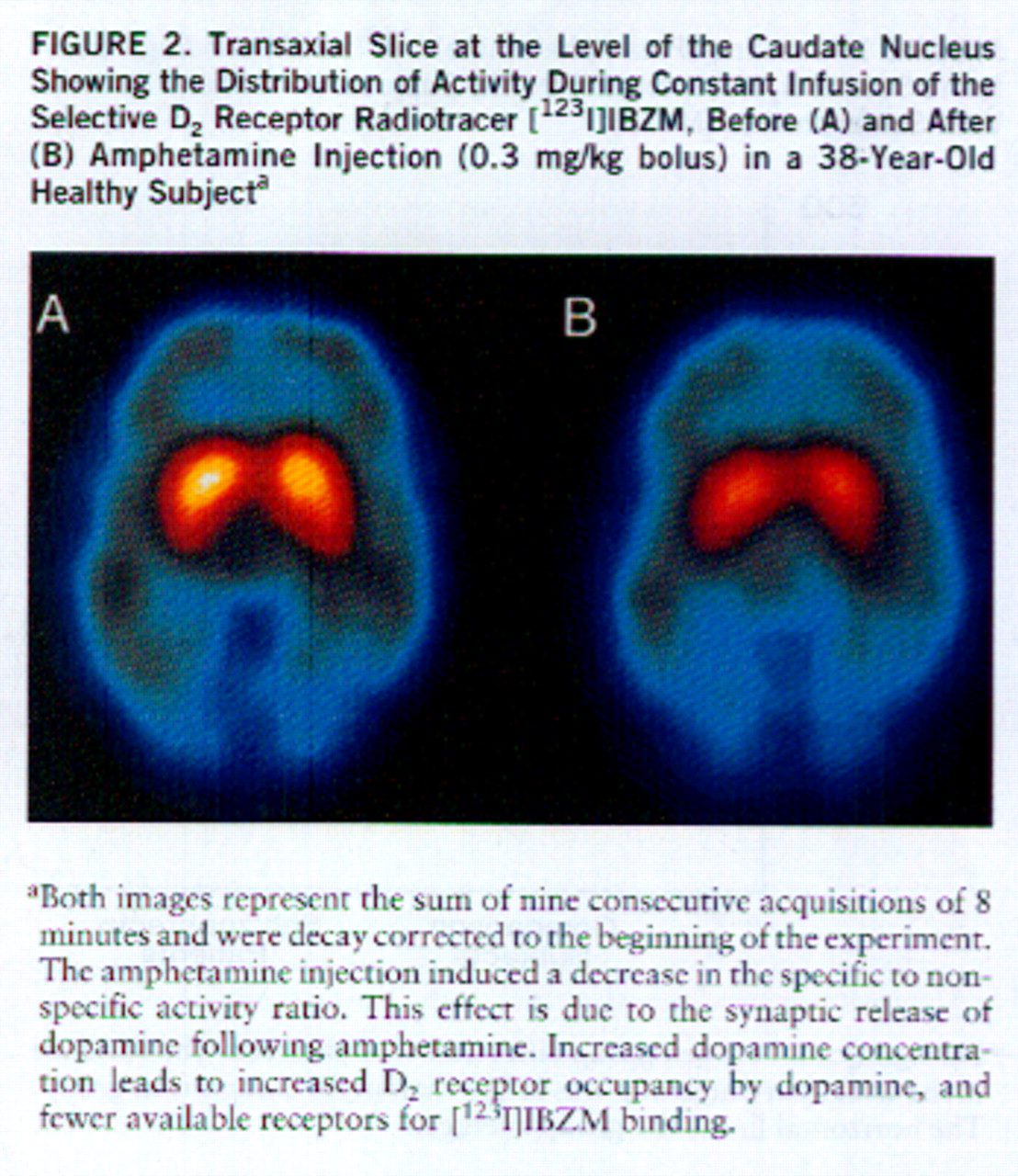
Specifically, we developed a method to measure amphetamine-induced dopamine release with SPECT and the radiotracer [
123I](
S)-(-)-3-iodo-2-hydroxy-6-methoxy-N-[(1-ethyl-2-pyrrolidinyl)methyl]benzamide ([
123I]IBZM), an antagonist at the D
2 and D
5 receptors (
3). We initially observed that amphetamine challenge reduced the [
123I]~IBZM binding potential in baboons. In vivo binding potential is the product of the density and affinity of available receptors, i.e., receptors not occupied by dopamine. Since amphetamine is devoid of significant affinity for D
2 receptors, we postulated that this effect was mediated by increased dopamine release and displacement of [
123I]~IBZM specific binding by dopamine. This mechanism was supported by the observation that pretreatment with the dopamine depleter alpha-methyl-para-tyrosine prevented the effect of amphetamine on [
123I]IBZM binding potential (
4). In addition, we established the existence of a good correlation between amphetamine-induced dopamine release measured with microdialysis and the amphetamine-induced decrease in [
123I]IBZM binding potential measured with SPECT (
4). Therefore, measuring the reduction in [
123I]IBZM binding potential following amphetamine administration provides a noninvasive method to estimate the magnitude of amphetamine-induced dopamine release. Extending this method to healthy volunteers demonstrated its feasibility in humans (
5).
