When historians of science turn their attention to the emergence of molecular medicine in the last half of the twentieth century, they will undoubtedly note the peculiar position occupied throughout this period by psychiatry. In the years following World War II, medicine was transformed from a practicing art into a scientific discipline based on molecular biology (
1). During that same period psychiatry was transformed from a medical discipline into a practicing therapeutic art. In the 1950s and in some academic centers extending into the 1960s, academic psychiatry transiently abandoned its roots in biology and experimental medicine and evolved into a psychoanalytically based and socially oriented discipline that was surprisingly unconcerned with the brain as an organ of mental activity.
This shift in emphasis had several causes. In the period after World War II, academic psychiatry began to assimilate the insights of psychoanalysis. These insights provided a new window on the richness of human mental processes and created an awareness that large parts of mental life, including some sources of psychopathology, are unconscious and not readily accessible to conscious introspection. Initially, these insights were applied primarily to what were then called neurotic illnesses and to some disorders of character. However, following the earlier lead of Eugene Bleuler (
2) and Carl Jung (
3), the reach of psychoanalytic therapy soon extended to encompass almost all of mental illness, including the major psychoses: schizophrenia and the major depressions (
4–
8).
Indeed, the extension of psychoanalytic psychiatry did not stop here; it next expanded to include specific
medical illnesses (
9,
10). Influenced in part by their experience in World War II, many psychiatrists came to believe that the therapeutic efficacy of psychoanalytic insights might solve not only the problems of mental illness but also otherwise intractable medical illnesses such as hypertension, asthma, gastric ulcers, and ulcerative colitis—diseases that did not readily respond to the pharmacological treatments available in the late 1940s. These illnesses were thought to be psychosomatic and to be induced by unconscious conflicts.
Thus, by 1960 psychoanalytically oriented psychiatry had become the prevailing model for understanding all mental and some physical illnesses. When in 1964 the Harvard Medical School celebrated the 20th year of the psychoanalytically oriented Department of Psychiatry at the Beth Israel Hospital, Ralph Kahana, a member of the faculty of that department, summarized the leadership role of psychoanalytically oriented psychiatry in the following way: “In the past 40 years, largely under the impact of psychoanalysis, dynamic psychotherapy has become the principal and essential curative skill of the American psychiatrist and, increasingly, a focus of his training” (
11).
By merging the descriptive psychiatry of the period before World War II with psychoanalysis, psychiatry gained a great deal in explanatory power and clinical insight. Unfortunately, this was achieved at the cost of weakening its ties with experimental medicine and with the rest of biology.
The drift away from biology was not due simply to changes in psychiatry; it was in part due to the slow maturation of the brain sciences. In the late 1940s the biology of the brain was neither technically nor conceptually mature enough to deal effectively with the biology of most higher mental processes and their disorders. The thinking about the relationship between brain and behavior was dominated by a view that different mental functions could not be localized to specific brain regions. This view was espoused by Karl Lashley (
12), who argued that the cerebral cortex was equipotential; all higher mental functions were presumed to be represented diffusely throughout the cortex. To most psychiatrists and even to many biologists, the notion of the equipotentiality of the cerebral cortex made behavior seem intractable to empirical biological analysis.
In fact, the separation of psychiatry from biology had its origins even earlier. When Sigmund Freud first explored the implications of unconscious mental processes for behavior (
13), he tried to adopt a neural model of behavior in an attempt to develop a scientific psychology. Because of the immaturity of brain science at the time, he abandoned this biological model for a purely mentalistic one based on verbal reports of subjective experiences. Similarly, in the 1930s B.F. Skinner (
14) rejected neurological theories in his studies of operant conditioning in favor of objective descriptions of observable acts.
Initially, this separation may have been as healthy for psychiatry as it was for psychology. It permitted the development of systematic definitions of behavior and of disease that were not contingent on still-vague correlations with neural mechanisms. Moreover, by incorporating the deep concern of psychoanalysis for the integrity of an individual's personal history, psychoanalytic psychiatry helped develop direct and respectful ways for physicians to interact with mentally ill patients, and it led to a less stigmatized social perspective on mental illness.
However, the initial separation of psychoanalysis from neural science advocated by Freud was stimulated by the realization that a merger was premature. As psychoanalysis evolved after Freud—from being an investigative approach limited to a small number of innovative thinkers to becoming the dominant theoretical framework in American psychiatry—the attitude toward neural science also changed. Rather than being seen as premature, the merger of psychoanalysis and biology was seen as unnecessary, because neural science was increasingly considered irrelevant.
Moreover, as the limitations of psychoanalysis as a system of rigorous, self-critical thought became apparent, rather than confronting these limitations in a systematic, questioning, experimental manner, and perhaps rejoining biology in searching for newer ways of exploring the brain, psychoanalytic psychiatry spent most of the decades of its dominance—the period from 1950 to 1980—on the defensive. Although there were important individual exceptions, as a group, psychoanalysts devalued experimental inquiry. Consequently, psychoanalysis slid into an intellectual decline that has had a deleterious effect on psychiatry, and because it discouraged new ways of thought, it has had a particularly deleterious effect on the training of psychiatrists.
Let me illustrate with a personal example the extent to which this unquestioning attitude came to influence my own psychiatry training. In the summer of 1960, I left my postdoctoral training in neural science at the National Institutes of Health (NIH) to begin residency training at the Massachusetts Mental Health Center, the major psychiatric teaching hospital of the Harvard Medical School. I entered training together with 20-odd other young physicians, many of whom went on to become leaders in American psychiatry: Judy Livant Rapaport, Anton Kris, Dan Buie, Ernst Hartmann, Paul Wender, Joseph Schildkraut, Alan Hobson, and George Vaillant. Yet in the several years in which this outstanding group of physicians was in training, at a time when training was leisurely and there was still a large amount of spare time, there were no required or even recommended readings. We were assigned no textbooks; rarely was there a reference to scientific papers in conferences or in case supervision. Even Freud's papers were not recommended reading for residents.
Much of this attitude came from our teachers, from the heads of the residency program. They made a point of encouraging us not to read. Reading, they argued, interfered with a resident's ability to listen to patients and therefore biased his or her perception of the patients' life histories. One famous, much quoted remark was that “there are those who care about people and there are those who care about research.” Through the efforts of the heads of the residency program, the whole thrust of psychoanalytic psychiatry at the Massachusetts Mental Health Center, and perhaps at the Harvard Medical School in general, was not simply to develop better psychiatrists but to develop better therapists—therapists prepared to understand and empathize with the patients' existential problems.
This view was summarized in 1978 by Day and Semrad (
7) in the following terms:
The essence of therapy with the schizophrenic patient is the interaction between the creative resources of both therapist and patient. The therapist must rely on his own life experience and translate his knowledge of therapeutic principles into meaningful interaction with the patient while recognizing, evoking, and expanding the patient's experience and creativity; both then learn and grow from the experience.
