More recently, gender differences in resting CBF and cerebral glucose metabolism have been examined. Higher resting cortical and global values in women than men have been noted in some studies
(11,
12). Regionally, higher metabolic rates have been observed in women in the cerebellum and in the cingulate gyrus, with opposite findings in some neocortical and limbic regions
(13,
14). These data have been interpreted as reflecting the neurobiological substrates underlying sex differences in cognitive and emotional processing. Subsequently, both age and sex influences on brain neuroanatomy and metabolic function have been reported
(15). With advancing age, men have greater declines in cerebral hemisphere volumes and frontal and temporal lobe volumes, while women show greater atrophy in parietal and hippocampal regions.
In the present study we examined regional differences in mu-opioid receptor binding measures in the brains of healthy human subjects, with the mu-selective agonist [11C]carfentanil and PET. The data described represent two separate analyses and subject groups. First, we retrospectively examined data from prior research protocols. In addition, a second study group was prospectively recruited to control for prior hormonal treatments and phase of the menstrual cycle.
RESULTS
The scan data obtained from the retrospective analysis and the prospectively recruited group were significantly different. Greater differences between regions and higher levels of receptor binding were shown by the data obtained with the newer-generation scanner (see Method, Prospective Analysis), reflecting improved spatial resolution and z axis sampling (data not shown: all regions showed differences between the data acquired with the two scanner types; unpaired, two-tailed t tests, df=64, p<0.05). Therefore, these groups were not combined for any of the analyses presented.
There were no significant differences in age between the male and female subjects in either group (unpaired, two-tailed t tests; retrospective data: t=0.1, df=34, p>0.05; prospective data: t=0.4, df=28, p>0.05). For the regions sampled in both hemispheres (neocortical regions, basal ganglia, and amygdala), no significant right-left asymmetries were observed in the men or women (paired, two-tailed t tests; retrospective group: df=34, p>0.05 for all regions examined; prospective group: df=28, p>0.05 for all regions); therefore, the right and left regions were averaged for subsequent analyses.