We used this technique to study amphetamine-induced dopamine release in patients with schizophrenia. In a first cohort of 15 patients and 15 matched healthy subjects, we observed a significantly greater amphetamine-induced decrease in the [
123I]IBZM binding potential in the schizophrenic group (mean=–19.5%, SD=15.7%) than in the healthy group (mean=–7.6%, SD=8.0%) (two-tailed t test, F=6.87, df=1,28, p<0.05) (
6). These data provided the first in vivo evidence for a dysregulation of striatal dopamine release in schizophrenia. This result was independently replicated by Breier et al. (
7) using PET, the radiotracer [
11C]raclopride, and a smaller dose of amphetamine (0.2 mg/kg).
In the present investigation, we attempted to replicate these findings in a new cohort of 15 drug-free schizophrenic subjects and 15 matched healthy subjects. We kept the experimental conditions similar to those of the first study in order to allow subsequent meta-analysis of the pooled data.
RESULTS
Subjects
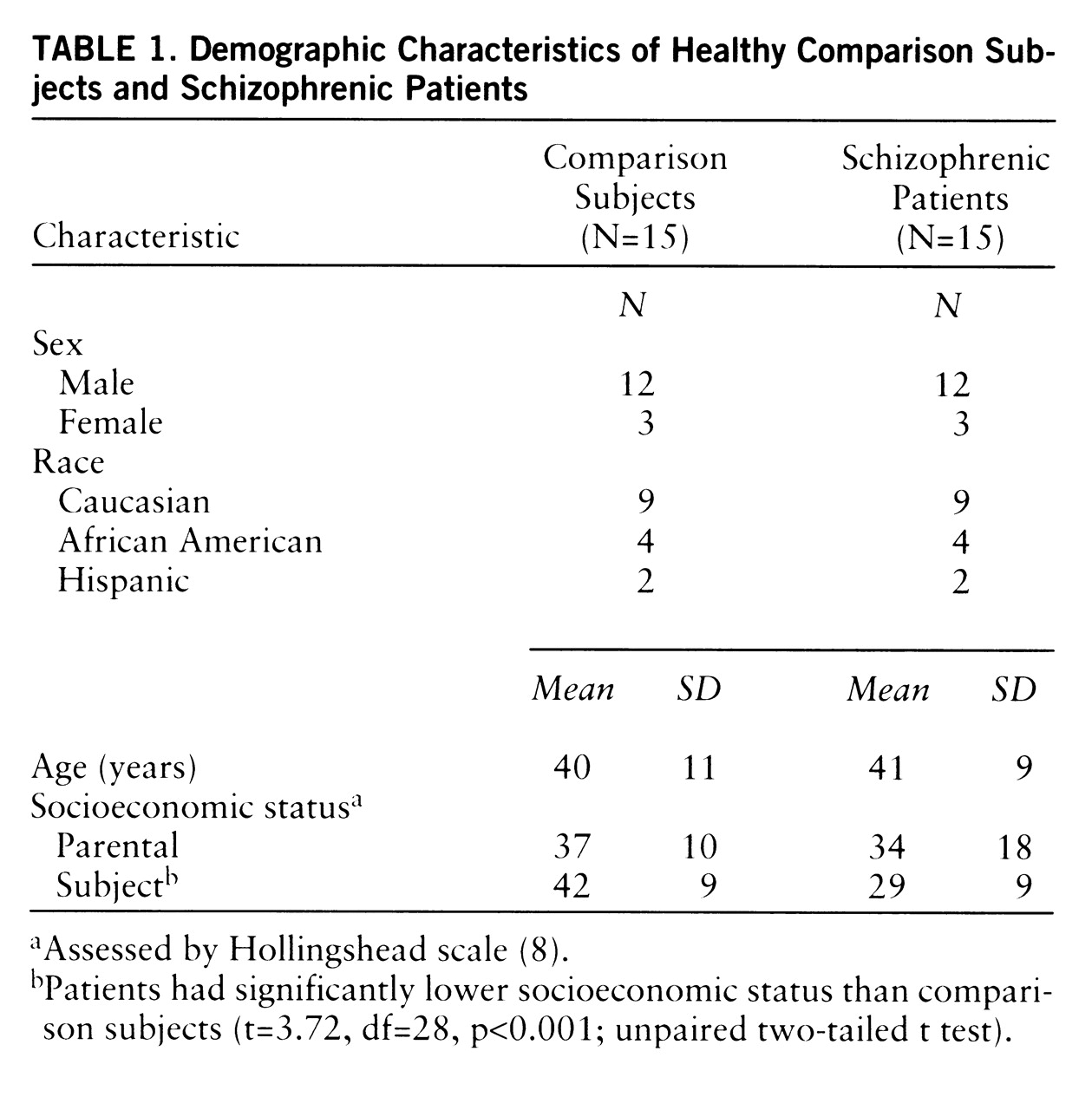
The groups did not significantly differ in age, gender, ethnicity, or parental socioeconomic status (
table 1). Patients had significantly lower socioeconomic status than subjects in the comparison group (F=13.8, df=1,28, p<0.001). Two patients were experiencing a first psychotic episode and were considered neuroleptic naive (one had never received a neuroleptic; the other had less than 1 week of lifetime exposure). The other 13 patients had a mean duration of illness of 17 years (SD=7). Antipsychotics were discontinued in three patients for the purpose of participation in this study (for 22–38 days). Ten patients were noncompliant with antipsychotic treatment at the time of recruitment. For these patients, the average neuroleptic-free interval was 169 days (SD=145) (an index of 365 days was used for patients drug free for more than 1 year [N=3]). After completion of the study, all patients were offered antipsychotic medication and clinical follow-up.