In order to engage a schizophrenic patient in therapy, the therapist's basic attitude must be an acceptance of the patient as he is—of his aims in life, his values, and his modes of operating, even when they are different and very often at odds with his own. Loving the patient as he is, in his state of decompensation, is the therapist's primary concern in approaching the patient. As a result the therapist must find his personal satisfactions elsewhere. His job is extremely taxing in its contradictions, for he must love the patient, expect him to change, and yet derive his additional satisfactions elsewhere and tolerate frustration.
In small measure this advice was sound, even in retrospect. A humane and compassionate perspective taught one to listen carefully and insightfully to one's patients. It helped us to develop the empathy essential for all aspects of a therapeutic relationship. But as a framework for a psychiatric education designed to train leaders in academic psychiatry, it was incomplete. For almost all residents it was intellectually limiting, and for some talented residents it proved stifling.
The almost unrealistic demand for empathy left little room for intellectual content. There were, for example, no grand rounds at the Massachusetts Mental Health Center. No outside speakers were invited to address the house officers on a regular basis to discuss current clinical or scientific issues. The major coordinated activity for the residents was a weekly group therapy session (with a wonderful and experienced group leader) in which the residents constituted the members of the group—the patients, so to speak.
It was only through the insistence of the house staff and their eagerness for knowledge that the first grand rounds were established at the Massachusetts Mental Health Center in 1965. To initiate these rounds, several of us tried to recruit a psychiatrist in the Boston area to speak about the genetic basis of mental illness. We could find no one; not a single psychiatrist in all of Boston was concerned with or even had thought seriously about that issue. We finally imposed on Ernst Mayr, the great Harvard biologist and a friend of Franz Kallmann, a founder of psychiatric genetics, to come and talk to us.
I am providing here an oversimplified description of the weakness of an environment that had many excellent qualities and many strengths. The intellectual quality of the house officers was remarkable, and the commitment of the faculty to the training of the house staff and to the treatment of the patients was admirable. Moreover, I am describing the predominant trend at the center; there were countervailing ones. While the heads of the training program actively discouraged both reading and research, the director of the center, Jack Ewalt, strongly encouraged research. Moreover, I have been assured that during this period Harvard psychiatry was remarkably out of step with the rest of the country, and that a lack of scholarly concern was not universal within academic psychiatry nationally. Clearly, scholarly concerns were not lacking at Washington University under Eli Robins, at a number of other centers in the midwest, or at Johns Hopkins University under Seymour Kety (
15). But a lack of critical questioning seemed to be widespread in Boston and at many other institutions on the east and west coasts of the country.
Our residency years—the decade of the 1960s—marked a turning point in American psychiatry. To begin with, new and effective treatments, in the form of psychopharmacological drugs, began to be available. Initially, a number of supervisors discouraged us from using them, believing that they were designed more to aid our anxiety than that of the patients. By the mid-1970s the therapeutic scene had changed so dramatically that psychiatry was forced to confront neural science if only to understand how specific pharmaco~logical treatments were working.
With the advent of psychopharmacology, psychiatry was changed, and that change brought it back into the mainstream of academic medicine. There were three components to this progress. First, whereas psychiatry once had the least effective therapeutic armamentarium in medicine, it now had effective treatments for the major mental illnesses and something that began to approach a practical cure for two of the three most devastating diseases: depression and manic-depressive illness. Second, led first by Eli Robins at Washington University and then by Robert Spitzer at Columbia University's New York State Psychiatric Institute, new clinically validated and objective criteria were established for diagnosing mental illness. Third, Seymour Kety used his leadership position at NIH to spark a renewed interest in the biology of mental illness and specifically in the genetics of schizophrenia and depression.
In parallel, the years since 1980 have witnessed major developments in brain sciences, in particular in the analysis of how different aspects of mental functioning are represented by different regions of the brain. Thus, psychiatry is now presented with a new and unique opportunity. When it comes to studying mental function, biologists are badly in need of guidance. It is here that psychiatry, and cognitive psychology, as guide and tutor, can make a particularly valuable contribution to brain science. One of the powers of psychiatry, of cognitive psychology, and of psychoanalysis lies in their perspectives. Psychiatry, cognitive psychology, and psychoanalysis can define for biology the mental functions that need to be studied for a meaningful and sophisticated understanding of the biology of the human mind. In this interaction, psychiatry can play a double role. First, it can seek answers to questions on its own level, questions related to the diagnosis and treatment of mental disorders. Second, it can pose the behavioral questions that biology needs to answer if we are to have a realistically advanced understanding of human higher mental processes.
A COMMON FRAMEWORK FOR PSYCHIATRY AND THE NEURAL SCIENCES
As a result of advances in neural science in the last several years, both psychiatry and neural science are in a new and better position for a rapprochement, a rapprochement that would allow the insights of the psychoanalytic perspective to inform the search for a deeper understanding of the biological basis of behavior. As a first step toward such a rapprochement, I here outline an intellectual framework designed to align current psychiatric thinking and the training of future practitioners with modern biology.
This framework can be summarized in five principles that constitute, in simplified form, the current thinking of biologists about the relationship of mind to brain.
Principle 1. All mental processes, even the most complex psychological processes, derive from operations of the brain. The central tenet of this view is that what we commonly call mind is a range of functions carried out by the brain. The actions of the brain underlie not only relatively simple motor behaviors, such as walking and eating, but all of the complex cognitive actions, conscious and unconscious, that we associate with specifically human behavior, such as thinking, speaking, and creating works of literature, music, and art. As a corollary, behavioral disorders that characterize psychiatric illness are disturbances of brain function, even in those cases where the causes of the disturbances are clearly environmental in origin.
Principle 2. Genes and their protein products are important determinants of the pattern of interconnections between neurons in the brain and the details of their functioning. Genes, and specifically combinations of genes, therefore exert a significant control over behavior. As a corollary, one component contributing to the development of major mental illnesses is genetic.
Principle 3. Altered genes do not, by themselves, explain all of the variance of a given major mental illness. Social or developmental factors also contribute very importantly. Just as combinations of genes contribute to behavior, including social behavior, so can behavior and social factors exert actions on the brain by feeding back upon it to modify the expression of genes and thus the function of nerve cells. Learning, including learning that results in dysfunctional behavior, produces alterations in gene expression. Thus all of “nurture” is ultimately expressed as “nature.”
Principle 4. Alterations in gene expression induced by learning give rise to changes in patterns of neuronal connections. These changes not only contribute to the biological basis of individuality but presumably are responsible for initiating and maintaining abnormalities of behavior that are induced by social contingencies.
Principle 5. Insofar as psychotherapy or counseling is effective and produces long-term changes in behavior, it presumably does so through learning, by producing changes in gene expression that alter the strength of synaptic connections and structural changes that alter the anatomical pattern of interconnections between nerve cells of the brain. As the resolution of brain imaging increases, it should eventually permit quantitative evaluation of the outcome of psychotherapy.
I now consider each of these principles in turn and illustrate the experimental basis of this new framework and its implications for the theory and practice of psychiatry.
