Working memory is a neuropsychological construct describing a set of cognitive processes involved in maintaining and manipulating information to guide task-appropriate behavior
(1,
2). The prefrontal cortex plays a key role in performance of working memory tasks in animals and in humans
(2–
4). A number of studies have also indicated that there is a physiological range of synaptic dopamine-regulating activity of prefrontal cortical neurons during working memory
(5,
6). Excessive facilitation or inhibition of dopamine signaling results in diminished working memory performance
(5). Functional imaging studies in humans have largely been consistent with these data
(7–
9).
Working memory deficits are cardinal features of schizophrenia
(10), and they are in part genetically determined. Several studies have demonstrated familial aggregation of schizophrenia with deficits in neuropsychological tests sensitive to prefrontal lobe damage, including tests of working memory
(11,
12). Moreover, working memory deficits scale linearly with degree of genetic loading for schizophrenia
(13).
There is overwhelming evidence of dorsolateral prefrontal cortex involvement in working memory deficits in schizophrenia
(14–
25). Moreover, there is converging evidence that reduced dopamine signaling in the dorsolateral prefrontal cortex may account for at least part of this deficit
(26–
31). Catechol
O-methyltransferase (COMT), because it metabolizes released dopamine, plays an important role in modulating the activity of prefrontal circuitry during performance of working memory tasks. Despite the widespread distribution of COMT principally in nondopaminergic neurons
(32), pharmacological studies have indicated that the catabolic flux of synaptic dopamine through the COMT pathway is characteristic of the prefrontal cortex in contrast to the striatum
(33,
34). Of note, studies of COMT knockout mice have demonstrated that only dopamine levels (in contrast to other biogenic amines) are increased and only in the prefrontal cortex
(35). Further, COMT inhibitors have been shown to improve working memory in animals and in humans
(36,
37). This regionally specific effect may be due to the fact that, in contrast to the striatum, dopamine transporters in the prefrontal cortex are expressed in low abundance, not within synapses, and appear to have little if any impact on synaptic dopamine levels
(33,
34). These data strongly support the notion that variation in COMT activity may have neurobiological effects specific to the prefrontal cortex. Recent studies in humans by Egan et al.
(38) and Mattay et al.
(8) have demonstrated a relationship between a common functional polymorphism (Val
108/158Met) in the COMT gene with working memory performance and related dorsolateral prefrontal cortex physiology measured with functional magnetic resonance imaging (fMRI). The Val and Met alleles are codominant in accounting for a significant variation in COMT enzyme activity and dopamine catabolism in peripheral blood
(39) and in human postmortem prefrontal cortex (unpublished data of J. Chen and D.R. Weinberger). Relative to low-activity Met allele carriers, carriers of the high-activity Val allele show inefficient cortical processing as reflected by lower performance along with greater prefrontal cortical blood-oxygen-level-dependent (BOLD) response. Several subsequent studies in different patient populations have confirmed the relationship between COMT and working memory performance
(40,
41). Furthermore, the high-activity Val allele has been shown to be associated with risk for schizophrenia in several family-based association studies
(38,
42).
Since treatment with second-generation antipsychotics enhances working memory in some patients (possibly via increasing prefrontal dopamine
[43]), we hypothesized that COMT genotype might interact with the effect of atypical antipsychotic drugs on the prefrontal cortex in patients with schizophrenia. Since working memory deficits share some variance with negative symptoms and since the latter have been associated with lower levels of prefrontal dopamine, we also investigated the effects of COMT genotype in determining differential improvements in negative symptoms.
Method
Subjects and Treatment
We studied 30 patients (23 men and seven women; mean age=28.6 years [SD=8.8]) suffering from an acute psychotic episode who had been assessed with the Structured Clinical Interview for DSM-IV Axis I Disorders
(44). Fourteen patients had a diagnosis of schizophrenia and were drug-free (mean=5.6 months, SD=5.7) at entry into the study. Sixteen had a diagnosis of schizophreniform disorder and were all drug-naive. These 16 patients were followed longitudinally and confirmed to have a diagnosis of schizophrenia. All patients were treated with olanzapine monotherapy. Titration was allowed for the first 2 weeks, and then the dose was kept constant until 8 weeks of treatment (mean dose=21.1 mg/day, SD=7.6). Exclusion criteria were history of significant drug or alcohol abuse, active drug use in the past year, head trauma with loss of consciousness, and any significant medical condition. While two of the patients had a history of sporadic drug use (cannabis), none of the patients had a history of chronic drug abuse. These two patients underwent urinary screening for major drugs of abuse at admission and the results were negative.