Five patients received lorazepam at doses lower than 3 mg/day up to 24 hours before the scan. At the time of the scan, baseline scores were 44 (SD=11) on the Brief Psychiatric Rating Scale (BPRS) (
12) and 18.5 (SD=5.1) and 19.6 (SD=7.0), respectively, on the positive and negative symptom subscales of the Positive and Negative Syndrome Scale.
Scan Parameters
As previously reported, the bolus plus constant infusion paradigm used in this study, with a bolus to hourly infusion ratio of 3.92 hours, induced steady-state [123I]IBZM plasma levels by the time of the scan. The steady-state quality of the plasma input function was evaluated by the slope of the metabolite-corrected plasma [123I]IBZM concentration from 180 to 260 minutes. This slope was small and not different between groups (schizophrenic patients: mean=–1.7%/hour, SD=7.3%; healthy subjects: mean=–2.3%/hour, SD=3.5%). Similarly, the slope of the occipital activity was negligible and did not differ between the groups (schizophrenic patients: mean=–0.5%/hour, SD=3.6%; healthy subjects: mean=–0.6%/hour, SD=2.9%). None of these slope distributions had a mean value significantly different from zero (one-sample t test), indicating that an adequate steady-state input function was achieved in both groups. Plasma [123I]IBZM clearance or free fraction did not differ between the groups (plasma [123I]IBZM clearance: schizophrenic patients, mean=63 liters/hour, SD=21; healthy subjects, mean=71 liters/hour, SD=10; plasma [123I]IBZM free fraction: schizophrenic patients, mean=3.2%, SD=0.7%; healthy subjects, mean=3.6%, SD=0.9%).
Baseline [123I]IBZM Binding Potential
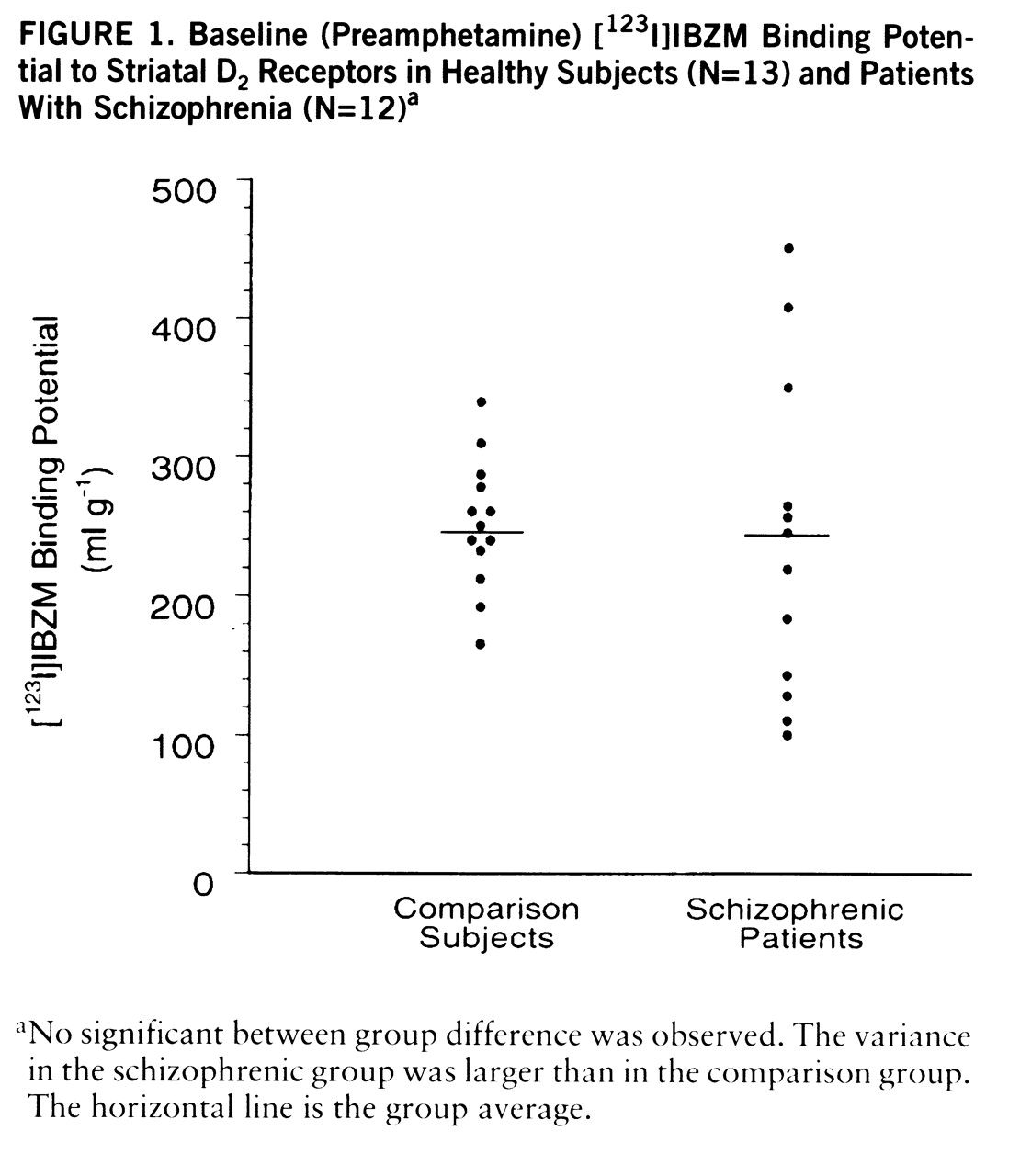
No differences were observed in baseline [
123I]IBZM binding potential between patients (mean=239 ml g
–1, SD=115) and healthy subjects (235 ml g
–1, SD=46) (