All Functions of Mind Reflect Functions of Brain
This principle is so central in traditional thinking in biology and medicine (and has been so for a century) that it is almost a truism and hardly needs restatement. This principle stands as the basic assumption underlying neural science, an assumption for which there is enormous scientific support. Specific lesions of the brain produce specific alterations in behavior, and specific alterations in behavior are reflected in characteristic functional changes in the brain (
16). Nevertheless, two points deserve emphasis.
First, although this principle is now accepted among biologists, the details of the relationship between the brain and mental processes—precisely how the brain gives rise to various mental processes—is understood poorly, and only in outline. The great challenge for biology and psychiatry at this point is to delineate that relationship in terms that are satisfying to both the biologist of the brain and the psychiatrist of the mind.
Second, the relationship of mind to brain becomes less obvious, more nuanced, and perhaps more controversial when we appreciate that biologists apply this principle to all aspects of behavior, from our most private thoughts to our most public expression of emotion. The principle applies to behaviors by single individuals, to behaviors between individuals, and to social behavior in groups of individuals. Viewed in this way, all sociology must to some degree be sociobiology; social processes must, at some level, reflect biological functions. I hasten to add that formulating a relationship between social processes (or even psychological processes) and biological functions might not necessarily prove to be optimally insightful in elucidating social dynamics. For many aspects of group or individual behavior, a biological analysis might not prove to be the optimal level or even an informative level of analysis, much as subatomic resolution is often not the optimal level for the analysis of biological problems. Nevertheless, it is important to appreciate that there are critical biological underpinnings to all social actions.
This aspect of the principle has not been readily accepted by all, especially not by all sociologists, as can be illustrated by one example from the Center for Advanced Studies in the Behavioral Sciences in Palo Alto, Calif., probably the country's premier think tank in the social sciences. In its annual report of 1996, the center described the planning of a special project entitled Culture, Mind, and Biology. As plans for this project pro~gressed, it became clear that many social scientists had a deep and enduring antipathy toward the biological sciences because they equated biological thinking with a view of human nature that they found simplistic, misguided, and socially and ethically dangerous. Since two earlier and influential biological approaches to the social sciences—scientifically argued racism and social Darwinism—had proven to be intellectually sterile and socially destructive, many social scientists objected to the idea. They objected to the notion
that a living organism's properties (not only its physical form but also its behavioral inclinations, abilities, and life prospects) are material and hence reducible to its genes. The conception of human nature that many social scientists associate with biological thinking asserts that individual and group differences as well as individual and group similarities in physical form, behavioral inclination, abilities, and life prospects can similarly be understood and explained by genesAs a result of this understanding, many disclaim the relevance of biological thinking for behavior and instead embrace some type of radical mind-body dualism in which it is assumed that the processes and products of the mind have very little to do with the processes and products of the body [my italics].
What is the basis of this unease among social scientists? Like all knowledge, biological knowledge is a double-edged sword. It can be used for ill as well as for good, for private profit or public benefit. In the hands of the misinformed or the malevolent, natural selection was distorted to social Darwinism, and genetics was corrupted into eugenics. Brain sciences have also been and can again be misused for social control and manipulation. How can we ensure that the advances of the brain sciences will serve to enrich our lives and to elevate our understanding of ourselves and each other? The only way to encourage the responsible use of this knowledge is to base the uses of biology in social policy on an understanding of biology.
The unease of social scientists derives in part from two misapprehensions (not unique to social scientists): first, that biologists think that biological processes are strictly determined by genes and, second, that the sole function of genes is the inexorable transmission of hereditary information from one generation to another. These profoundly wrong ideas lead to the notion that invariant, unregulated genes, not modifiable by external events, exert an inevitable influence on the behavior of individuals and their progeny. In this view, social forces as such have little influence on human behavior. They are powerless in the face of the predetermined, relentless actions of the genes.
This fatalistic and fundamentally wrong view was behind the eugenics movements of the 1920s and 1930s. As a basis for social policy, this view justifiably elicits fear and distrust in clear-thinking people. However, this view is based on a fundamental misconception of how genes work, which even some psychiatrists may not fully appreciate. The key concept of importance here is that genes have dual functions.
First, genes serve as stable templates that can replicate reliably. This template function is exercised by each gene, in each cell of the body, including the gametes. It is this function that provides succeeding generations with copies of each gene. The fidelity of the template replication is high. Moreover, the template is not regulated by social experience of any sort. It can only be altered by mutations, and these are rare and often random. This function of the gene, its template (transmission) function, is indeed beyond our individual or social control.
Second, genes determine the phenotype; they determine the structure, function, and other biological characteristics of the cell in which they are expressed. This second function of the gene is referred to as its transcriptional function. Although almost every cell of the body has all of the genes that are present in every other cell, in any given cell type (be it a liver cell or a brain cell) only a fraction of genes, perhaps 10%–20%, are expressed (transcribed). All of the other genes are effectively repressed. A liver cell is a liver cell and a brain cell is a brain cell because each of these cell types expresses only a particular subset of the total population of genes. When a gene is expressed in a cell, it directs the phenotype of that cell: the manufacture of specific proteins that specify the character of that cell.
Whereas the template function, the sequence of a gene—and the ability of the organism to replicate that sequence—is not affected by environmental experience, the transcriptional function of a gene—the ability of a given gene to direct the manufacture of specific proteins in any given cell—is, in fact, highly regulated, and this regulation is responsive to environmental factors.
