional state of the emotion regulation system, and potentially the reward system, prior to treatment has important consequences for the efficacy of cognitive-behavioral therapy. This may not be surprising. Cognitive-behavioral therapy is believed to engage and strengthen the patient’s ability to regulate and alter his or her emotional states. Little doubt remains that the ventral portions of the ACC play a key role in determining the likelihood of treatment response for depression. However, more work is needed to specify which specific subregions are critical, and what patterns of activity (to which tasks) are predictive of good or bad outcomes.
In order for brain-based predictive findings to become applicable in the clinic, greater clarity is needed regarding the precise task and imaging parameters that will lead to reproducible results at the single-patient level. Future work should aim to build on the strengths of recent studies, which used larger samples (
11,
15), randomization of participants to different treatments (
26), and more sophisticated analytic approaches that allow researchers to estimate the added benefit of neuroimaging data (
11), to examine patterns of communication between brain regions (
25), and to draw inferences that are valid at the individual patient level (
15). These advances will help to address questions such as
which treatment option is best for
which patient. Additionally, when multiple brain regions are observed to predict response, as in (
26), effort needs to be made to combine these multiple predictors in order to make a single treatment recommendation for the patient (see [
55] for a new method for doing so.)
Examining change in brain function over the course of treatment can be a valuable tool with which to understand the mechanisms through which a given treatment is operating. As others have noted (see
30,
56), the best designs for such studies would have at least three groups: the group receiving the treatment in question, a group of similar psychiatric patients who either receive a different treatment or no treatment, and a group of relatively well-matched healthy control subjects. With such a design, researchers would be able to determine whether or not the mechanisms observed are unique to the treatment under investigation, and whether the treatment involves normalization of function or the recruitment of compensatory systems. Additionally, researchers must examine and demonstrate that the psychometric properties of neuroimaging-derived markers, e.g., test-retest reliability, are sound (
57).
Currently, pre-post psychotherapy neuroimaging studies in anxiety disorders reveal some evidence consistent with hypothesized psychotherapy substrates (e.g., PFC increases, limbic decreases, and/or increased PFC-limbic functional connectivity). Evidence for increased activation in top-down PFC regions has been found most consistently in studies of posttraumatic stress disorder, whereas the findings in other anxiety disorders tend to suggest limbic (e.g., insular) decreases, either accompanied by concomitant PFC decreases (e.g., obsessive-compulsive disorder), or without consistent evidence of increases or decreases in regulatory regions.
Regarding changes during psychotherapy for depression, the most consistent findings are decreased activity across several regions of the PFC following multiple forms of psychotherapy, and reductions in negatively biased information processing in the PFC following cognitive-behavioral therapy. The fact that psychotherapy is broadly associated with decreased activity in several PFC regions may be surprising, and it appears to run counter to the hypothesis that psychotherapy should strengthen the ability of top-down control regions to modify the processing in down-stream systems. There is simply not yet enough information to resolve this issue, which is likely related to the methodological factors discussed above. It is also possible that the hypothesized mechanism of action of psychotherapy for depression is incorrect. More work will be needed to test these possibilities. The most informative studies will be those that test the core hypothesis that psychotherapy exerts effects via a top-down cortical route. If this hypothesis is true, psychotherapies should involve increasing communication between PFC and limbic areas, as evidenced by increased “functional connectivity” across these regions. To date, only one treatment study in depression has examined this issue (
25), and many more are needed to determine under what conditions psychotherapy can and cannot affect the functioning of the key neural circuits relevant to depression.
Methodological Challenges and Future Research
Although neuroimaging studies have tremendous potential to advance our understanding of psychotherapy process and outcomes, this area of research is relatively new. As such, the clinical relevance and applicability of findings to date are limited. Treatment outcome prediction studies have rarely included more than one treatment condition, and thus cannot differentiate between prescriptive effects (which would allow for the selection of the specific treatment most likely to work for a given patient, relative to other treatments) and prognostic effects (which indicate more generally whether treatment itself is likely to be effective). Similarly, treatment mechanism studies have often used methods that cannot adequately distinguish between mechanisms of change through which the therapy is exerting its specific effects, practice effects associated with repeated testing, and nonspecific neural correlates of symptom improvement.
A handful of more recent studies offer a glimpse of the cutting edge within this area of research, through the inclusion of larger, well-controlled samples (
26,
38,
41,
58), tasks designed to probe specific therapy skills (
38,
58), and explicit analysis of the relationship between neural change and treatment efficacy (e.g., symptom reduction [
41] or adherence [
58]). Additional research is also needed to examine the effects of specific components of treatment. One notable example is a study utilizing EEG measures of cortical activity to examine the association between brain function and cortical activity before and after a brief, 30-minute, cognitive-behavioral therapy analog training session (
59). Approaches like this may aid in identifying the neural mechanisms associated with specific therapeutic interventions. Finally, newer technologies, such as fNIRS, may provide researchers with additional tools with which to mitigate some of the methodological challenges noted above (
60). During fNIRS, participants wear skullcaps containing devices that emit and detect near-infrared light, with which changes in cortical blood flow can be examined. fNIRS frees participants from the confines of a magnetic scanner, and as such, allows participants to engage in tasks not possible during more standard neuroimaging protocols.
Neuroimaging in psychotherapy is an active and growing area of research. It holds tremendous promise for helping clinicians tailor specific interventions to the needs of individual patients, and it may help scientists determine how psychotherapeutic treatments work. By continuing to build on recent methodological advances, research in this area has the potential to fulfill its “bench-to-bedside” promise and improve clinical care across the full range of psychological suffering.