figure 1). The variability of [
123I]IBZM binding potential was significantly larger in patients (F=6.10, df=11,12, p<0.005). Baseline [
123I]IBZM binding potential was significantly correlated with age (r
2=0.18, F=5.2, df=1,23, p<0.05), showing a mean decline of 14% (SD=6%) per decade. The interaction between age and diagnosis was not significant, showing no differences in the rate of [
123I]IBZM binding potential decrease with age between schizophrenic and healthy subjects. In the schizophrenic group, baseline [
123I]IBZM binding potential was not correlated with baseline clinical symptoms (total BPRS and positive and negative subscales of the Positive and Negative Syndrome Scale), the neuroleptic-free interval, or lifetime neuroleptic exposure.
Amphetamine Peripheral Effects
Amphetamine plasma levels were similar between groups (schizophrenic patients: mean=23 ng/ml, SD=7; healthy subjects: mean=23 ng/ml, SD=10). Amphetamine induced a marked but transient increase in systolic and diastolic blood pressure. No between-group differences were observed in the blood pressure response to amphetamine (peak increase in systolic blood pressure: schizophrenic patients, mean=36 mm Hg, SD=12; healthy subjects, mean=41 mm Hg, SD=14; peak increase in diastolic blood pressure: schizophrenic patients, mean=20 mm Hg, SD=24; healthy subjects, mean=18 mm Hg, SD=6). Plasma HVA levels at baseline were not different between groups (schizophrenic patients: mean=9.7 ng/ml, SD=2.7; healthy subjects: mean=9.7 ng/ml, SD=3.9). Mean postamphetamine plasma HVA levels were 3.6 ng/ml (SD=1.1) and 3.2 ng/ml (SD=0.6) in healthy subjects and schizophrenic patients, respectively. No differences between groups were observed in the reduction of plasma HVA induced by amphetamine (schizophrenic patients: mean=–60%, SD=10%; healthy subjects: mean=–63%, SD=13%).