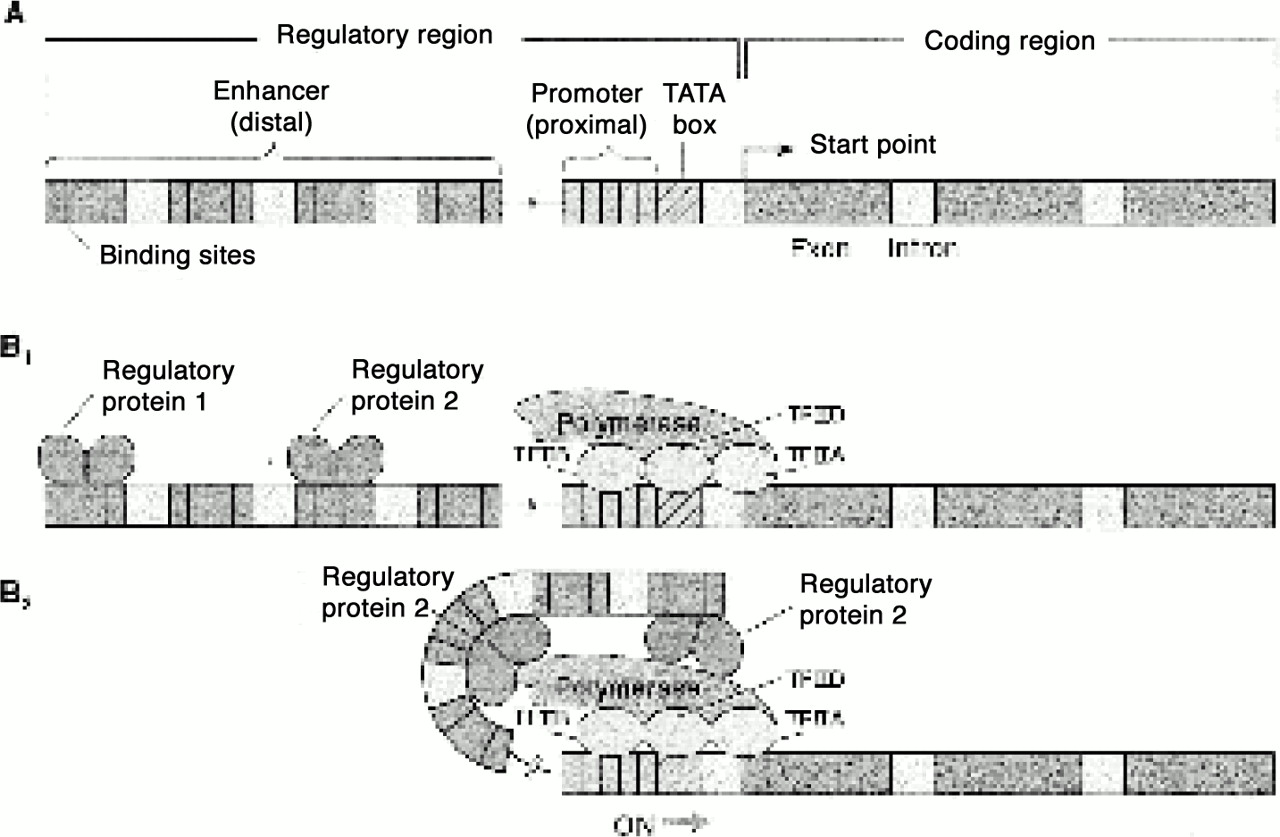
A gene has two regions (
figure 1). A coding region encodes mRNA, which in turn encodes a specific protein. A regulatory region usually lies upstream of the coding region and consists of two DNA elements. The
promoter element is a site where an enzyme, called RNA polymerase, will begin to read and transcribe the DNA coding region into mRNA. The
enhancer element recognizes protein signals that determine in which cells, and when, the coding region will be transcribed by the polymerase. Thus, a small number of proteins, or transcriptional regulators, that bind to different segments of the enhancer element determine how often RNA polymerase binds to the promoter element and transcribes the gene. Internal and external stimuli—steps in the development of the brain, hormones, stress, learning, and social interaction—alter the binding of the transcriptional regulators to the enhancer element, and in this way different combinations of transcriptional regulators are recruited. This aspect of gene regulation is sometimes referred to as
epigenetic regulation.
Stated simply, the regulation of gene expression by social factors makes all bodily functions, including all functions of the brain, susceptible to social influences. These social influences will be biologically incorporated in the altered expressions of specific genes in specific nerve cells of specific regions of the brain. These socially influenced alterations are transmitted culturally. They are not incorporated in the sperm and egg and therefore are not transmitted genetically. In humans the modifiability of gene expression through learning (in a nontransmissible way) is particularly effective and has led to a new kind of evolution: cultural evolution. The capability of learning is so highly developed in humans that humankind changes much more by cultural evolution than by biological evolution. Measurements of skulls found in the fossil record suggest that the size of the human brain has not changed since Homo sapiens first appeared approximately 50,000 years ago; yet clearly, human culture has evolved dramatically in that same time.
Genes Contribute Importantly to Mental Function and Can Contribute to Mental Illness
Let us consider the contribution of the template functions of DNA—the heritable aspects of gene action. Here we first need to ask, How do genes contribute to behavior? Clearly, genes do not code for behavior in a direct way. A single gene encodes a single protein; it cannot by itself encode for a single behavior. Behavior is generated by neural circuits that involve many cells, each of which expresses specific genes that direct the production of specific proteins. The genes expressed in the brain encode proteins that are important in one or another step of the development, maintenance, and regulation of the neural circuits that underlie behavior. A wide variety of proteins—structural, regulatory, and catalytic—are required for the differentiation of a single nerve cell, and many cells and many more genes are required for the development and function of a neural circuit.
To account for what we now appreciate as variations in the template functions of a gene, Darwin and his followers first postulated that variations in human behavior may, in part, be due to natural selection. If this is so, some element of the behavioral variation in any population will necessarily have a genetic basis. Some portion of this variation in turn should show up as clearly heritable differences. Control studies of heritable factors in human behavior have proven difficult to devise, because it is not possible or desirable to control an individual's environment for experimental purposes except in some very limited situations. Thus, behavioral studies of identical twins provide important information not otherwise available.
Identical twins share an identical genome and are therefore as alike genetically as is possible for two individuals. Similarities between identical twins who have been separated early in life and raised in different households, as occasionally happens, will therefore be more attributable to genes than to environment. Identical twins, compared with a group of individuals matched in age, sex, and socioeconomic status, share a remarkable number of behavioral traits. These include tastes, religious preferences, and vocational interests that are commonly considered to be socially determined and distinctive features of an individual. These findings argue that human behavior has a significant hereditary component. But the similarity is far from perfect. Twins can and do vary a great deal. Thus, twin studies also emphasize the importance of environmental influences; they indicate quite clearly that environmental factors are very important (
16).
A similar situation applies to disturbances of behavior and to mental illness. The first direct evidence that genes are important in the development of schizophrenia was provided in the 1930s by Franz Kallmann (
18). Kallmann was impressed with the fact that the incidence of schizophrenia throughout the world is uniformly about 1%, even though the social and environmental factors vary dramatically. Nevertheless, he found that the incidence of schizophrenia among parents, children, and siblings of patients with the disease is 15%, strong evidence that the disease runs in families. However, a genetic basis for schizophrenia cannot simply be inferred from the increased incidence in families. Not all conditions that run in families are necessarily genetic: wealth and poverty, habits, and values also run in families, and in earlier times even nutritional deficiencies such as pellagra ran in families.
To distinguish genetic from environmental factors, Kallmann turned to twin studies and compared the rates of illness in identical (monozygotic) and fraternal (dizygotic) twins. As we have seen, monozygotic twins share almost all of each other's genes. By contrast, dizygotic twins share only 50% of their genes and are genetically equivalent to siblings. Therefore, if schizophrenia is caused entirely by genetic factors, monozygotic twins should be identical in their tendency to develop the disease. Even if genetic factors were necessary but not sufficient for the development of schizophrenia, because environmental factors were involved, a mono~zygotic twin of a patient with schizophrenia should be at substantially higher risk than a dizygotic twin. The tendency for twins to have the same illness is called concordance. Studies on twins have established that the concordance for schizophrenia in mono~zygotic twins is about 45%, compared to only about 15% in dizygotic twins, which is about the same as for other siblings.