In addition, no age or sex effects were observed in activity in the occipital cortex, a reference region used for the estimation of nonspecific and free tracer activity (see Method) (one-way ANCOVA; retrospective group: df=1, 33, p>0.05; prospective group: df=1, 27, p>0.05). Activity in this region is a constant within each subject and reflects factors such as amount of radiotracer administered, metabolism and clearance of the parent compound, and tracer transport across the blood-brain barrier (see reference
21 and Method section).
Retrospective Analysis
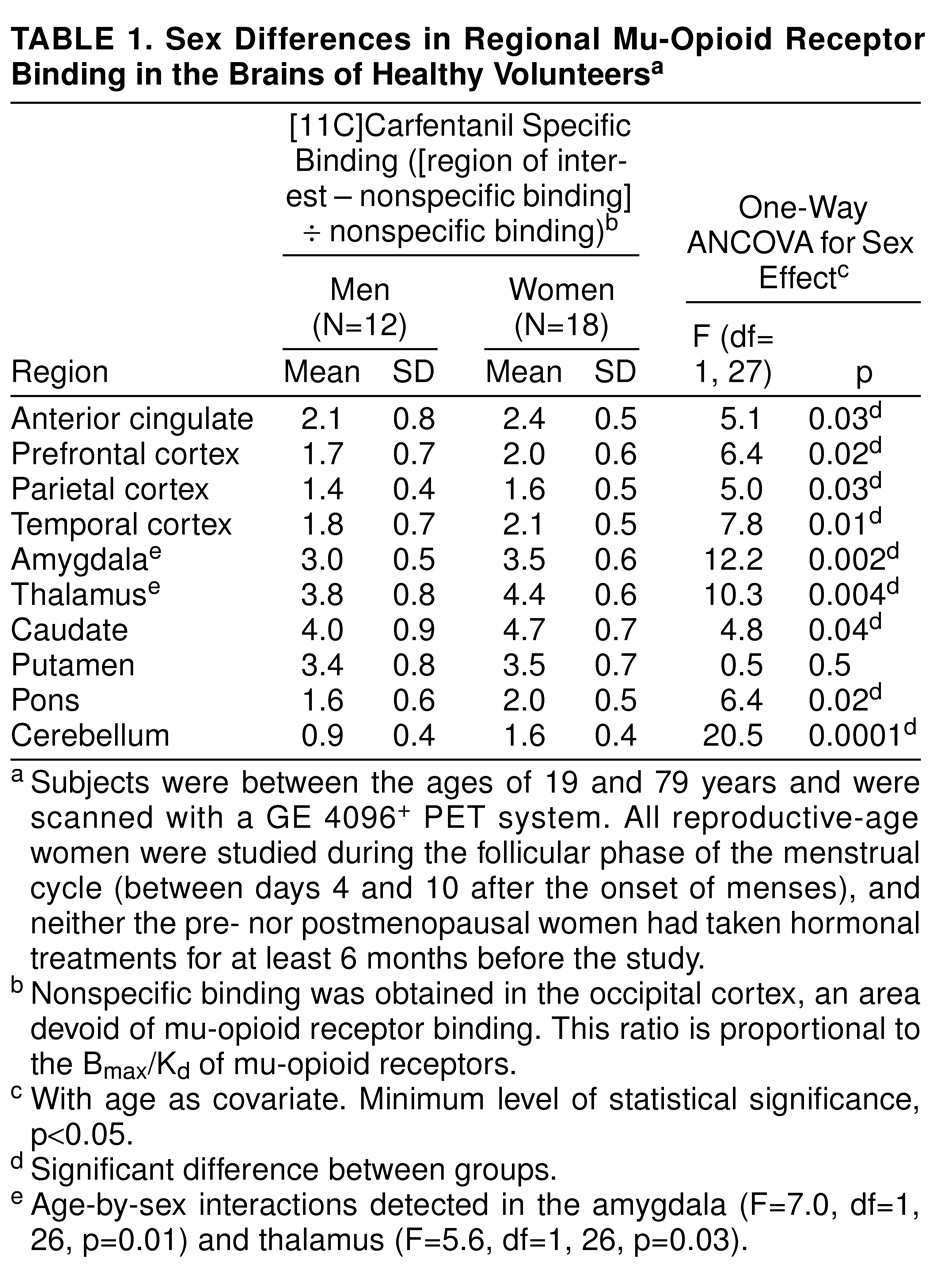
Separate ANCOVAs for each region were calculated, with sex as a grouping factor, region as a dependent variable, and age as a covariate. The ANCOVAs did not show significant age-by-gender interactions for any brain region. Accordingly, the interaction term was eliminated from the model for subsequent analyses. Age effects were detected in the prefrontal (F=11.4, df=1, 33, p<0.001), temporal (F=6.7, df=1, 33, p<0.01), and parietal (F=7.7, df=1, 33, p<0.01) cortices, amygdala (F=7.3, df=1, 33, p<0.01), thalamus (F=6.4, df=1, 33, p<0.01), and pons (F=10.8, df=1, 33, p<0.01). No significant age effects were observed in the anterior cingulate cortex, caudate, or putamen. Gender effects, with higher mu-receptor availability in the women than in the men, were detected in the temporal cortex (F=5.8, df=1, 33, p=0.02), amygdala (F=9.6, df=1, 33, p<0.01), and pons (F=6.8, df=1, 33, p<0.01) and, to a lesser extent, in the parietal cortex (F=4.2, df=1, 33, p<0.05) and caudate nucleus (F=4.2, df=1, 33, p<0.05). No significant gender differences were observed in the prefrontal or anterior cingulate cortex, putamen, or thalamus.
The direction of age-related changes in mu-opioid receptor binding potential were further analyzed by means of Pearson’s correlations. Negative associations between mu-opioid receptor binding and age were found in some subcortical structures: amygdala (r=–0.38, df=34, p=0.02), thalamus (r=–0.41, df=34, p<0.01), and pons (r=–0.46, df=34, p<0.005). Conversely, mu-receptor binding increased with age in neocortical areas: prefrontal cortex (r=0.49, df=34, p=0.002), temporal cortex (r=0.39, df=34, p=0.02), and parietal cortex (r=0.42, df=34, p<0.01). These corresponded to mean increases in mu-opioid receptor binding of 17%–34%, from the third decade of age (N=15) to the seventh and eighth decades (N=7), in neocortical areas. In the amygdala, thalamus, and pons, binding decreased by 19%, 34%, and 39%, respectively.