Amphetamine Effect on [123I]IBZM Binding Potential
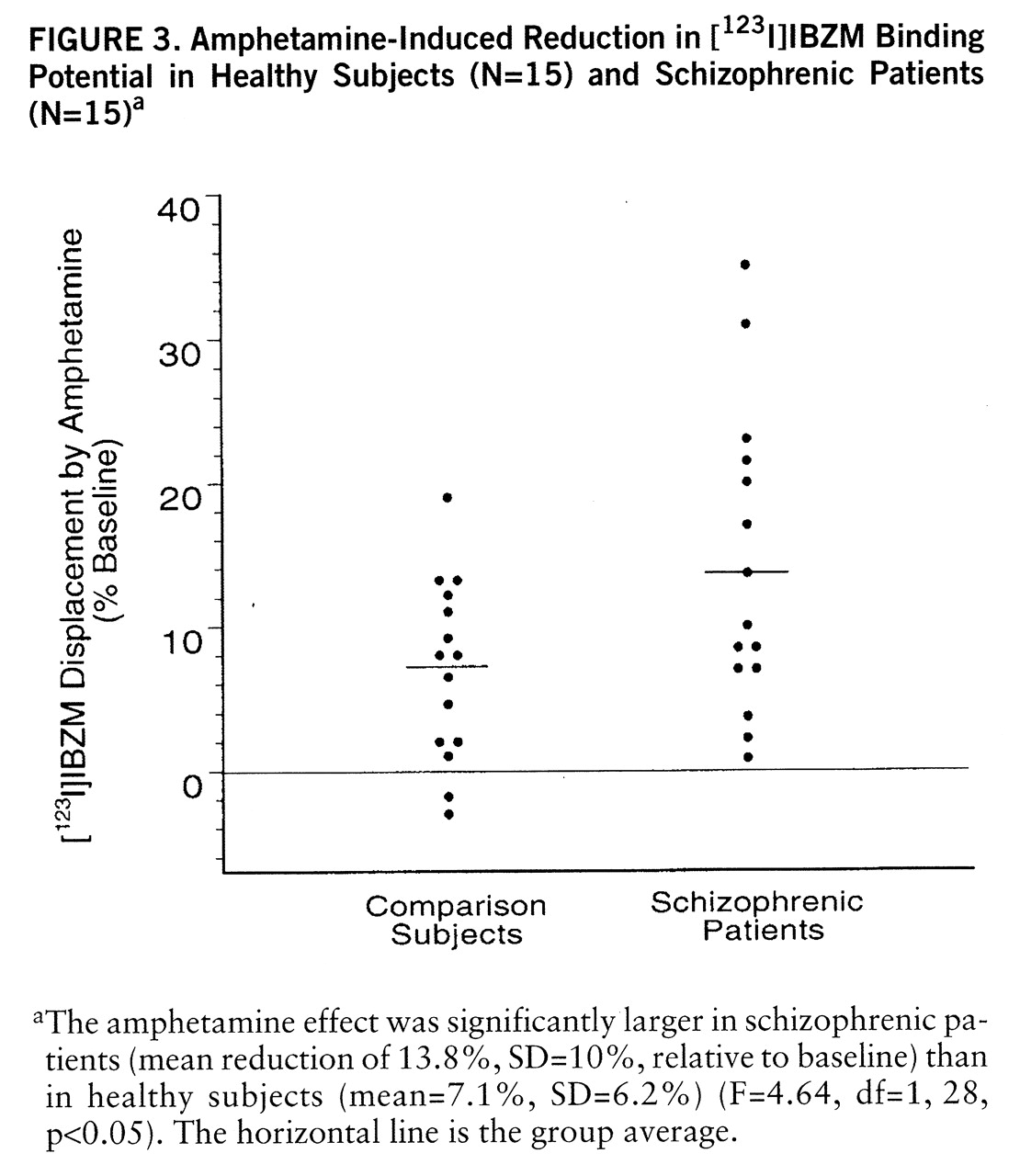
Amphetamine-induced reduction in [
123I]IBZM binding potential was larger in schizophrenic patients (mean=–13.8%, SD=10.3%) than in healthy subjects (mean=–7.1%, SD=6.2%) (F=4.64, df=1,28, p<0.05) (figures
2 and
3). A nonsignificant higher variability in the response was observed in the schizophrenic group (F=2.71, df=14,14, p=0.07). No relationship between age and amphetamine-induced [
123I]~IBZM displacement was observed in the two groups (patients and healthy subjects) combined (r
2<0.01, N=30, p=0.71) or analyzed separately (schizophrenic patients: r
2<0.01, N=15, p=0.91; healthy subjects: r
2=0.02, N=15, p=0.58). No relationship was observed between amphetamine plasma levels and [
123I]IBZM displacement, either in the entire study group (r
2<0.01, N=30, p=0.98) or in each group considered separately (schizophrenic patients: r
2<0.01, N=15, p=0.68; healthy subjects: r
2=0.03, N=15, p=0.58). No correlations were observed between amphetamine-induced [
123I]IBZM displacement and other indices of peripheral amphetamine activity, such as changes in systolic blood pressure (r
2=0.03, N=30, p=0.33) or changes in plasma HVA (r
2=0.05, N=30, p=0.28).
