Several reports have described a subset of relatives of individuals with autism who display social and language deficits that are qualitatively similar to those of the autistic proband but milder in severity
(5,
6). This constellation of impairments, which relatives of probands exhibit at higher rates than do people in the general population, has been characterized as the broader phenotype for autism
(7–
11). Although evidence for the broad phenotype in families varies across studies
(3,
12,
13), investigations of monozygotic twins
(7) and of families with multiple-incidence autism
(9,
10) have demonstrated support for the concept.
In comparisons of families with multiple-incidence autism to families with Down’s syndrome, Piven and colleagues reported significantly higher rates of social and communication deficits and stereotyped behaviors
(9) and of psychiatric disorder
(10) in the families of individuals with autism. Following up on a study by Folstein and Rutter
(2), Le Couteur and co-workers
(7) evaluated a group of 28 monozygotic pairs and 20 dizygotic same-sex pairs in whom one or both twins had autism, and they found social and communication deficits in 77% of the monozygotic co-twins (compared to 5% of the dizygotic twins) who were discordant for autism. Rutter and colleagues
(8) have suggested that in such families a genetic substrate for autism may produce the social and cognitive deficits that are included in the broader phenotype but that an interaction between the genetic substrate and unknown deleterious environmental factors culminates in a “second hit” that ultimately produces the narrow phenotype for autism.
Little is known about the differences in neuroanatomy that could ultimately be associated with the narrow and broader behavioral phenotypes for autism. The findings from imaging studies of individuals with the narrow phenotype have not been consistent. Although several studies have indicated higher than normal overall brain volume
(14–
16) and high regional (parietal, temporal, occipital) brain volumes
(17), a cross-sectional study demonstrated that high volumes of cerebral gray, cerebral white, and cerebellar white matter in early childhood were followed by low volumes in late childhood and adolescence
(18).
In a case report that compared the neuroanatomy of one monozygotic twin pair discordant for the narrow phenotype of autism to five age- and gender-matched unaffected comparison subjects, we found dysfunction of two separate brain clusters
(19). Relative to the comparison subjects, both the twin with autism and his co-twin exhibited lower volumes in the frontal lobe and the superior temporal gyrus. The twin with autism also exhibited lower volumes in noncortical structures, including the caudate nucleus, the amygdala, the hippocampus, and the cerebellar vermis. On the basis of the findings from that preliminary report, in the present study we investigated the neuroanatomic phenotypes of 16 pairs of monozygotic twins of which seven twin pairs were concordant and nine twin pairs were discordant for the narrow phenotype for autism.
Method
Study Group
The original group consisted of 54 children, including 18 pairs of monozygotic twins and 18 age- and gender-matched unaffected peers. One twin pair was excluded from the analyses for this report because their scans were acquired in the sagittal plane, thereby producing segmented data that were not comparable to segmented data acquired in the coronal plane
(20). A second twin pair was excluded for reasons to be described.
The final study group consisted of 48 children, including 16 pairs of monozygotic twins (N=32) and 16 age- and gender-matched unaffected peers. Fourteen of the twin pairs, and their matched comparison subjects, were boys. The mean age of the twin pairs was 8.4 years (SD=2.6, range=5.3–13.8). The mean age of the comparison subjects was 8.3 years (SD=2.4, range=5.4–13.9).