To disentangle further the effects of nature and nurture, Heston (
19) studied patients in the United States and Rosenthal and colleagues (
20) studied patients in Denmark. In both sets of studies, the rate of schizophrenia was higher among the biological relatives of adopted children who had schizophrenia than among those of adopted children who were normal. The difference in rate, about 10%–15%, was the same as that observed earlier by Kallman.
This familial pattern of schizophrenia is most dramatically evident in an analysis of the data from Denmark by Gottesman (
21). Gottesman examined the data from 40 Danish patients with schizophrenia, identifying all relatives with schizophrenia for whom good family pedigrees were available. He then ranked the relatives in terms of the percentage of genes shared with the schizophrenic patient. He found a higher incidence of schizophrenia among first-order relatives—those who share 50% of the patient's genes, including siblings, parents, and children—than among second-order relatives—those who share 25% of the patient's genes, including aunts, uncles, nieces, nephews, and grandchildren. Even the third-degree relatives, who share only 12.5% of the patient's genes, had a higher incidence of schizophrenia than the 1% found in the population at large. These data strongly suggest a genetic contribution to schizophrenia.
If schizophrenia were caused entirely by genetic abnormalities, the concordance rate for monozygotic twins, who share almost all of each other's genes, would be nearly 100%. The fact that the rate is 45% clearly indicates that genetic factors are not the only cause. Multiple causality is also evident from studies of the genetic transmission of the disease. Relatively routine studies of pedigrees are sufficient to pinpoint whether a disease is transmitted by dominant or recessive Mendelian inheritance, but this has not proven to be the mode of transmission of schizophrenia. The most likely explanation for the unusual genetic transmission of schizophrenia is that it is a multigenic disease involving allelic variations in perhaps as many as 10–15 loci in the population worldwide, and that perhaps combinations of three to five loci are needed to cause the disease in an individual. Moreover, these several genes can vary in the degree of penetrance.
In a natural population, any gene at any locus will exist in a number of different, clearly related forms called alleles. The penetrance of an allele depends on the interaction between that allele and the remainder of the genome, as well as with environmental factors. One twin can inherit a set of genes that program tall growth, but without good nutrition that twin may never grow tall. Similarly, not all people with the same dominant and abnormal Huntington's disease gene will have the full-blown movement disorders and accompanying cognitive disturbances; a few may have a more moderate form of the disease.
As in other polygenic diseases, such as diabetes and hypertension, most forms of schizophrenia are thought to require not only the accumulation of several genetic defects but also the actions of developmental and environmental factors. To understand schizophrenia it will be essential to learn how several genes combine to predispose an individual to a disease and to determine how the environment influences the expression of these genes.
The fact that many genes are involved does not mean, however, that in some cases single genes are not essential for the expression of a behavior. The importance of specific genes to behavior can best be demonstrated in simple animals, such as fruit flies or mice, in which mutations in a single gene can be more easily studied. Mutations of single genes in Drosophila or in mice can produce abnormalities in a variety of behaviors, including learned behavior as well as innate behavior such as courtship and locomotion.
Behavior Itself Can Also Modify Gene Expression
I have considered the template function of the gene, which is transmissible but not regulated. I now turn to that aspect of genetic function that is regulated but not transmitted. Studies of learning in simple animals provided the first evidence that experience produces sustained changes in the effectiveness of neural connections by altering gene expression. This finding has profound ramifications that should revise our view of the relationship between social and biological processes in the shaping of behavior.
To appreciate the importance of this relationship, consider for a moment the situation in American psychiatry as recently as 1968, when DSM-II appeared. A common view in psychiatry at that time was that biological and social determinants of behavior act on separate levels of the mind: one level had a clear empirical basis, and the other was unspecified. As a result, until the 1970s psychiatric illnesses were traditionally classified into two major categories: organic and functional. Thus, Seltzer and Frazier (
22) wrote in 1978, “Organic brain syndrome is a general term used to describe those conditions of impaired function of the nervous system that are manifest by psychiatric symptoms. This contrasts with the majority of psychiatric syndromes called `functional.'”
These organic mental illnesses included the dementias, such as Alzheimer's disease, and the toxic psychoses, such as those that follow the chronic use of cocaine, heroin, and alcohol. Functional mental illnesses included not only the neurotic illnesses but also the depressive illnesses and the schizophrenias.
This distinction originally derived from the observations of nineteenth-century neuropathologists, who examined the brains of patients at autopsy and found gross and readily demonstrable distortions in the architecture of the brain in some psychiatric diseases but not in others. Diseases that produced anatomical evidence of brain lesions were called organic; those lacking these features were called functional.
This distinction, now clearly outdated, is no longer tenable. There can be no changes in behavior that are not reflected in the nervous system and no persistent changes in the nervous system that are not reflected in structural changes on some level of resolution. Everyday sensory experience, sensory deprivation, and learning can probably lead to a weakening of synaptic connections in some circumstances and a strengthening of connections in others. We no longer think that only certain diseases, the organic diseases, affect mentation through biological changes in the brain and that others, the functional diseases, do not. The basis of the new intellectual framework for psychiatry is that all mental processes are biological, and therefore any alteration in those processes is necessarily organic.
As is now evident in DSM-IV, the classification of mental disorders must be based on criteria other than the presence or absence of
gross anatomical abnormalities. The absence of detectable structural changes does not rule out the possibility that more subtle but nonetheless important biological changes are occurring. These changes may simply be below the level of detection with the still-limited techniques available today. Demonstrating the biological nature of mental functioning requires more sophisticated anatomical methodologies than the light-microscopic histology of nineteenth-century pathologists. To clarify these issues it will be necessary to develop a neuropathology of mental illness that is based on anatomical function as well as anatomical structure. Imaging techniques such as positron emission tomography and functional magnetic resonance imaging have opened the door to the noninvasive exploration of the human brain at a level of resolution that begins to approach that which is required to understand the physical mechanisms of mentation and therefore of mental disorders. This approach is now being pursued in the study of schizophrenia, depression, obsessive-compulsive disorders, and anxiety disorders (
23).
We now need to ask, How do the biological processes of the brain give rise to mental events, and how in turn do social factors modulate the biological structure of the brain? In the attempt to understand a particular mental illness, it is more appropriate to ask, To what degree is this biological process determined by genetic and developmental factors? To what degree is it environmentally or socially determined? To what degree is it determined by a toxic or infectious agent? Even the mental disturbances that are considered to be most heavily determined by social factors must have a biological component, since it is the activity of the brain that is being modified.