In the schizophrenic group, amphetamine-induced [123I]IBZM displacement was not associated with duration of the neuroleptic-free interval (r2=0.03, N=13, p=0.50), lifetime neuroleptic exposure (r2<0.01, N=13, p=0.88), duration of illness (r2=0.03, N=15, p=0.54), or the use of lorazepam in the week preceding the study (patients taking lorazepam [N=5]: mean=–20%, SD=12%; patients without lorazepam [N=10]: mean=–11%, SD=1%) (F=3.10, df=1,13, p=0.10). The two neuroleptic-naive patients had an amphetamine-induced [123I]IBZM binding potential decrease of 7% and 35%, respectively. The baseline [123I]IBZM binding potential was not associated with the amphetamine-induced [123I]IBZM reduction, either in the groups analyzed together (r2=0.04, N=30, p=0.31) or separately (schizophrenic patients: r2=0.06, N=15, p=0.40; healthy subjects: r2<0.01, N=15, p=0.85).
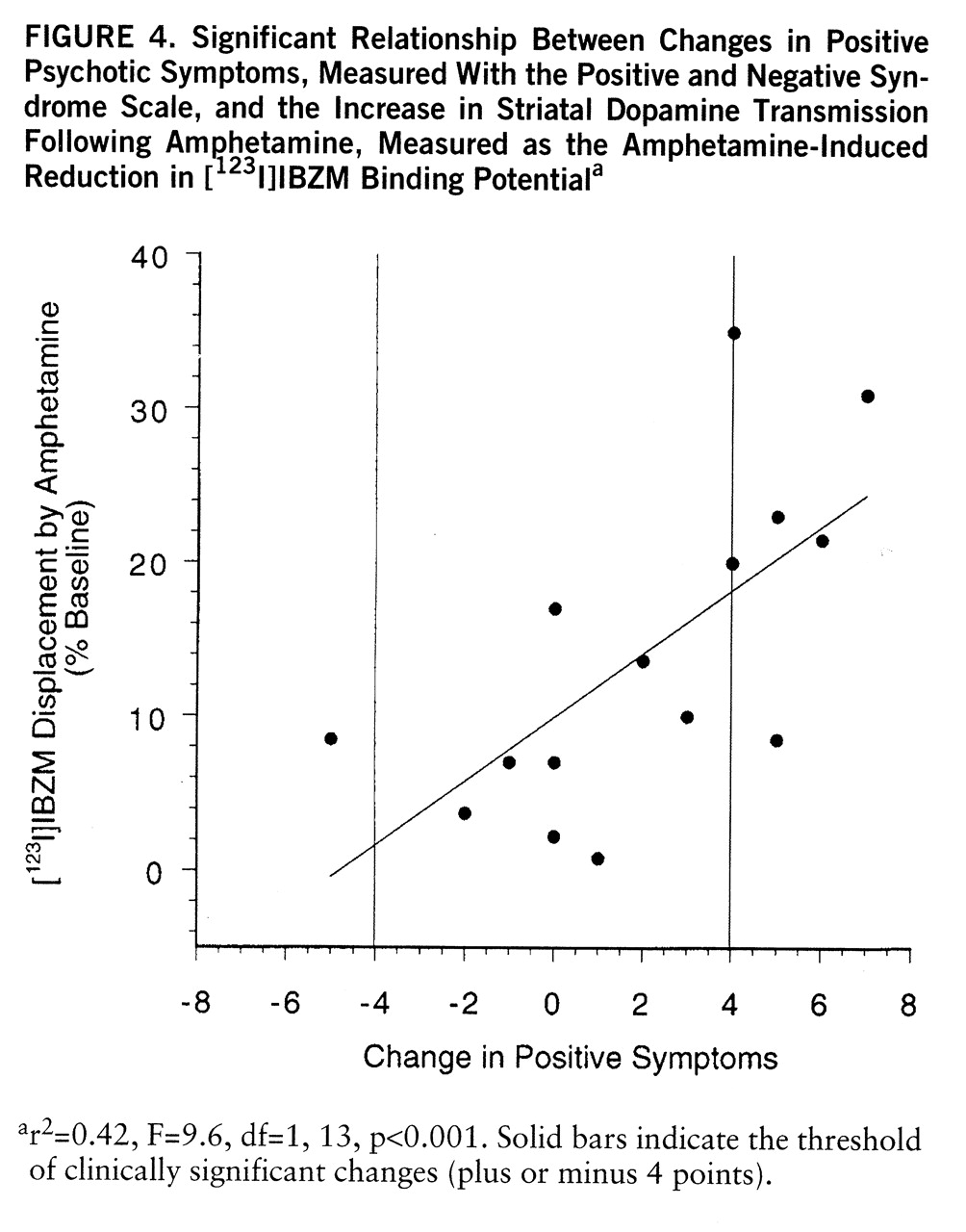
Relationship Between Clinical and Biochemical Response
No psychotic reaction was observed in the comparison group (healthy subjects). In the schizophrenic group, six patients experienced a psychotic reaction to the amphetamine injection (i.e., increase of 4 or more points on the positive subscale of the Positive and Negative Syndrome Scale), eight patients showed no significant change in positive symptoms, and one patient significantly improved (i.e., decrease of 4 or more points on the positive subscale of the Positive and Negative Syndrome Scale). Amphetamine-induced dopamine release was significantly larger in the patients who had a psychotic reaction (mean=–23%, SD=9%, N=6) than in the patients who did not have a psychotic reaction (mean=–8%, SD=5%, N=9) (F=16.6, df=1,13, p<0.005). The increase in positive symptoms was significantly correlated with the reduction in [
123I]IBZM binding potential (r
2=0.42, F=9.6, df=1,13, p<0.001) (
figure 4). Baseline [
123I]IBZM binding potential was not different between schizophrenic patients who had a psychotic reaction (mean=216 ml g
–1, SD=95, N=6) and those who did not (mean=255 ml g
–1, SD=132, N=9). The severity of positive symptoms at baseline was not associated with the magnitude of the amphetamine effect on positive symptoms (r
2=0.03, N=15, p=0.91) or [
123I]IBZM displacement (r
2=0.07, N=15, p=0.32).