Symptoms were assessed at study entry and days 7, 14, 28 (4 weeks), and 56 (8 weeks) with the Positive and Negative Syndrome Scale. Other demographic information collected included length of illness (mean=62.1 months [SD=76.8]), parental socioeconomic status (Hollingshead scale: mean=29.2 [SD=15.9]), handedness (Edinburgh Inventory: mean=0.75 [SD=0.45]), total IQ (WAIS-R: mean=82.1 [SD=16]), and premorbid IQ (Italian version of Wide Reading Achievement Test—Revised: mean=98.1 [SD=7.6]).
The present study was approved by the local institutional review board. Moreover, after complete description of the study to the subjects, written informed consent was obtained.
Genotype Determination
COMT Val
108/158 Met genotype was determined as a restriction fragment length polymorphism after polymerase chain reaction amplification and digestion with
NlaIII
(38).
Working Memory Paradigm
Working memory was assessed with N-back tasks as in earlier reports
(4). Briefly, “N-back” refers to how far back in the sequence of stimuli that the subject had to recall. The stimuli consisted of numbers (1–4) shown in random sequence and displayed at the points of a diamond-shaped box. There was a non-memory-guided control condition (0-back) that presented the same stimuli but simply required subjects to identify the stimulus currently seen. As memory load increased, the task required the recollection of a stimulus seen one stimulus (1-back) or two stimuli (2-back) beforehand while continuing to encode additionally incoming stimuli. Performance data were recorded as the number of correct responses (accuracy) and reaction time.
fMRI Data Acquisition and Processing
Echo planar imaging BOLD fMRI data were acquired from 20 patients at 4 and 8 weeks as described previously (TE=30 msec, TR=2 seconds, 20 contiguous slices, voxel dimensions=3.75×3.75×5 mm)
(18) on a conventional GE 3-T machine equipped with a standard head coil. Ten patients did not complete both fMRI studies or were excluded for technical reasons. We used a simple block design in which each block consisted of eight alternating 0-back and rest (subjects were instructed to fixate the diamond on the screen) conditions (each lasting 30 seconds). Similar blocks were used for the 1-back or 2-back working memory conditions alternating with the 0-back condition. Each task combination was obtained in 4 minutes and 8 seconds, 120 whole-brain scans. The first four scans at the beginning of each time series were acquired to allow the signal to reach a steady state and were not included in the final analysis. The order of the task combinations was counterbalanced across subjects but maintained within subjects across time.
All fMRI data were reconstructed, registered, linear detrended, globally normalized, and then smoothed (10-mm Gaussian kernel) before analysis within SPM 99
(45). The fMRI data were then interrogated in two ways for high data quality (scan stability) prior to inclusion in any further analysis. First the registration parameters were extracted and used to exclude subjects with excessive interscan motion (>2 voxels translation, >1° rotation)
(18). Second, we used evidence of motor cortex activation as an internal activation standard for both intra- and interscan variability
(18). Since subjects responded using their right thumb, subjects had to demonstrate activation of the contralateral (left) primary motor cortex in comparison with rest (p<0.001). Subjects without such activation were excluded based on the assumption that MRI artifact of some kind remained in the data after reconstruction and registration (N=3 of the 10 patients not providing fMRI data for the study).
Statistical Analysis
The positive symptom, negative symptom, general psychopathology, and total scores of the Positive and Negative Syndrome Scale were entered into separate multivariate analyses of covariance (MANCOVAs) (covarying for the score at baseline and for gender), with genotype as a between-subject factor and time as a within-subject factor. For a qualitative analysis, the Positive and Negative Syndrome Scale scores were entered into separate chi-square analyses; 30% improvement from baseline at 8 weeks was used as a cutoff to determine treatment response from nonresponse. MANCOVA (covarying for gender) with genotype as a between-subject factor and time as well as N-back task (0-back, 1-back, and 2-back) as within-subject factors were used to investigate working memory data.
