We have recently studied auditory cortical processing in
adults with autism using complex speech-like sounds
(1) and found
less activation of the left temporal word processing network in autistic adults than in healthy comparison subjects. The results obtained in autistic adults suggest a dysfunction of specific temporal regions specializing in the perception and integration of complex sounds
(2). Such stimuli are never recognized as speech; therefore, an abnormal pattern of activation found in autistic adults could reflect basic anomalies of prelinguistic auditory processing rather than a consequence of abnormal language development
(3). Since autism is a developmental disorder, we believed it important to investigate the cortical response to sounds in
children with autism. All reported activation studies in autism have been performed in adults, but no data are available for children, to our knowledge. Therefore, we performed a positron emission tomography (PET) auditory activation study in autistic children using the same speech-like stimuli previously described in the autistic adult study. Autism is associated with abnormal auditory behavior, including aversive reactions to everyday life sounds. In addition, autistic children are often initially misdiagnosed as deaf
(4). An abnormal perception of speech-like sounds in childhood may account for inadequate behavioral response to sounds and thus for language impairments typical of autism.
Method
Eleven autistic children (10 boys) ages 4 to 10 years (mean=6.6, SD=1.6) with primary autism were selected. Autism was diagnosed according to DSM-IV criteria. The Autism Diagnostic Interview—Revised
(5) confirmed the diagnosis: mean social interaction score=27.3 (SD=6.6); mean nonverbal communication score=11.9 (SD=3.5); mean stereotypy score=7.9 (SD=4.7); mean age-onset criteria=4.3 (SD=0.8). Four children had verbal communication; their mean score was 17 (SD=7). Their IQ or development quotient was determined with the WISC-R or Brunet-Lézine test
(6). Their mean IQ or development quotient was 43 (SD=21).
Six children (four boys) ages 3 to 9 years (mean=7.2 years, SD=2.4) with idiopathic mental retardation (mean IQ or development quotient=64, SD=15) were selected as control children since regional cerebral blood flow (rCBF) studies are not performed on normal children in our institute for ethical reasons. The diagnosis of mental retardation conformed to DSM-IV criteria. No etiology has been found after extensive investigations.
We excluded from this study children with known infectious, metabolic, neurological, or genetic diseases; chromosomal abnormality; seizures; and abnormal magnetic resonance imaging (MRI).
All subjects were free of medication. An ethics committee approved this study, and examinations were performed with the written informed consent of the parents.
Relative rCBF was determined from the radioactivity distribution measured with PET (ECAT-EXACT-HR+) after bolus intravenous injections of [
15O]H
2O
(7). The protocol included three rCBF measurements carried out in a single session, performed at 10-minute intervals during three conditions: the first at rest and the second and third during passive listening to complex speech-like sounds
(1).
We used a subset of the synthetic nonverbal speech-like auditory stimuli previously published elsewhere
(1,
3). These stimuli contain spectral maxima (speech-like formants) that change over time. They consist of sounds with a central 200-msec steady-state period surrounded by initial and final changes in spectral maxima frequency. Their acoustic structure was similar to consonant-vowel-consonant, but as stated, normal volunteers never recognized them as speech.
PET studies were performed during sleep induced by premedication (7 mg/kg of sodium pentobarbital) for both autistic and control children to obtain immobility.
rCBF images were analyzed by using Statistical Parametric Mapping 99 for image realignment, transformation into standard stereotactic anatomical space, smoothing, and statistical analysis
(8). State-dependent differences in global flow were covaried by using proportional scaling. Comparisons across conditions were made by using the t statistic subsequently transformed into a normally distributed z statistic in a multistudy design. Two statistical comparisons were performed: 1) a within-group comparison of activation for listening to complex sounds versus resting and 2) a between-group comparison of activations.