Prospective Study
ANCOVA detected significant age and sex effects, as well as sex-by-age interactions, in various brain regions. Age effects were observed in the prefrontal (F=38.7, df=1, 27, p<0.0001), anterior cingulate (F=19.9, df=1, 27, p<0.0001), temporal (F=34.9, df=1, 27, p<0.0001), and parietal (F=53.3, df=1, 27, p<0.0001) cortices and in the putamen (F=14.4, df=1, 27, p<0.001). The coefficients for Pearson correlations between age and regional binding potential reached r values of 0.72, 0.70, 0.70, 0.78, and 0.58 for the preceding respective regions (df=28, p<0.001). The subjects in the third decade of age (N=9) had higher levels than did the subjects older than 60 (N=10); the percentage differences for the preceding regions were 79%, 50%, 62%, 89%, and 36%, respectively.
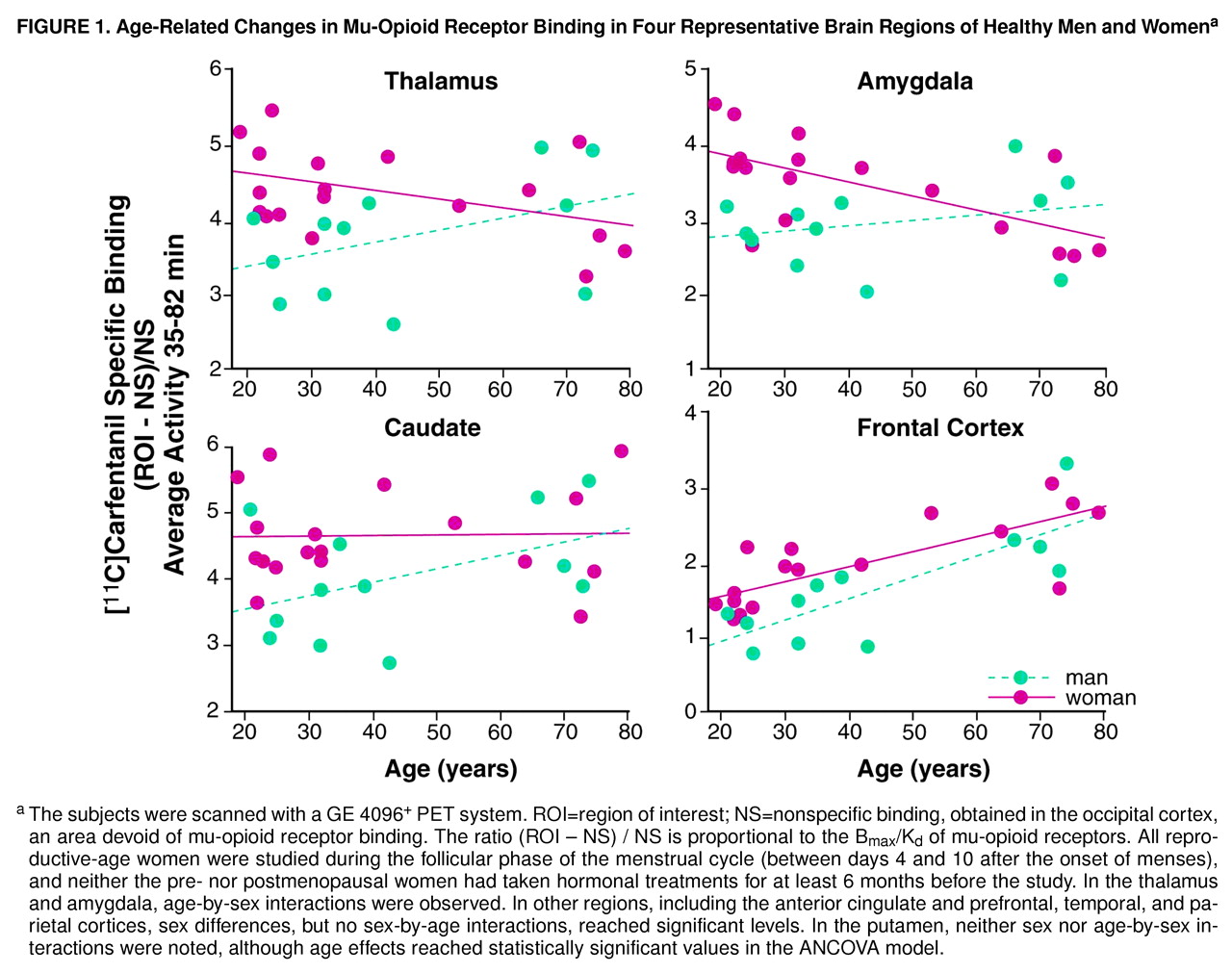
Gender effects were detected in the prefrontal, anterior cingulate, temporal, and parietal cortices, the caudate nucleus, and the cerebellum; higher levels of mu-opioid receptor availability were present in the women (
Table 1 and
Figure 1). No sex differences were noted in the putamen.