A New View of the Relationship Between Inherited and Acquired Mental Illnesses
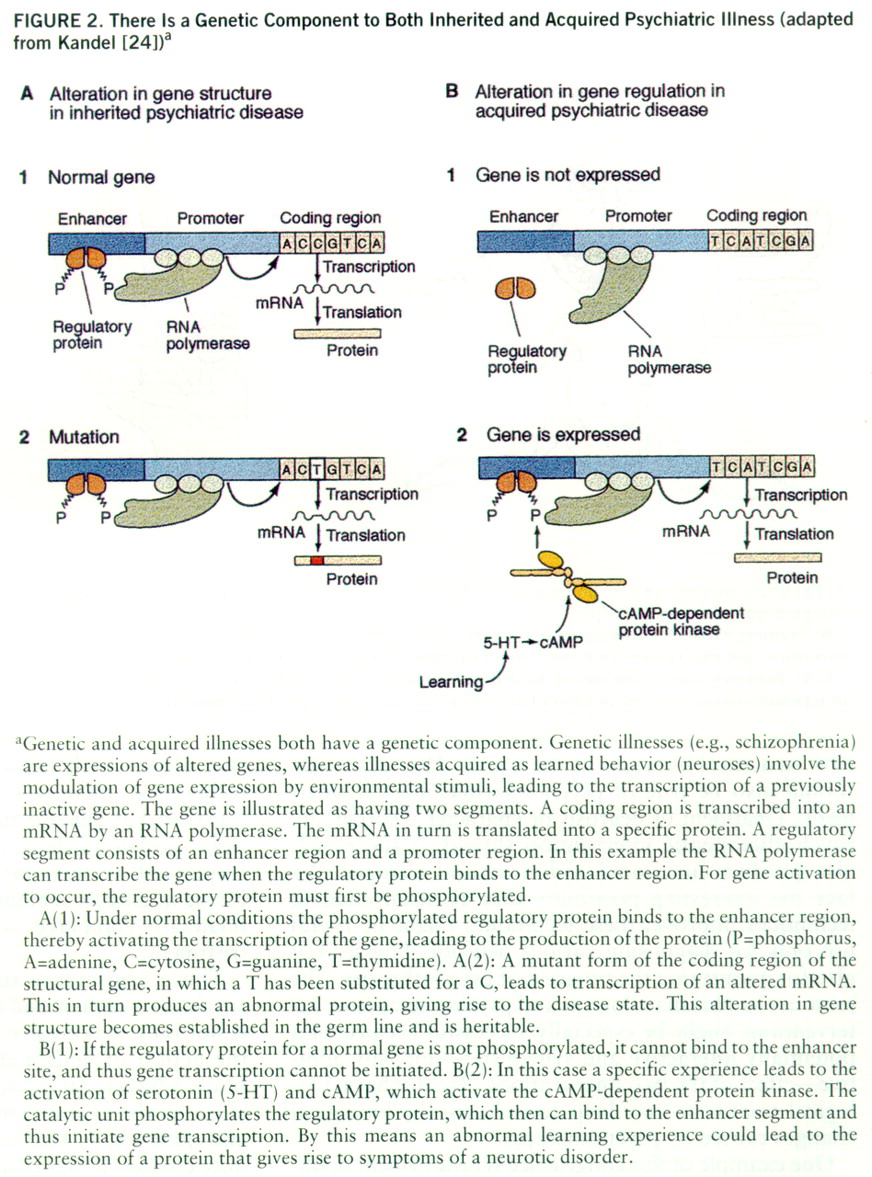
In the few instances where it has been possible to examine rigorously the persistent changes in mental functions, these functions have been shown to involve alterations in gene expression. Thus, in studying the specific changes that underlie persistent mental states, normal as well as disturbed, we should also look for altered gene expression. As we have seen, there is now substantial evidence that the susceptibility to major psychotic illnesses (schizophrenia and manic-depressive disorders) is heritable. These illnesses in part reflect alterations in the template function of the gene—in the nucleotide sequence of a number of different genes—leading to abnormal mRNAs and abnormal proteins. It is therefore tempting to think that insofar as psychiatric illnesses such as posttraumatic stress syndrome are acquired by experience, they are likely to involve alterations in the transcriptional function of the gene—in the regulation of gene expression. Nonetheless, some individuals may be much more susceptible to this syndrome because of the combination of genes they have inherited.
Development, stress, and social experience are all factors that can alter gene expression by modifying the binding of transcriptional regulators to each other and to the regulatory regions of genes. It is likely that at least some neurotic illnesses (or components of them) result from reversible defects in gene regulation, which may be due to altered binding of specific proteins to certain upstream regions that control the expression of certain genes (
figure 2).
Maintenance of Learned Alterations in Gene Expression by Structural Alterations in Neural Circuits of the Brain
How does altered gene expression lead to the stable alterations of a mental process? Animal studies of alterations in gene expression induced by learning indicate that one major consequence of such alterations in gene activation is the growth of synaptic connections. This growth was first delineated by studies in simple invertebrate animals such as the snail
Aplysia (
25). Animals subjected to controlled learning that gave rise to long-term memory had twice as many presynaptic terminals as untrained animals. Some forms of learning, such as long-term habituation, produce the opposite changes; they lead to a regression and pruning of synaptic connections. These morphological changes seem to be a signature of the long-term memory process. They do not occur with short-term memory.
In mammals, and especially in humans, each functional component of the nervous system is represented by hundreds of thousands of nerve cells. In such complex systems a specific instance of learning is likely to lead to alterations in a large number of nerve cells insofar as the interconnections of the various sensory and motor systems involved in the learning are changed. Indeed, studies have shown that such vast changes do occur. The most detailed evidence has come from studies of the somatic sensory system.