Families were recruited through the Autism Society of America, the National Alliance for Autism Research, the Kennedy Krieger Center for Autism and Related Disorders, and clinical neurologists at Kennedy Krieger Institute and Johns Hopkins Hospital. Subject recruitment followed a two-step process. A medical history form and the Autism Behavior Checklist
(21) were initially mailed to interested families in order to screen twin pairs for potential participation in the study. Subjects with a genetic disorder or a history of status epilepticus were excluded. The behavioral criterion for potential participation in the study was based on the Autism Behavior Checklist: if at least one of the twins scored above 57 (representing the narrow phenotype for autism) on the Autism Behavior Checklist, the family was contacted by telephone by a project staff member. The Autism Diagnostic Inventory—Revised
(22) was administered (separately for each co-twin) in a series of telephone interviews with a parent of the twins by a project staff member (L.A.S.) who was trained in the reliable administration of the instrument. If the diagnosis of autism was confirmed by the Autism Diagnostic Inventory for at least one co-twin of each twin pair, the family was invited to the Kennedy Krieger Institute for additional assessment. At our research center, the Autism Diagnostic Observational Scale—Generic
(23) was administered to each co-twin by a reliable examiner (L.A.S.). The final diagnosis of each child was based on both the Autism Diagnostic Inventory and the Autism Diagnostic Observational Scale. In order to qualify for the diagnosis of autism, each child had to meet the criteria for autism on the Autism Diagnostic Inventory and score within one point of meeting the criteria for autism on the Autism Diagnostic Observational Scale.
No twin pair that qualified for the study on the basis of the Autism Behavior Checklist was then excluded on the basis of the Autism Diagnostic Inventory. However, for three twin pairs, the two instruments produced discrepant results regarding concordance between co-twins: one twin pair identified as discordant by the Autism Behavior Checklist (i.e., only one co-twin in the pair obtained a score higher than 57) was concordant according to the Autism Diagnostic Inventory, and two twin pairs identified as concordant by the Autism Behavior Checklist were discordant according to the Autism Diagnostic Inventory. In addition, one of the twin pairs whose scores indicated concordance on the Autism Behavior Checklist and discordance on the Autism Diagnostic Inventory was ultimately excluded from the study on the basis of the Autism Diagnostic Observational Scale, on which neither co-twin scored within the autistic range.
The unaffected singleton comparison subjects, individually matched by age and gender to each twin pair, were recruited from the office of a local pediatrician. Letters describing the study were sent by the pediatrician to parents of patients who, according to their medical records, did not have a history of developmental delays, emotional disturbance, or learning disabilities. Parents then contacted one of us (W.R.K. or C.P.B.) if they were interested in participating in the study. They were screened first by telephone (in order to rule out current emotional, developmental, or learning difficulties) and then invited to participate in the study if the telephone interview confirmed that their children met the inclusion criteria. Developmental diagnoses, including language delay, and psychiatric diagnoses (e.g., attention deficit hyperactivity disorder) were ruled out by using parental report and child psychiatric checklists (e.g., the Child Behavior Checklist). Accordingly, none of the comparison subjects had any known neurologic, developmental, learning, or psychiatric disorder.
Parents of all subjects signed informed consent forms approved by the institutional review board of Johns Hopkins Hospital.
DNA fingerprinting probes were used to confirm zygosity in all twin pairs. Eight independent loci were tested for each twin pair. The DNA profiles for all twin pairs were statistically identical at every locus, indicating that the probability of monozygosity for each twin pair was approximately 99.99%.
Image Acquisition and Processing
Magnetic resonance imaging (MRI) was performed on a 1.5-T GE Signa scanner (GE Medical Systems, Milwaukee). A three-dimensional high-resolution MRI scan was acquired for each subject by using a T1-weighted spoiled gradient echo sequence (TR=35–45 msec, TE=5–7 msec, number of excitations=1) with a flip angle of 45°, matrix size of 256×128, and field of view of 20–24 cm, yielding 124 coronal slices with a slice thickness of 1.5 mm and an in-plane resolution of 0.131 mm.
The image data were imported into the program BrainImage (A.L. Reiss, Stanford University; http://spnl.stanford.edu/tools/brainimage.htm) for visualization, processing, and quantitation. The data were prepared for measurement by correcting for radio frequency inhomogeneity artifacts, removing voxels representing nonbrain material, and reformatting the images to produce data sets representing cubic voxels
(24). These data sets were opened into the multiplanar visualization module of BrainImage, so that three orthogonal representations of the data could be viewed simultaneously.