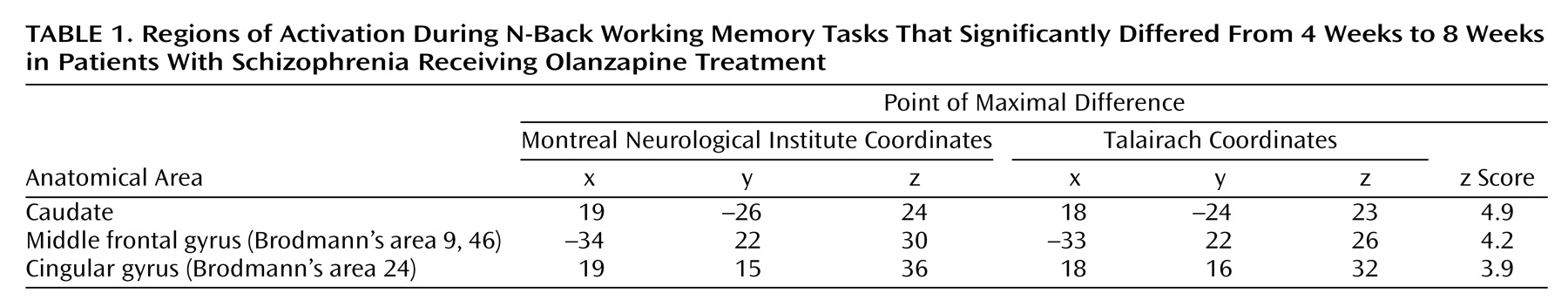
The fMRI data were analyzed as a time series modeled by a sine wave shifted by an estimate of the hemodynamic response. Individual subject maps were created by using one-sample t tests. The resultant contrast images were then entered into second-level (random effects) analyses for the two time points (4 and 8 weeks) and then into analyses of covariance (ANCOVAs) (covarying for performance and gender, p<0.001 uncorrected, cluster size eight voxels). Statistically significant group differences (
Table 1) were reported as voxel-intensity z values. For anatomical localization, statistical maxima of activation were converted to conform to the standard space of Talairach and Tournoux
(46).
Results
Genotype Determination
The genotype results for the patients were as follows: Met/Met: N=5, Val/Met: N=17, and Val/Val: N=8; this distribution was consistent with Hardy-Weinberg expectations (χ2=0.6, df=2, p>0.70). The three genotype subgroups did not differ in any demographic variable other than gender (χ2=7.5, df=2, p<0.02). All subsequent analyses were covaried for gender.
Symptoms
ANCOVA of the Positive and Negative Syndrome Scale total score at baseline showed a significant main effect of genotype (F=8.1, df=2, 27, p<0.001), with the Val/Met group having a significantly higher score than the other two groups (all p<0.05). Therefore, all subsequent analyses were covaried for the baseline score. MANCOVA of Positive and Negative Syndrome Scale total score (covarying for the score at baseline) showed a significant effect of time (F=19, df=3, 81, p<0.001) but no effect of genotype and no interaction between genotype and time. Similar results were obtained for the three subscale scores of the Positive and Negative Syndrome Scale. The total, positive symptom, and general psychopathology scores of the Positive and Negative Syndrome Scale did not show any differential effect of COMT genotype in terms of response at 8 weeks (all χ2<2.4, df=2, all p>0.25). On the other hand, negative symptom score showed a significant load effect (χ2=6.2, df=2, p<0.04) for the Met allele in terms of the number of subjects who responded (Met-Met: N=4 of 5; Val-Met: N=9 of 17; Val-Val: N=1 of 8). Analogous results were obtained by grouping subjects based on being a Met allele carrier or not (Met/Met and Val/Met, N=22, Val/Val, N=8) (χ2=5.1, df=2, p<0.02).