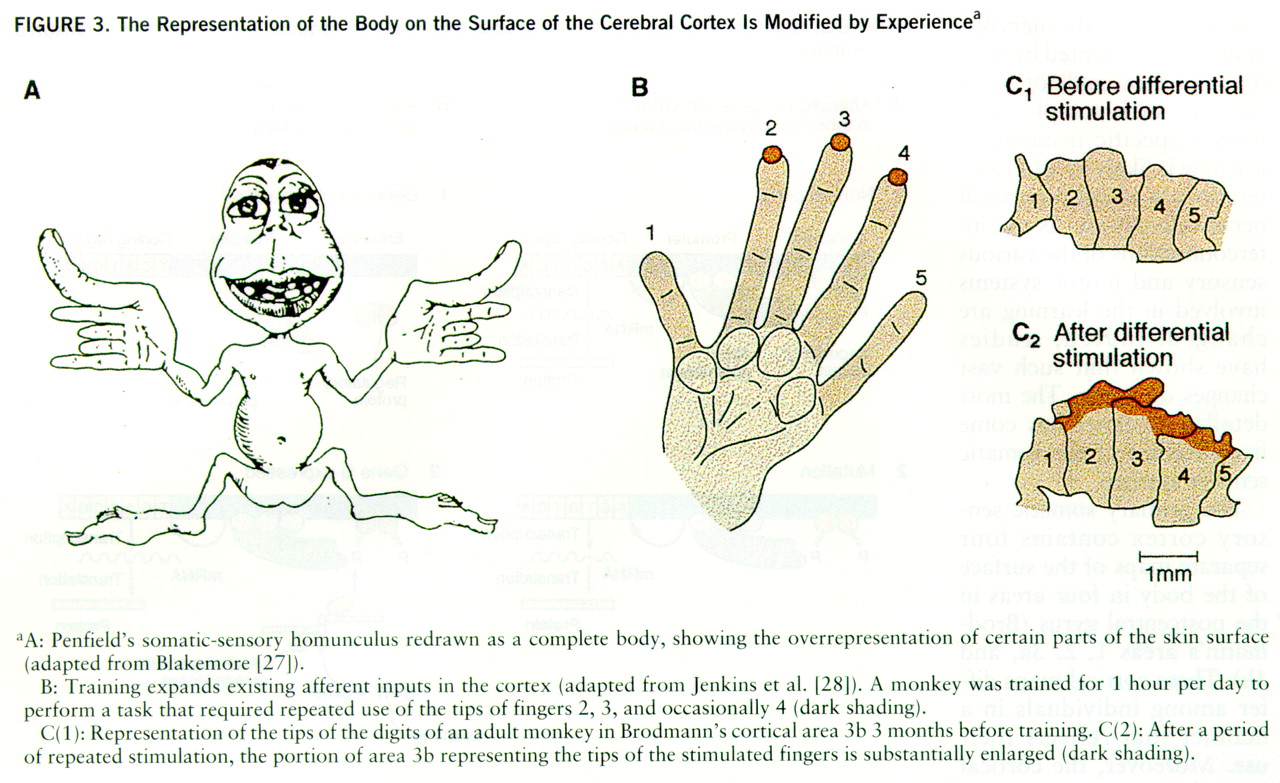
The primary somatic sensory cortex contains four separate maps of the surface of the body in four areas in the postcentral gyrus (Brodmann's areas 1, 2, 3a, and 3b). These cortical maps differ among individuals in a manner that reflects their use. Moreover, the cortical maps for somatic sensations are dynamic, not static, even in mature animals (
26). The distribution of these functional connections can expand and retract, depending on the particular uses or activities of the peripheral sensory pathways. Since each of us is brought up in a somewhat different environment, exposed to different combinations of stimuli, and we develop motor skills in different ways, each brain is modified in unique ways. This distinctive modification of brain architecture, along with a unique genetic makeup, constitutes the biological basis for individuality.
Two studies provide evidence for this view (
26). One study found that the somatosensory maps vary considerably among normal animals. However, this study did not separate the effects of different experiences from the consequences of different genetic endowment. Another study was conducted to see whether activity is important in determining the topographic organization of the somatosensory cortex. Adult monkeys were encouraged to use three middle fingers at the expense of two other fingers of the hand to obtain food. After several thousand trials, the area of cortex devoted to the three fingers was greatly expanded at the expense of the area normally devoted to the other fingers (
figure 3). Practice alone, therefore, may not only strengthen the effectiveness of existing patterns of connections, but also change cortical connections to accommodate new patterns of actions.
PSYCHOTHERAPY AND PHARMACOTHERAPY MAY INDUCE SIMILAR ALTERATIONS IN GENE EXPRESSION AND STRUCTURAL CHANGES IN THE BRAIN
As these arguments make clear, it is intriguing to suggest that insofar as psychotherapy is successful in bringing about substantive changes in behavior, it does so by producing alterations in gene expression that produce new structural changes in the brain. This obviously should also be true of psychopharmacological treatment. Treatment of neurosis or character disorders by psychotherapeutic intervention should, if successful, also produce functional and structural changes. We face the interesting possibility that as brain imaging techniques improve, these techniques might be useful not only for diagnosing various neurotic illnesses but also for monitoring the progress of psychotherapy. The joint use of pharmacological and psychotherapeutic interventions might be especially successful because of a potentially interactive and synergistic—not only additive—effect of the two interventions. Psychopharmacological treatment may help consolidate the biological changes caused by psychotherapy.
One example of this congruence is now evident in obsessive-compulsive disorder (OCD). This common debilitating psychiatric illness is characterized by recurrent unwanted thoughts, obsessions, and conscious ritualized acts and compulsions that are usually attributed to attempts to deal with the anxiety generated by the obsessions. Medications that are selective serotonin reuptake inhibitors (SSRIs) and specific behavioral therapies that use the principles of deconditioning, involving exposure and response prevention, are effective in reducing the symptoms of many patients with OCD.
Many investigators have postulated a role for the cortical-striatal-thalamic brain system in the mediation of OCD symptoms. OCD is associated with functional hyperactivity of the head of the right caudate nucleus. After effective treatment of OCD with either an SSRI (such as fluoxetine) alone or with behavioral modification alone (with exposure and response prevention techniques), there is a substantial decrease in activity (measured as glucose metabolic rate) in the head of the right caudate nucleus. In one study (
29) patients who responded to behavior therapy had a significant decrease in glucose metabolic rate in the caudate nucleus bilaterally compared to those who did not respond to treatment.
These arguments suggest that when a therapist speaks to a patient and the patient listens, the therapist is not only making eye contact and voice contact, but the action of neuronal machinery in the therapist's brain is having an indirect and, one hopes, long-lasting effect on the neuronal machinery in the patient's brain; and quite likely, vice versa. Insofar as our words produce changes in our patient's mind, it is likely that these psychotherapeutic interventions produce changes in the patient's brain. From this perspective, the biological and sociopsychological approaches are joined.
IMPLICATIONS OF A NEW FRAMEWORK FOR THE PRACTICE OF PSYCHIATRY
The biological framework that I have outlined here is not only important conceptually; it is also important practically. To function effectively in the future, the psychiatrists we are training today will need more than just a nodding familiarity with the biology of the brain. They will need the knowledge of an expert, a knowledge perhaps different from but fully comparable to that of a well-trained neurologist. In fact, it is likely that in the decades ahead we will see a new level of cooperation between neurology and psychiatry. This cooperation is likely to have its greatest impact on patients for whom the two approaches—neurological and psychiatric—overlap, such as those in treatment for autism, mental retardation, and the cognitive disorders due to Alzheimer's and Parkinson's diseases.
It can be argued that an intellectual framework so fully embedded in biology and aligned with neurology is premature for psychiatry. In fact, we are only beginning to understand the simplest mental functions in biological terms; we are far from having a realist neurobiology of clinical syndromes and even farther from having a neurobiology of psychotherapy. These arguments have some validity. Thus, the decision for psychiatry revolves around the question, When will the time be optimal for a more complete rapprochement between psychiatry and biology? Is it when the problem is still premature—when the biology of mental illness still confronts us as deep mysteries—or is it when the problem is already postmature—when mental illness is on the way to being understood? If psychiatry will join the intellectual fray in full force only when the problems are largely solved, then psychiatry will deprive itself of one of its main functions, which is to provide leadership in the attempts to understand the basic mechanisms of mental processes and their disorders. Since the presumed function of academic psychiatry is to train people who advance knowledge—people who can not only benefit from the insights of the current biological revolution but also contribute to it—psychiatry must take its commitment to the training of biological scientists more seriously. It must put its own oars into the water and pull its own weight. If the biology of mental processes continues to be solved by others without the active participation of psychiatrists, we may well ask, What is the purpose of a psychiatric education?