Statistically significant age-by-gender interactions were detected in the amygdala and thalamus (
Figure 1 and table 1 footnote). The nature of these interactions was further explored by analyzing age and gender effects separately. In the men (N=12), no significant correlations with age were observed in these regions (Pearson correlations, df=10, p>0.05). In the women (N=18), regional binding was negatively correlated with age (amygdala: r=–0.63, df=16, p<0.01; thalamus: r=–0.42, df=16, p=0.07). Reproductive-age women (N=11, mean age=26 years, SD=5, range=19–32) were also compared with men in the same age range (N=7, mean age=30 years, SD=7, range=21–39), and those women showed higher levels of binding (unpaired, two-tailed t tests). In the amygdala, the mean difference was 29% (t=3.7, df=16, p<0.005), and in the thalamus the mean difference was 23% (t=3.4, df=16, p<0.005). When the postmenopausal women (N=5, mean age=73 years, SD=6, range=64–79) were compared to men in the same age range (N=4, mean age=71 years, SD=4, range=66–74), their binding was found to be below the men’s levels, with mean differences of –24% in the amygdala and –12% in the thalamus. Two-tailed, unpaired t tests did not show statistically significant differences for this age group, likely because of the small number of subjects included.
Even in the regions in which sex effects but no sex-by-age interactions were observed, there was a tendency toward greater female-male differences in mu-opioid receptor binding during the reproductive years than after the menopause. Reproductive-age women had an average of 26% more binding than men in the prefrontal cortex, 24% in the anterior cingulate, 25% in the temporal cortex, 8% in the parietal cortex, 20% in the caudate nuclei, and 62% in the cerebellum. In the postmenopausal women and same-age men, these differences were somewhat less pronounced: 14% higher binding among the women in the prefrontal cortex, 7% in the anterior cingulate, and 13% in the temporal cortex. The women’s receptor availability was 13% below male levels in the caudate nucleus. Binding was 19% and 59% higher in the parietal cortex and cerebellum, respectively.
DISCUSSION
The endogenous opioid system, the effects of which are mediated by three primary receptor subtypes (mu, delta, and kappa), has been implicated in a number of normal functions and may be altered in various pathological states. It appears to modulate responses to stress, learning, and memory acquisition, it is involved in the effects of substances of abuse, it regulates pain mechanisms, and it has been implicated in the pathophysiology of various illnesses, such as seizure disorders, depressive illness, eating disorders, and Alzheimer’s disease.
The data presented here, replicated in two different study groups, demonstrate both age- and sex-associated differences in mu-opioid receptor binding measures in the brains of living healthy humans. The influences of age and sex on this neuroreceptor site were observed in some, but not other, regions.
We observed increases in mu-opioid receptor availability with advancing age in neocortical regions and the putamen, findings that are consistent with data obtained in studies of experimental animals and two human postmortem studies
(16,
17). While our method of analysis (binding potential, B
max/K
d [22]) does not differentiate changes in receptor binding (B
max) from changes in receptor affinity (K
d), the study of Gross-Isseroff et al.
(16) showed that regional age-related changes in mu-opioid receptor binding observed in postmortem subjects (up to 30%–60% of whom were in their third to sixth decades) were related to alterations in B
max and not K
d. The percentage increase described in their work is also comparable to that observed in the present study for subjects in the prospectively recruited group, who were studied with the higher-resolution scanning system.