Eleven patients showed no significant changes in negative symptoms following the amphetamine challenge, and four patients showed a clinically significant improvement (i.e., decrease of 4 or more points on the negative subscale of the Positive and Negative Syndrome Scale). No significant relationship was observed between improvement in negative symptoms and [123I]IBZM displacement (r2=0.07, N=15, p=0.31). Amphetamine-induced changes in positive and negative symptoms were not correlated (r2<0.01, N=15, p=0.76).
Comparison With First Study
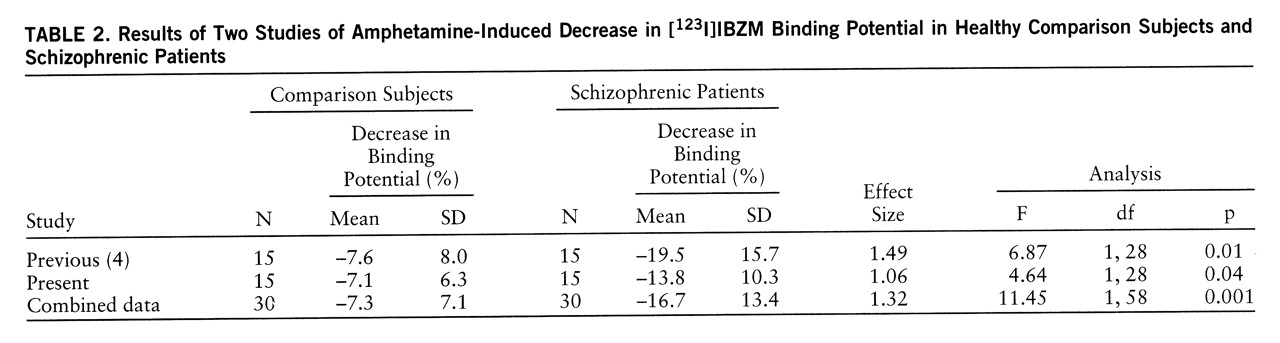
Results in this second cohort of subjects are comparable to the results obtained in the first cohort (
6) (
table 2). Both cohorts were relatively similar in terms of clinical and demographic composition. In each study, no between-group differences were observed in baseline [
123I]IBZM binding potential. In comparison subjects, the amphetamine effects on [
123I]IBZM binding potential were identical in studies 1 and 2. In both studies, the amphetamine effect on [
123I]IBZM binding potential was larger in schizophrenic patients than in healthy subjects. However, in this study, the effect size was lower (1.06) than in the first study (1.49), and the significance level of the group difference was lower (p=0.04 for this study and p=0.01 for the first study). Since the study protocols were identical, results of both studies can be combined. When the results were combined, the effect size was 1.32 and the p value was 0.001 (F=11.4, df=1,58, p=0.001).
DISCUSSION
In a new cohort of 15 schizophrenic subjects and 15 matched healthy subjects, we replicated the observation of a larger decrease in [
123I]IBZM binding potential following amphetamine challenge in schizophrenic patients than in healthy subjects. Findings from these two studies, together with the independent replication by Breier et al. (
7) and the preliminary results recently presented by a third group (
13), clearly indicate abnormal reactivity of dopaminergic transmission to amphetamine exposure in the brain of schizophrenic subjects. We also replicated the observation that this excessive reaction is associated with behavioral consequences, i.e., worsening or emergence of psychotic symptoms.
As in the first study, the increased stimulation of dopamine transmission observed in the striata of schizophrenic patients could not be attributed to differences in plasma amphetamine levels. The measurement of plasma HVA before and after amphetamine was a new feature of this study. Since HVA is formed mainly inside the cell, the decrease in plasma HVA following amphetamine is generally attributed to depletion of the dopamine cytoplasmic pool (
14–
16). Thus, the decrease in plasma HVA is an indirect measure of the intracellular effect of amphetamine in the peripheral nervous system. The similarity in plasma HVA response to amphetamine indicates that the intracellular bioavailability of amphetamine was comparable between the groups. Thus, the observed group differences in striatal dopamine response to amphetamine exposure are related to central nervous system factors.