Image Measurement
The isolated brain tissue was subdivided into cerebral lobes and subcortical, brainstem, and cerebellar regions according to a method based on the revised Talairach atlas
(25) that is specific for measurement in pediatric study groups
(26–
28). With this approach, high levels of sensitivity and specificity are achieved for all volumes of lobar brain regions based on the revised Talairach atlas
(28). Each region was then segmented into gray, white, and ventricular compartments by using a constrained fuzzy algorithm that assigns voxels to one or more tissue categories on the basis of intensity values and tissue boundaries
(29).
Statistical Analyses
Intraclass correlation coefficients (ICCs) were calculated in order to determine the degree of concordance in brain volumes in both the concordant and discordant twin pairs. We used r-to-z transformations to compare the magnitudes of the ICCs for the two groups. Multiple analyses of covariance (ANCOVAs) were conducted in order to compare the brain volumes of the discordant twin pairs and singleton comparison subjects. Although limiting our volumetric analyses to the discordant pairs and comparison subjects reduced statistical power to some extent, this strategy permitted us to focus specifically on the extent to which discordance in clinical phenotype was reflected in differences in cerebral, cerebellar, and ventricular regions of interest. These analyses were conducted with diagnostic group as the independent variable, age and whole brain volume as the covariates, and volumes of neuroanatomic regions of interest as the dependent variables. Post hoc analyses were conducted with the Bonferroni-Dunn statistic to correct for multiple comparisons. Linear regression analyses were conducted in order to determine the relation between brain volumes and severity of autism (represented by scores on the Autism Diagnostic Inventory and Autism Diagnostic Observational Scale) in all twins with autism and their co-twins.
Results
Behavioral Phenotype
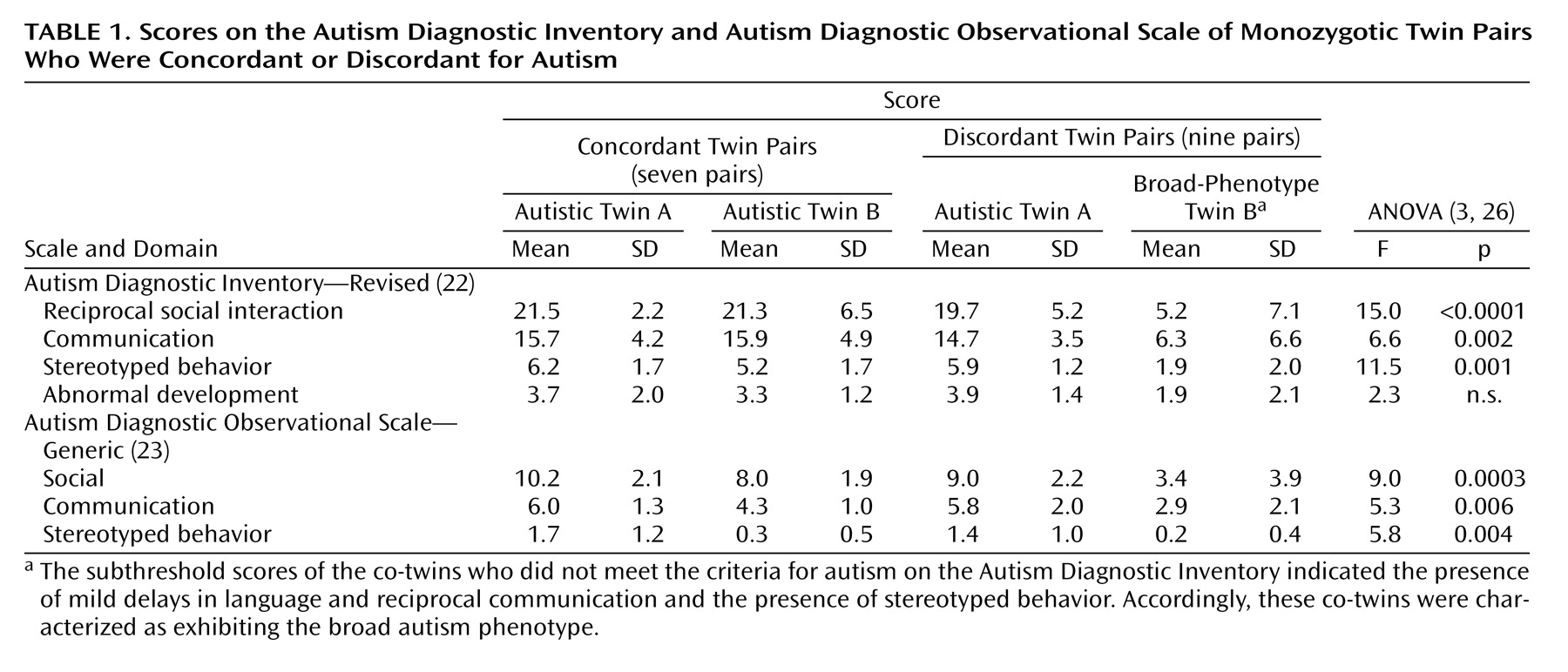
Seven pairs of twins had scores on the Autism Diagnostic Inventory and Autism Diagnostic Observational Scale that qualified both of them for the diagnosis of autism; those twin pairs were designated as concordant for the narrow phenotype for autism. The remaining nine pairs of twins were designated as discordant.