Working Memory
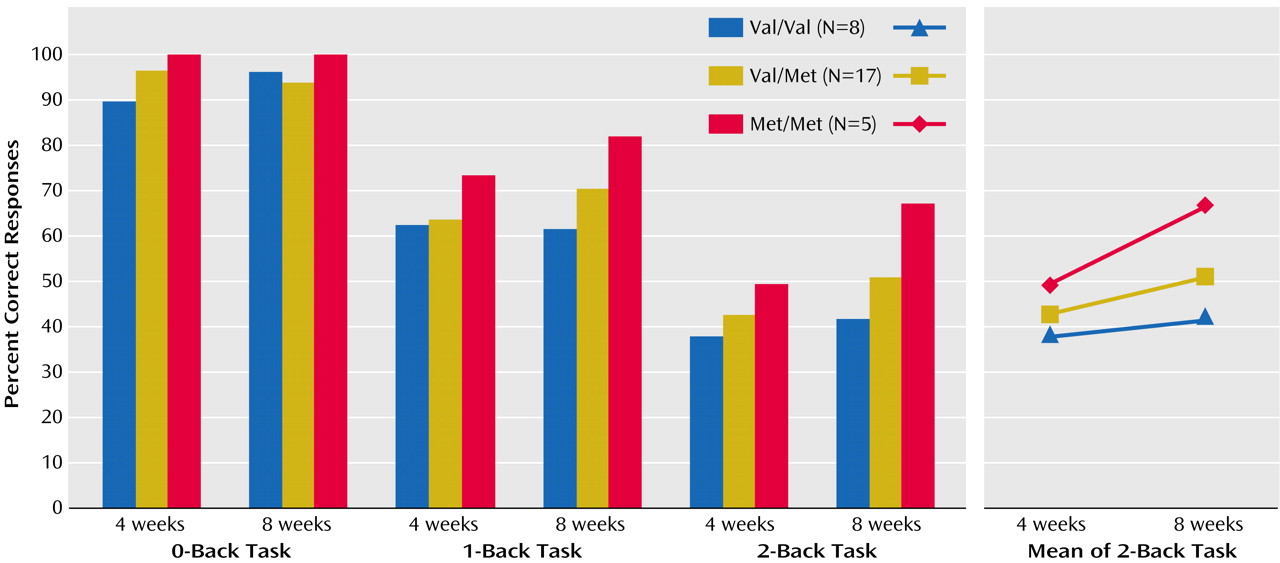
MANCOVA of performance accuracy revealed a significant effect of genotype (F=4.1, df=2, 26, p<0.02), N-back load (F=98, df=2, 54, p<0.001), and time (F=12.6, df=1, 27, p<0.001) and a significant interaction among genotype, N-back load, and time (F=2.7, df=4, 54, p<0.03) (
Figure 1). Post hoc analysis with Tukey’s honestly significant difference indicated that at 4 weeks of treatment there was no significant difference between the three genotype groups. On the other hand, at 8 weeks of treatment there were statistically significant differences in 2-back task performance between the Met/Met group and both the Val/Val (p<0.01) and Val/Met (p<0.01) groups, indicating that the Met/Met patients improved more than the other two groups (
Figure 1). Analogous results were obtained by grouping subjects on the basis of having a Met allele or not. The interaction between genotype, N-back load, and time was again significant (F=4.7, df=2, 56, p<0.01), with Met carriers performing significantly better at the 2-back task at 8 weeks than the other two groups (p<0.01, Tukey’s honestly significant difference).
Similar statistical analyses on reaction time during performance of the N-back working memory tasks revealed a significant effect of N-back load (p<0.02, with 0-back being significantly faster than the other two conditions) but no effect of genotype or time and no interaction .
fMRI
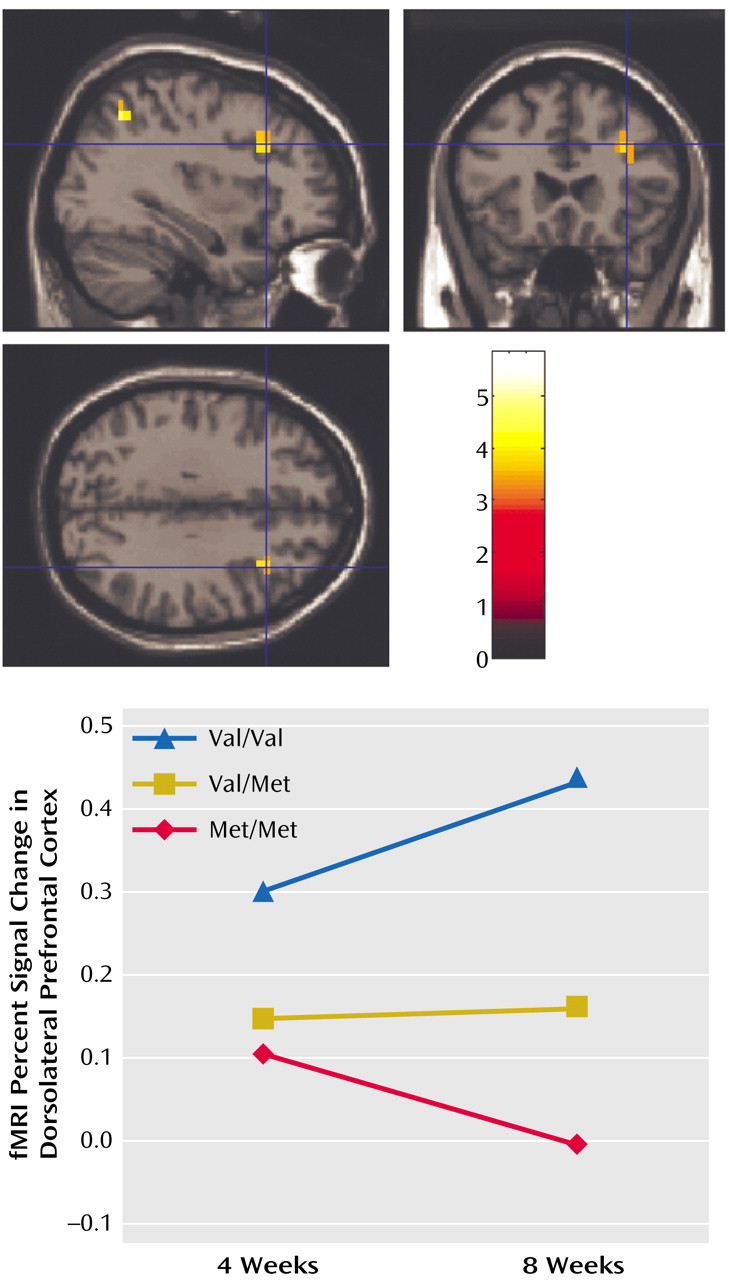
The genotypes of the 20 patients who completed the fMRI part of the experiment were as follows: Met/Met: N=5, Val/Met: N=12, Val/Val: N=3. Second-level (random effects) ANCOVA across working memory load levels showed that at 4 weeks of treatment patients had greater activation in the working memory network, including the dorsolateral prefrontal cortex (Brodmann’s area 9/46), than they did at 8 weeks, suggesting that information processing in the working memory network became more efficient during treatment. The inverse analysis (i.e., 4-week activation less than 8 weeks) did not show any significant differences. Subsequent analyses were performed to evaluate which of the three groups of patients contributed most to the working memory network becoming more efficient over the time period investigated. ANCOVAs of only the 2-back fMRI data showed no difference at 4 weeks. At 8 weeks, locales in the dorsolateral prefrontal cortex (Brodmann’s area 46) and in the parietal cortex (Brodmann’s area 7) showed the predicted genotype effects, with Val/Val individuals having the greatest activation (i.e., being least efficient), followed by Val/Met and then Met/Met individuals (