While psychiatrists debate the degree to which they should immerse themselves in modern molecular biology, most of the remaining scientific community has resolved that issue for itself. Most biologists sense that we are in the midst of a remarkable scientific revolution, a revolution that is transforming our understanding of life's processes—the nature of disease and of medical therapeutics. Most biologists believe that this revolution will have a profound impact on our understanding of mind. This view is shared by students just beginning their scientific training. Many of the very best graduate students in biology and the best M.D.-Ph.D. students are drawn to neural science and particularly to the biology of mental processes for this very reason. If the progress of the past few years and the continued influx of talented people is any guide, we can expect a major growth in our understanding of mental processes.
We thus are confronting an interesting paradox. While the scientific community at large has become interested in the biology of mental processes, the interest of medical students in a psychiatric career is declining. Thus, from an educational point of view, psychiatry is in a trough. One reason for the loss of interest, beyond the economic issue of managed care, is the current intellectual scene in psychiatry. Medical students realize that insofar as the teaching of psychiatry is often based primarily on doing psychotherapy, a major component of psychiatry as it is now taught does not require a medical education. As Freud so clearly emphasized, psychotherapy can be carried out effectively by nonmedical specialists. Why, then, go to medical school?
As a greater emphasis on biology begins to change the nature of psychiatry, it also is likely to draw an increasing number of talented medical students into psychiatry. In addition, it will make psychiatry a more technologically sophisticated and more scientifically rigorous medical discipline. A biological orientation can help revitalize the teaching and practice of psychiatry by bringing to bear on the problems of mental illness a critical understanding of brain processes, a familiarity with therapeutics, and an understanding of both neurological and psychiatric diseases—in short, an ability to encompass mental and emotional life within a framework that includes biological as well as social determinants. A renewed involvement of psychiatry with biology and with neurology, therefore, not only is scientifically important but also emphasizes the scientific competence that, I would argue, should be the basis for the clinical specialty of psychiatry in the twenty-first century.
BIOLOGY AND THE POSSIBILITY OF A RENAISSANCE OF PSYCHOANALYTIC THOUGHT
It would be unfortunate, even tragic, if the rich insights that have come from psychoanalysis were to be lost in the rapprochement between psychiatry and the biological sciences. With the perspective of time, we can readily see what has hindered the full intellectual development of psychoanalysis during the last century. To begin with, psychoanalysis has lacked any semblance of a scientific foundation. Even more, it has lacked a scientific tradition, a questioning tradition based not only on imaginative insights but on creative and critical experiments designed to explore, support, or, as is often the case, falsify those insights. Many of the insights from psychoanalysis are derived from clinical studies of individual cases. Insights from individual cases can be powerful, as we have learned from Paul Broca's study of the patient Leborgne (
30). The analysis of this patient is a historical landmark; it marks the origin of neuropsychology. Study of this one patient led to the discovery that the expression of language resides in the left hemisphere and specifically in the frontal cortex of that hemisphere. But as Broca's cases illustrate, clinical insights, especially those based on individual cases, need to be supported by independent and objective methods. Broca achieved this by studying Leborgne's brain at autopsy and by subsequently discovering eight other patients with similar lesions and similar symptoms. It is, I believe, the lack of a scientific culture more than anything else that led to the insularity and anti-intellectualism which characterized psychoanalysis in the last 50 years and which in turn influenced the training of psychiatrists in the period of World War II, the period in which psychoanalysis was the dominant mode of thought in American psychiatry.
But the sins of the fathers (and mothers) need not be passed on to succeeding generations. Other disciplines have recovered from similar periods of decline. American psychology, for example, went through a period of insularity and myopia in the 1950s and 1960s despite its being a rigorous and experimental discipline. Under the leadership of Hull, Spence, and Skinner, the behaviorist tradition they espoused focused only on the reflexive and observable aspects of behavior and dealt with these as if they represented all there is to mental life.
With the emergence of computers to model and test ideas about mind, and with the development of more controlled ways of examining human mental processes, psychology reemerged in the 1970s in its modern form as a cognitive psychology that has explored language, perception, memory, motivation, and skilled movements in ways that have proven stimulating, insightful, and rigorous. Modern psychology is still evolving. The recent merger of cognitive psychology with neural science—the discipline we now call cognitive neural science—is proving to be one of the most exciting areas in all of biology. What is the aspiration of psychoanalysis if not to be the most cognitive of neural sciences? The future of psychoanalysis, if it is to have a future, is in the context of an empirical psychology, abetted by imaging techniques, neuroanatomical methods, and human genetics. Embedded in the sciences of human cognition, the ideas of psychoanalysis can be tested, and it is here that these ideas can have their greatest impact.
The following is but one example from my own field, the cognitive neural science of memory. One of the great insights of modern cognitive neural science in the study of memory is the realization that memory is not a unitary function of mind but has at least two forms, called explicit and implicit: a memory for what things are as compared to a memory for how to do something. Explicit memory encodes conscious information about autobiographical events and factual knowledge. It is a memory about people, places, facts, and objects, and it requires for its expression the hippocampus and the medial temporal lobe. Implicit memory involves for its recall an unconscious memory for motor and perceptual strategies. It depends on the specific sensory and motor systems as well as on the cerebellum and the basal ganglia.
Patients with lesions of the medial temporal lobe—or the hippocampus, which lies deep in it—cannot acquire new explicit memories for people, places, and objects. But they fully are able to learn motor skills and are also able to improve their performance on perceptual tasks. Implicit memory is not limited to simple tasks. It also includes a sophisticated form of memory called priming, in which recognition of words or objects is facilitated by prior exposure to the words or visual clues. Thus, a subject can recall the cued item better than other items for which no cues have been provided. Similarly, when shown the first few letters of previously studied words, a subject with temporal lobe lesions often responds by selecting correctly the previously presented word, even though he cannot remember ever seeing the word before!
The tasks that patients who lack explicit memory are capable of learning have in common that they do not require conscious awareness. The patient need not deliberately remember anything. Thus, when given a highly complex mechanical puzzle to solve, the patient may learn it as quickly as a normal person, but on questioning will not remember seeing the puzzle or having worked on it previously. When asked why his performance on a task is much better after several days of practice than on the first day, the patient may respond, “What are you talking about? I've never done this task before.”
What a momentous discovery! Here we have, for the first time, the neural basis for a set of unconscious mental processes. Yet this unconscious bears no resemblance to Freud's unconscious. It is not related to instinctual strivings or to sexual conflicts, and the information never enters consciousness. These sets of findings provide the first challenge to a psychoanalytically oriented neural science. Where, if it exists at all, is the other unconscious? What are its neurobiological properties? How do unconscious strivings become transformed to enter awareness as a result of analytic therapy?
There are other challenges, of course. But at the very least, a biologically based psychoanalysis would redefine the usefulness of psychoanalysis as an effective perspective on certain specific disorders. At its best, psychoanalysis could live up to its initial promise and help revolutionize our understanding of mind and brain.