Analyses of variance were conducted to compare the scores on the Autism Diagnostic Inventory and Autism Diagnostic Observational Scale (
Table 1) of the twin pairs. The differences between scores of the twins within the concordant pairs were not significant. Within the discordant twin pairs, Bonferroni-Dunn post hoc analyses revealed that the autistic co-twin received significantly higher scores on all but one summary measure; the exception was the rating of abnormal development on the Autism Diagnostic Inventory.
Within the discordant pairs, six of the nine co-twins who did not meet the criteria for autism on the Autism Diagnostic Inventory were reported by a parent to have exhibited language or social delays before participation in the study. In at least three cases, the delays were severe enough to warrant a diagnosis (before enrollment in the study) of developmental delay or pervasive developmental disorder. Two of the three remaining children whose histories were negative for social and language delay nevertheless exhibited subthreshold scores on the Autism Diagnostic Observational Scale that indicated mild impairment of communication and social skills. According to these criteria, the co-twins of the twins with autism were characterized as exhibiting the broad autism phenotype. We refer to these co-twins as “broad-phenotype co-twins.” (It should be noted, however, that one twin pair was discordant for both traditional autism and the broader phenotype.)
The mean age of the concordant twin pairs was 9.6 years (SD=2.6), and the mean age of the discordant pairs was 7.6 years (SD=2.4). This difference was not statistically significant.
Valid intellectual testing was completed on all comparison subjects, as well as on eight of the 14 twins in the concordant pairs and 17 of the 18 in the discordant pairs. (IQ testing could not be completed on the remaining seven children because of behavioral, attentional, and severe language deficits.) The IQs of the co-twins in the concordant pairs did not differ significantly (p=0.21) from each other, whereas the IQs of the co-twins in the discordant pairs did (t=–2.98, df=13, p=0.01). The IQs of the twins in the concordant and discordant pairs differed significantly from those of the comparison subjects. The twins in the concordant pairs had a mean IQ of 63.2 (SD=22.2), the co-twins with autism in the discordant pairs had a mean IQ of 69.6 (SD=18.7), the broad-phenotype co-twins in the discordant pairs had a mean IQ of 94.4 (SD=13.4), and the comparison subjects had a mean IQ of 123.6 (SD=9.7). After reviewing school records of the participants whose IQs we were unable to assess directly, we found that the frequency of mental retardation was somewhat higher for the twins in the concordant pairs (11 of 14 twins) than for the twins affected with autism in the discordant pairs (three of nine twins) (χ2=5.25, df=3, p=0.07).