Figure 2). The same analysis performed on 0-back or 1-back data did not show any significant effect even after lowering the statistical threshold to p<0.01. To corroborate the latter analyses, we performed further analyses. Contrast images of the 4-week versus 8-week contrast for the 2-back condition were compared across genotypes in a second-level random effects analysis to show that the change in fMRI activation from 4 to 8 weeks of olanzapine treatment was different across the genotype groups. Again, locales in the dorsolateral prefrontal cortex (Brodmann’s area 46: x=40, y=19, z=21; p<0.02) showed the predicted genotype effects, revealing that from 4 to 8 weeks, Met/Met patients became more efficient, whereas Val/Val patients did not, and that the differences in directionality of the two groups was significant.
Discussion
The results of the present study suggest that COMT genotype contributes to variation in the cognitive and negative symptom response to olanzapine in patients with schizophrenia. Specifically, we found evidence that Met alleles are associated with greater improvement in negative symptom ratings, in working memory performance, and in prefrontal cortical physiology. Our results are consistent with other evidence that COMT genotype impacts prefrontal cortical information processing and that Met allele carriers have a greater capacity for optimum prefrontal function. The specific mechanism by which this interaction occurs is unclear. Both the Val/Met polymorphism in the COMT gene and the pharmacology of olanzapine affect prefrontal dopamine metabolism, a potentially convergent mechanism of interacting benefit. In this sense, the present data might be taken as the first evidence in humans that increasing prefrontal dopamine with atypical antipsychotics can be clinically beneficial. However, it is also conceivable that the combined effects of olanzapine treatment and the Met allele are less specific, converging through complex effects on the functional stabilization of cortical organization.