Results
Passive listening to speech-like sounds versus resting was assessed in each group independently (z=3.67, df=28, p<0.0005). We found significant activation of the auditory cortex in the bilateral superior temporal gyrus (Brodmann’s area 22) in both groups while subjects were listening to speech-like stimuli. However, the activation pattern was different between two groups. The control children activated the superior temporal cortex bilaterally with left-biased asymmetry, as we have previously observed in normal adult comparison subjects
(1). This left dominance was not observed in the autistic group. In addition, autistic children had additional significant activation outside the auditory cortex: the left temporal pole (Brodmann’s area 38), the bilateral cingulum (Brodmann’s areas 32 and 24), the bilateral posterior parietal (Brodmann’s area 19), the cerebellar hemispheres, and the brainstem.
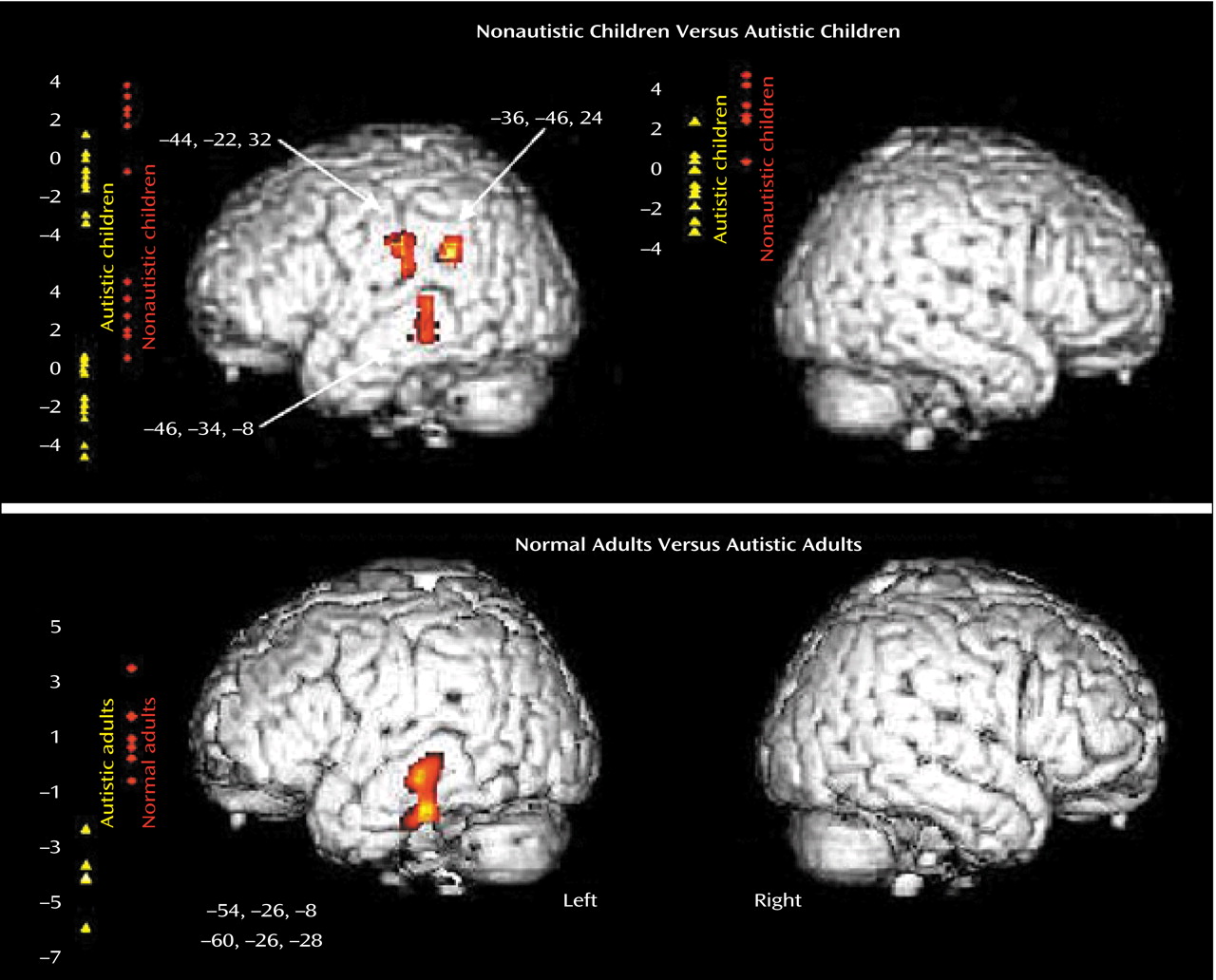
The direct comparison showed a significant difference between the autistic and control groups in three areas in the left cortical network. The left middle temporal gyrus (Brodmann’s areas 21 and 39) and the left precentral gyrus (Brodmann’s area 43/6) (z>3.41, df=28, p<0.001) were significantly
less activated in the autistic children than in the nonautistic control children (
Figure 1, top). In addition, we obtained exactly the same results by performing an additional analysis and excluding the girls from the autistic and control groups. We found no region that was significantly more activated in the autistic group.