Comparison of Concordant and Discordant Twin Pairs
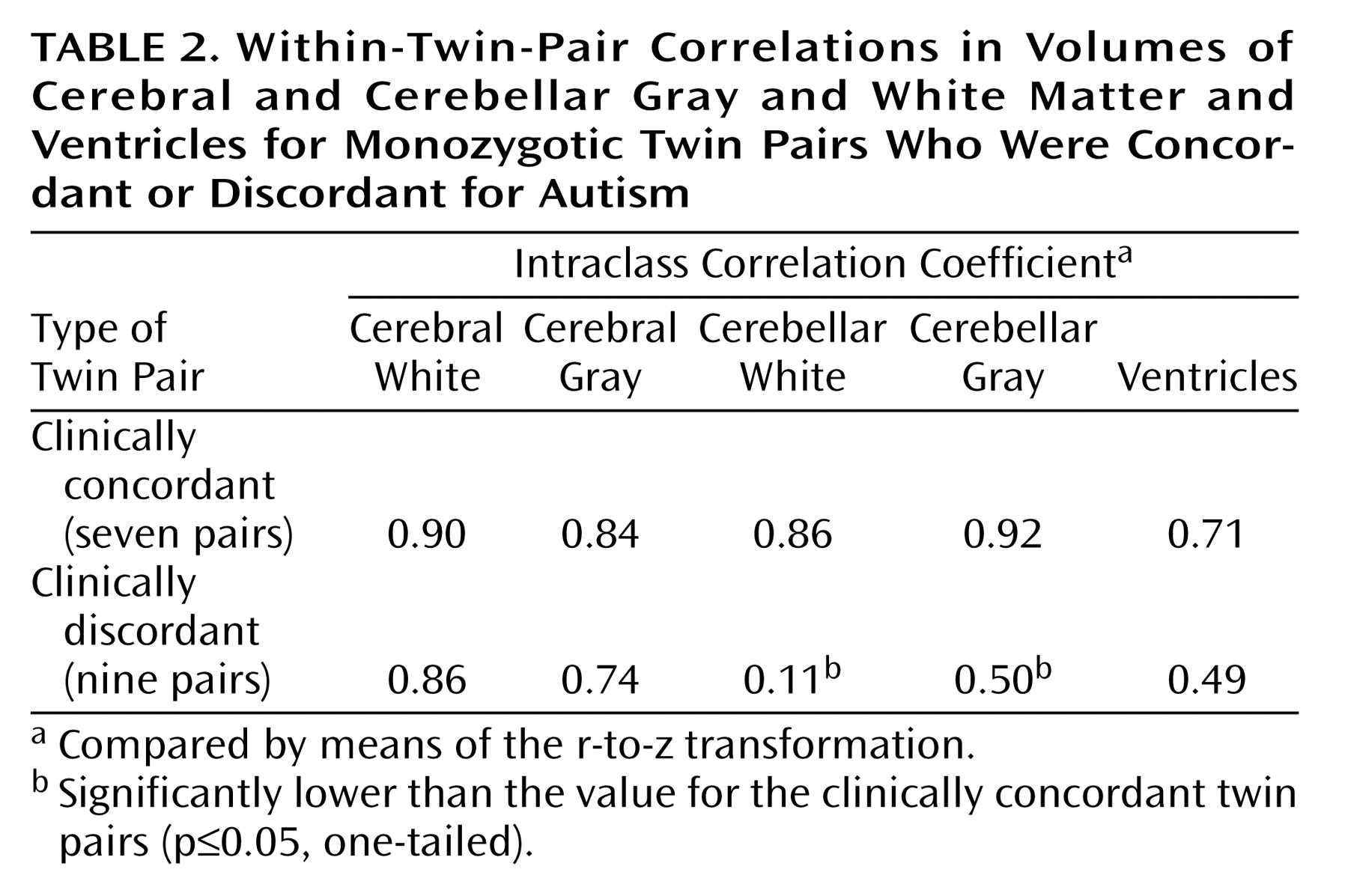
As already stated, the degree of neuroanatomic concordance within twin pairs was determined with the ICC. The Fisher r-to-z transformation was then used to compare the difference in magnitude of the ICCs for the clinically concordant and discordant pairs. As
Table 2 illustrates, the ICCs for cerebral gray and white volumes were high for all sets of twin pairs, regardless of degree of concordance in clinical phenotype. In contrast, the ICCs for gray and white cerebellar volumes were significantly lower for clinically discordant twin pairs than for clinically concordant twin pairs. The ICCs for ventricular volumes were somewhat (although not significantly) lower for the clinically discordant pairs than for the concordant pairs.