Women showed higher mu-opioid receptor binding values than men in several brain regions, both cortical and subcortical. With the ANCOVA model, significant sex effects were detected in neocortical areas (prefrontal, parietal, temporal) and in the caudate nuclei, amygdala, thalamus, and cerebellum (but not in the putamen), if no correction for multiple comparisons was applied. If the threshold of significance was reduced to account for multiple tests, the regions that remained significant would be limited to the amygdala, thalamus, and cerebellum. It is interesting that the age-associated changes differed between the sexes in the amygdala and thalamus. While women had higher opioid binding potential than men during the reproductive years, binding decreased below that in men after the menopause. This was reflected in the ANCOVA model as a sex-by-age interaction. A number of other regions also showed smaller male-female differentials after the menopause, although not to the extent observed in the amygdala or thalamus. While the sex differences in mu-opioid binding during the female reproductive years may reflect early, gender-specific ontogenetic factors, the changes observed in the postmenopausal women suggest that the adult hormonal milieu is also capable of modulating this neurochemical system at least in some brain regions.
Some differences between the data acquired with the two scanning systems and patient groups were apparent. More regions were found to show age and gender effects with the newer scanning system, a finding that is likely accounted for by improved spatial resolution and sensitivity. In addition, a larger number of women were studied in the prospectively recruited group, which may have increased our ability to find differences between genders and age-by-gender interactions.
We considered the possibility that technical issues may have influenced the data. Factors that sometimes confound the interpretation of PET receptor measures include changes in blood flow (and therefore blood-brain transport of the radiotracer). For some radiotracers, regional changes in CBF may influence the receptor measure if uptake is “flow dependent”
(24). However, computer simulations in which modifications in radiotracer transport (K
1) were introduced showed that the distribution of [
11C]carfentanil is not subject to significant flow effects. Also, rCBF tends to decrease with advancing age, while mu-opioid receptors increased in most brain regions with advancing age, in agreement with postmortem data
(16,
17).
A second element that causes erroneous measurements is the presence of partial volume effects due to limited resolution and differential degrees of regional atrophy
(25). Smaller brain regional volumes imply less recovery of tracer activity. While correction for atrophic changes was not applied to these data, women and men have not been shown to have such differences in cortical gray volumes
(10,
15), and so this issue does not explain either the sex differences observed during the reproductive years or the decline in binding observed in the postmenopausal women.
The data presented have important theoretical and practical implications. First, they experimentally support the recent policy of including both sexes in human research. They also imply that functions in which the opioid system plays a role, such as antinociceptive or reward mechanisms, may be differentially modulated by gender and age.
In one study
(19) a more profound effect of kappa-opioid agonists was observed in women than in men. While systematic data addressing sex differences in mu-opioid agonist antinociception in humans are not available, our results suggest that these may also be present. In this regard, the amygdala and the thalamus have been implicated in antinociceptive function
(26). The higher mu-opioid receptor binding would predict that sensitivity to mu-opioid analgesics would be greater in reproductive-age women than in men, with the reverse being the case in postmenopausal women. However, in the present study there were too few older subjects to compare them to the younger subjects, so this hypothesis should be reexamined in larger cohorts. Also, the finding of sex-by-age interactions in the amygdala and the thalamus would not survive after correction for multiple comparisons. Further studies will require specific attention to reproductive status to confirm our findings and explore their physiological implications.
Other behavioral and cognitive implications of our findings are still to be explored. The opioid system is thought to be involved in a variety of human functions, including emotional and reward regulation, learning, and memory acquisition. For example, mu-opioid receptors are modulated as a response to stressors
(27,
28). High levels have been found in the frontal cortex of suicide victims
(16,
17) and may be present in patients diagnosed with major depression
(29). High doses of naloxone, a nonselective opioid antagonist, appear to interfere with performance in verbal learning tasks
(30). It is interesting that women usually outperform men in verbal functions but have worse performance after menopause or surgical oophorectomy
(31,
32). Hypoestrogenemia may also worsen the course of Alzheimer’s disease
(33). In this illness, low mu-opioid receptor binding in the amygdala has been described in human postmortem tissue
(34). The influence of gonadal steroids on amygdala mu-opioid binding and its implications remain to be examined.
Much work remains to be done in the study of gender and age effects on neurochemical systems in human subjects. This report points to one neurotransmitter system, the endogenous opioid, for which both factors have significant brain regional influences. Further studies should be targeted to the examination of the behavioral and cognitive implications of these influences in normal processes (e.g., cognition, emotional processing) and in disease states (e.g., substance abuse, painful conditions) in which the opioid system may play a key role.