Discussion
The present findings of an abnormal cortical pattern of auditory activation in autistic children are similar to those previously described in autistic adults when using the same auditory paradigm
(1). The direct comparison between the two groups of children while they were listening to speech-like stimuli revealed
less activation in left speech-related areas, including Wernicke’s area, in autistic children. Similar results were found in autistic adults in relation to normal comparison adults
(1). The middle and inferior temporal gyrus (Brodmann’s area 21) were significantly
less activated in autistic adults than in comparison subjects.
Figure 1 shows the similarity of the results obtained in children and adults with autism compared to nonautistic age-matched control subjects.
By comparing speech-like sounds to the resting state in each group independently, we also observed remarkable differences in the activation pattern between the two groups. Autistic and nonautistic children activated the temporal auditory cortex bilaterally, but the nonautistic children activated the left side more than the autistic group. This left dominance was also observed in normal adults
(1). In addition, we observed in autistic children diffuse activation outside the temporal lobe, located in the bilateral cingulum, the posterior parietal, the brainstem, and the cerebellum. Thus, in autistic children, listening to complex sounds induced an abnormal cortical activation including an aberrant functional network. This may explain why autistic children have exaggerated behavioral responses to sounds, one of the most pronounced signs of autism. However, we did not observe in autistic children the abnormal right frontotemporal activation previously described in autistic adults
(1). This difference may be related to relatively small group sizes in both studies or, more likely, may indicate that a marked right-side asymmetry emerges in autistic patients across their development.
To perform activation studies in typical mentally retarded autistic children, we needed to use premedication. Even under sedation, we observed in both autistic and nonautistic children a bilateral activation of the temporal auditory cortex that was induced by complex sounds. These results suggest that the sound’s perception with the subjects premedicated is effective and opens new perspectives about the passive activation paradigm in children with developmental disorder. Likewise, auditory functional MRI in pentobarbital-sedated children showed auditory cortical activation
(9). Since both groups were studied under same conditions, the findings should reflect true differences. However, the present results need to be replicated in a larger well-matched population of autistic and control children.
The present results, together with the adult study, indicate that autism in both children and adults is associated with a dysfunction of specific temporal regions specialized in perception and the integration of complex sounds. The areas found to be less activated by complex sounds in autistic children (Brodmann’s areas 21 and 39) are thought to be auditory associative areas that are involved in word processing
(10) and are also presumed to act as an interface between word perception and long-term representations of familiar words in memory
(10,
11). In the dominant hemisphere for language, these areas play a critical role in the ability to understand and produce meaningful speech. Thus, a dysfunction of left speech-related cortical areas could be at the origin of the language developmental impairments observed in autism.
Finally, the
resting bilateral temporal lobe dysfunction previously described in autistic children
(12,
13) could be implicated in the developmental disorganization of temporal neural circuits leading to severe perturbation in the auditory activation pattern.
In conclusion, the present results extend to autistic children previous findings obtained in adults and are compatible with the general hypothesis of disturbances in the establishment of neural circuits in the autistic brain.