Neuroanatomic Differences Between Twins in Discordant Pairs
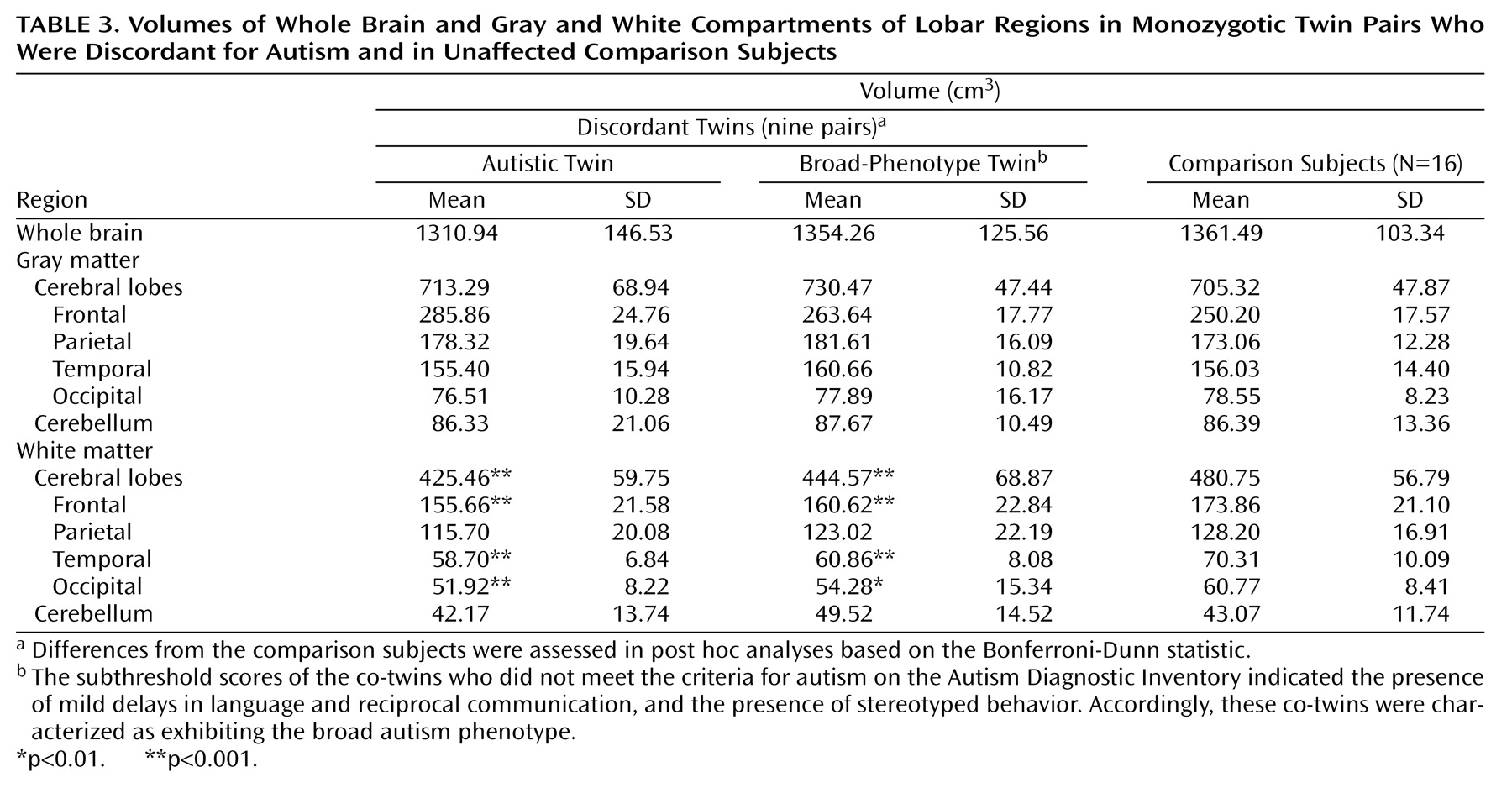
The following analyses were conducted with data from a subgroup consisting of the nine discordant twin pairs and the total group of 16 unaffected comparison subjects. Three groups were compared: co-twins with autism, broad-phenotype co-twins, and unaffected comparison subjects (
Table 3). The groups did not differ significantly in whole brain volume. At the cortical level, cerebral gray and white volumes were compared with ANCOVAs, with whole brain volume entered as the covariate. (Although the group differences in whole brain volume were not significant, it was included in the model in order to investigate the extent to which its variation accounted for differences in specific tissue and regional volumes.) Cerebral gray matter volume did not differ between groups. Cerebral white matter volume was 11.5% lower in the co-twins with autism and 7.5% lower in the broad-phenotype co-twins than in the comparison subjects. Both diagnostic group (F=9.2, df=2, 1, 30, p=0.0008) and whole brain volume (F=212.9, df=2, 1, 30, p=0.0001) contributed significantly to the variance in cerebral white matter volume. Follow-up multiple ANCOVAs were conducted to determine whether specific lobar regions accounted for the differences in total cerebral white matter volume among the children with autism, their broad-phenotype co-twins, and the unaffected comparison subjects. Diagnostic group (autistic co-twin, broad-phenotype co-twin, comparison subject) was entered as the factor, whole brain volume was entered as a covariate, and frontal, parietal, temporal, and occipital white matter volumes were entered as dependent variables. The Wilks’s lambda values for both diagnostic group (lambda=0.46, F=3.3, df=8, 54, p=0.004) and whole brain volume (lambda=0.11, F=54.1, df=4, 27, p=0.0001) were significant. Relative to the typically developing children, both the autistic and broad-phenotype co-twins had significant smaller frontal white matter (F=4.8, df=2, 1, 30, p=0.02), temporal white matter (F=11.0, df=2, 1, 30, p=0.0003), and occipital white matter (F=4.3, df=2, 1, 30, p=0.03).