The effect of COMT genotype on negative symptoms was less straightforward than the effect on cognition and physiology. While a quantitative analysis did not show any differential effects of COMT on clinical symptoms, a qualitative analysis based on the frequency of good response did show a specific albeit small effect on negative symptoms assessed with the Positive and Negative Syndrome Scale. These data are consistent with the notion that decreased dopamine activity in the dorsolateral prefrontal cortex contributes to negative symptoms in schizophrenia
(26,
47). They are also consistent with several imaging studies that have suggested a specific relationship between reduced levels of dopamine in the dorsolateral prefrontal cortex and negative symptoms
(48,
49). However, it is also clear that the clinical response to olanzapine was broader than the cognitive response, as most patients improved in terms of psychotic symptoms. Thus, although intriguing, the clinical importance of the COMT effect will have to be explored in larger samples of patients.
Consistent with our hypothesis and with a load effect of the Met allele, our data indicate that Met homozygotes improve most, heterozygotes have intermediate improvement, and Val homozygotes improve the least. Indeed, at the level of prefrontal function (i.e., cognition, negative symptoms, and fMRI response), Val/Val individuals show little if any response. The load effect of the Met allele was most evident in performance on the 2-back task, which was the greatest working memory load used in the present study. In other words, when the task becomes more difficult, the effect of COMT genotype becomes more evident. This effect was not present during the control task (0-back), which does not specifically engage the working memory circuitry. Moreover, this effect does not seem to be dependent upon simple speed of processing, since the reaction time data did not show any effect of COMT genotype. These data suggest, therefore, that COMT genotype interacts with olanzapine treatment at the level of working memory capacity. The fMRI results are consistent with the cognitive data and provide potential insight into the underlying mechanism of the effects. The results across all working memory loads show that at 4 weeks of treatment, all patients have higher activation in the working memory cortical network than at 8 weeks of treatment. Since working memory performance improves from 4 to 8 weeks of treatment, the 8-week fMRI data suggest that patients tend to become more efficient over time (less activation required for better performance)
(18,
20). On the other hand, the statistical analyses performed to assess the interaction between genotype and time of treatment showed that while at 4 weeks there was no significant difference between genotype groups, at 8 weeks there was a load effect of the Met allele such that for 2-back task performance the Met homozygotes became most efficient, specifically in the dorsolateral prefrontal cortex. Analogous to the cognition results, no such effect was found for the 0-back or 1-back tasks (data not shown). These data are consistent with earlier studies that indicated patients with schizophrenia are inefficient at high working memory loads when compared with healthy subjects
(18,
20). The studies by Egan et al.
(38) and Mattay et al.
(8) also showed that dorsolateral prefrontal cortex inefficiency is associated with COMT genotype, with Val homozygotes being the least efficient. Our data are consistent with and extend these prior studies by showing that 8 weeks of treatment with olanzapine were more beneficial for Met homozygotes, who became more efficient in the dorsolateral prefrontal cortex at higher working memory loads.
Limitations
Since our study involved only 8 weeks of treatment, we cannot say whether this differential effect will be maintained over longer periods of time. However, since a load effect of the Met allele has already been demonstrated in clinically stable patients
(38), we do not anticipate that longer periods of treatment can reverse the effect of COMT on working memory performance in schizophrenia. Moreover, for clinical reasons we did not acquire fMRI data during working memory at baseline. Therefore, despite the lack of genotype effects on the fMRI data at 4 weeks, the study cannot rule out the possibility that olanzapine’s effects on working memory and prefrontal physiology are in fact present in all of the patients by 4 weeks of treatment. Indeed, it remains possible that the Val/Val group showed an early improvement (within 4 weeks) in response to olanzapine and reached a plateau, whereas the Val/Met and Met/Met groups showed a delayed response to olanzapine that was not evident until the 8th week of treatment. However, this possibility seems somewhat unlikely based on earlier data. In a previous study in clinically stable patients switched to olanzapine, Purdon et al.