As in the first study, we could not attribute the increased dopamine response in schizophrenic patients to prior neuroleptic exposure, because of the lack of association between the amphetamine effect and previous neuroleptic exposure or duration of neuroleptic interval. These results indicate that the effect is unlikely to represent a long-term effect of previous neuroleptic medication. Similarly, we were unable to link the effect with lorazepam exposure during the week preceding the study. In conclusion, the effect seems to be associated with the illness process per se.
Abnormal dopaminergic transmission has been suspected in schizophrenia for a long time. These studies represent the first direct demonstration of abnormal regulation of striatal dopaminergic transmission in schizophrenia. The increase in dopaminergic transmission following amphetamine might be due to an increased affinity of D
2 receptors for dopamine (for example, due to a higher proportion of high versus low affinity sites), a larger or more prolonged dopamine release (i.e., presynaptic factors), or a combination of both. Additional studies are needed to better characterize the exact mechanisms responsible for this increased transmission (
4,
6).
Several limitations of this study should be discussed. This method has a relatively low sensitivity. We previously examined in primates the relationship between increase in extrastriatal dopamine, measured with microdialysis, and decrease in [
123I]IBZM binding potential, measured with SPECT. An increase of 45% above baseline of striatal dopamine is needed to decrease [
123I]IBZM binding potential by 1% (
4). A similar ratio (44:1) has been reported by Breier et al. (
7) for [
11C]~raclopride. In a previous study, we showed that in the absence of amphetamine, the reproducibility of [
123I]~IBZM binding potential measurement had an average variability (test minus retest divided by the average) of plus or minus 3.3% (SD=1.7%, range=5% to –5%). The low signal to noise ratio of the method explains why two comparison subjects had an apparent increase in [
123I]IBZM binding potential following amphetamine. On the other hand, the low sensitivity of the method implies that a very large difference in synaptic dopamine increase between patients and comparison subjects corresponded to the measured differences in [
123I]IBZM binding potential decrease.
Another important limitation of the method is that it does not provide any information about the baseline dopamine concentration, i.e., concentration in the absence of amphetamine. In this study we measured only the relative increase in dopamine concentration resulting from the amphetamine challenge. The observation of a normal D
2 receptor availability at baseline does not imply that baseline dopamine levels are unaltered in schizophrenia (
17). A study based on rapid induction of dopamine depletion with alpha-methyl-para-tyrosine is currently being conducted in our group to address this issue (
18).
Finally, the low number of patients not previously exposed to antipsychotics makes it difficult to rule out the possibility that the observed effect might represent a long-term consequence of prolonged neuroleptic exposure. More neuroleptic-naive patients should be studied to address this important question.
The effect size in the second cohort (1.06) was lower than that in the first cohort (1.49). The reasons for the lower effect size are unclear. The scan protocol was identical, comparison subjects had similar effects (
table 2), and the clinical characteristics of the patients were similar (tables 1 in this article and in reference 6). Therefore, it is likely that the difference in effect size is due to a sampling effect, possibly related to the large variance of the amphetamine effect in the schizophrenic group.
The mechanism, specificity, significance, and etiology of this increased dopaminergic response in schizophrenia remains to be elucidated. However, recent preclinical studies do offer interesting hypotheses. Neonatal hippocampal lesions in rodents and primates provide a potential neurodevelopmental animal model for some clinical and biochemical features of schizophrenia (
19). Regulations of striatal dopamine release were compared with microdialysis between adult rhesus monkeys who underwent ablation of the amygdala-hippocampal formation within 3 weeks of birth, comparison monkeys, and monkeys who underwent the same lesion ablation during adulthood (
20). Local perfusion of amphetamine in the prefrontal cortex induced a decrease in striatal dopamine release in comparison monkeys and in monkeys with adult lesions. This observation is consistent with the inhibition of subcortical dopamine by prefrontal dopamine (
21). In contrast, in monkeys with neonatal lesions, prefrontal amphetamine perfusion induced an increase in striatal dopamine release, i.e., a reversal of the normal effect (inhibition). Thus, a neonatal hippocampal lesion does affect prefrontal inhibition of striatal dopamine release. The relevance of this model to schizophrenia remains to be established. Brain imaging techniques now provide the tools to directly test these preclinical models in patients.
In conclusion, we replicated in this second cohort our previous observation that schizophrenia is associated with an increased dopamine transmission following acute amphetamine challenge (
6). The results of both studies were very similar, with the exception that the effect size in the second study was less pronounced than that in the first study. These studies, as well as similar results provided by other groups (
7,
13), are consistent in documenting this excessive neurochemical response and provide direct evidence to support the hypothesis of a dysregulation of central dopamine transmission in schizophrenia.