The cerebellar gray matter, cerebellar white matter, and ventricular volumes of the three groups (autistic co-twins, broad-phenotype co-twins, comparison subjects) were compared with an ANCOVA in which whole brain volume was entered as a covariate. The cerebellar gray and white matter volumes did not differ between groups. Both diagnostic group (F=8.2, df=2, 1, 30, p=0.002) and whole brain volume (F=17.8, df=2, 1, 30, p=0.0002) contributed significantly to the variance in ventricular volume. Planned comparisons indicated that these findings were due to significant higher ventricular volume in the co-twins with autism than in the comparison subjects (p=0.002, Bonferroni-Dunn statistic).
Association Between Behavioral Phenotype and Brain Volumes
Data from all twin pairs were used for these analyses. The association between brain volumes and the behavioral phenotype for autism was determined by calculating the within-twin-pair difference in cerebral gray matter volume, cerebral white matter volume, cerebellar gray matter volume, and cerebellar white matter volume. The resulting difference scores were correlated with within-twin-pair difference scores derived from the sum of the communication and social scores on the Autism Diagnostic Observational Scale. (The summary score was used in order to reduce the number of comparisons.) The difference score for the Autism Diagnostic Observational Scale was not associated with the difference score for either cerebral gray or white matter volume. There was a nearly significant association between the Autism Diagnostic Observational Scale difference score and the difference score for cerebellar gray matter (r=0.48, N=16, p=0.07) and a significant association with the difference score for cerebellar white matter (r=0.55, N=16, p=0.04).