(50) reported that the earlier cognitive benefits were evident after 6 weeks of treatment. Also, the putative neurobiology of COMT regulation of dopamine metabolism in prefrontal synapses would make this prediction counterintuitive in patients with schizophrenia. Therefore, even if theoretically possible, it is unlikely that there was an early improvement in our Val/Val group that reached a plateau within 4 weeks of treatment.
As with all longitudinal studies, reliability of the data might confound the results. Manoach et al.
(51) have reported poor reliability of the magnitude and spatial extent of activation during the Sternberg Item Recognition Paradigm
within schizophrenia subjects across scanning sessions. Despite limited test-retest reliability among patients as individuals, averaged over the group, the identical network of structures was activated over time
(51). These authors suggested that it is important to control for sources of variation, both artifactual and intrinsic. We attempted to control as much as possible for a series of factors potentially contributing to variation. The three groups of patients did not differ at any time point in the degree of residuals of motion correction as assessed with SPM 99 (all p>0.40). Moreover, we controlled for task performance in the statistical analysis. Another factor that could possibly contribute to variation is differences in proceduralizing a working memory task over time
(52). However, we failed to find any difference between the three groups of patients over time in terms of reaction time, suggesting that this possibility is not a major concern. Another factor that speaks against the possibility that our data are significantly affected by random error variability is the specificity and consistency of the findings (found in the dorsolateral prefrontal cortex only for 2-back working memory task performance). In other words, it is difficult to imagine that a predicted effect specific to the dorsolateral prefrontal cortex is only due to artifactual systematic variance. Therefore, even if poor test-retest reliability of fMRI working memory activity could be attenuating the sensitivity to improvements in the Val/Val patients, on the same note it would also be true that the improvement observed in the Met/Met patients was likely a conservative estimate of the true improvement that would have been observed if the N-back fMRI activity had perfect test-retest reliability.
Another limitation of our data is that they are based on a relatively small group of patients, particularly those with Met/Met genotypes, who were our best outcome subjects. Therefore, caution is warranted in interpreting our results. However, we have several reasons to believe that our results are robust. First, we examined 30 untreated patients with schizophrenia, a large fraction (N=16) of whom had never been treated with antipsychotics. Patients who have never been treated provide a unique opportunity because the findings are not complicated by issues of chronicity or of pharmacological treatment. Another strength of our study is the longitudinal within-subject design, which is well suited and powerful to address issues related to response to drug treatment. Further, despite the relatively small study group size, the effect size of the difference in performance on the 2-back version of our working memory task in Met homozygotes was 0.85, which, statistically speaking, is a reasonably large effect size. Clinical experience suggests that cognitive improvement in working memory is an infrequent phenomenon in patients with schizophrenia, so that our small Met/Met subgroup may be representative of subjects who show such improvement. Third, the analysis of working memory data performed by grouping subjects according to whether they carry a Met allele or not, i.e., Met-Met and Val-Met subjects (N=22) produced results consistent with that obtained in the three groups of patients. The effect size of these results was smaller (0.52) but still intermediate. Fourth, another strength of our study is the fMRI component, which is internally consistent with the neuropsychological data. Furthermore, the fMRI data represent an unbiased in vivo measure of the change of neuronal activity during performance of the behavioral task. It has to be underlined that the statistical threshold used for the fMRI data is conservative (p<0.001), further speaking to the robustness and genetic specificity of the findings. The effect size of the difference in fMRI signal change in Met homozygotes between 4 and 8 weeks of treatment was 0.88, which again is a large effect size.
In conclusion, our data suggest that 8 weeks of treatment with olanzapine differentially enhances working memory performance and related dorsolateral prefrontal cortex efficiency in patients according to COMT genotype: patients homozygous for the Met allele show greatest improvements (higher performance and reduced dorsolateral prefrontal cortex inefficiency as defined by reduced neuronal activation); patients heterozygous have intermediate improvements; patients homozygous for the Val allele have the least if any improvements. Our data may suggest that increased prefrontal levels of dopamine induced by olanzapine have a greater impact in Met/Met individuals, possibly because the effect lasts longer or crosses a threshold in dopamine signaling that positively modulates intrinsic prefrontal processing. Met/Met individuals also may be more effective in compensating for other deficits in prefrontal information processing that are intrinsic to the disorder. This interpretation is consistent with data suggesting that the COMT Val allele represents a susceptibility allele for schizophrenia further compounding an abnormality in information processing in the prefrontal cortex
(38,
53).