Discussion
The present study is the first, to our knowledge, to use a design of monozygotic twins concordant and discordant for the narrow phenotype for autism to investigate neuroanatomic variation in children with autism spectrum disorder. We found that within discordant pairs, both the autistic and broad-phenotype co-twins exhibited lower white (but not gray) matter volumes in the frontal, temporal, and occipital lobes than did unaffected comparison subjects. The finding that
both autistic and broad-phenotype co-twins exhibited low cerebral white matter volume supports our hypothesis and is consistent with the deficits in executive function, higher-order cognitive processing, and language that characterize children with autistic spectrum disorders. Although a caveat to these findings is the potential contribution of the large discrepancy in IQs between the twin pairs and comparison subjects, previous volumetric studies of children with autism provide a developmental context in which to place these findings. Several studies
(18,
30,
31) give cross-sectional evidence for an overproduction of cerebral gray and white matter in very young children (<5 years of age) with autism that appears to give way to hypoplasia or “normalization” of gray and white matter in children as they approach adolescence. Our subjects were somewhat older than the subjects used in at least one of these studies, and therefore we may be observing the effects of abnormally slowed cerebral white matter growth only.
It is not clear, however, why the autism-specific low volume was limited to white matter. It has been hypothesized that autism is associated with impairment of structural
(32) or functional
(33–
35) connectivity between specific regions of the brain. In a PET study, Castelli and colleagues
(35) observed that adults with autism or Asperger syndrome exhibited low functional connectivity between the extrastriate cortex and the superior temporal sulcus at the temporoparietal junction while viewing animated sequences that elicited the description of mental states. In a spectroscopy study, Murphy and coworkers
(36) found high concentrations of
N-acetylaspartate and choline in the prefrontal cortex of individuals with autism. Whereas
N-acetylaspartate is associated with neuronal integrity, choline is found in large concentrations in myelin sheaths. Accordingly, these differences may represent alterations in axons that derail the efficient transfer of information from the prefrontal cortex to other brain regions. Future studies using diffusion tensor imaging are needed to confirm the presence of alterations in the axonal connections of individuals with autism; potential findings from such studies coupled with the present findings of structural abnormalities in white matter would support the hypothesis of impaired connectivity in autism.
The finding that co-twins did not differ from each other or from comparison subjects in gray or white cerebellar volume was unexpected, although not surprising in light of the long-standing controversy over the extent to which the cerebellum is affected in autism. Differences between individuals with autism and comparison subjects in the volumes of cerebellar hemispheres and in the area of the cerebellar vermis have not been found consistently. Although several studies have shown that either the tissue
(16,
31) or the white matter
(18) of the cerebellar hemispheres is higher in individuals with autism, the differences are not consistently disproportionate to differences in whole brain volume. The current findings (including the results from comparisons of concordant and discordant twin pairs, discussed in the following) suggest that the precise nature of abnormalities of the cerebellum in autism is still not well understood.
Our hypothesis that clinically concordant twin pairs would exhibit a higher degree of neuroanatomic concordance than clinically discordant twin pairs was supported to a large extent. Whereas cerebral gray and white volumes were highly correlated for both clinically concordant and discordant pairs, cerebellar and ventricular volumes were more highly correlated for clinically concordant pairs than for clinically discordant pairs. This finding of neuroanatomic discordance in monozygotic twin pairs would not have been predicted from studies of unaffected, typically developing monozygotic twin pairs, for whom both cerebral and cerebellar volumes have been shown to be highly correlated
(37–
39). Previous studies of unaffected monozygotic twin pairs have led to the conclusion that key aspects of both cerebral and cerebellar development are under tight genetic control in typical subject groups. As we will discuss, however, our findings suggest that for individuals with autism, the cerebellum may not be under as high a degree of genetic control. Moreover, the within-twin-pair differences in cerebellar white and gray volumes (but not cerebral volumes) were associated with within-twin-pair differences in Autism Diagnostic Observational Scale scores, suggesting that cerebellar (as opposed to cerebral) anatomy may serve as an important mediating feature between genetic liability and the expression of behavioral features of autism.
We do not know the etiology or pathology of these phenotypic differences in neuroanatomy and associated behavior between individuals whose genetic makeup is identical. One explanation is that
within-pair differences in the expression of susceptibility genes could lead to differences in the regulation of brain development. The overexpression of neurotrophins, which regulate axonal and dendritic development as well as synaptogenesis
(40), has been found in the blood of infants who were later diagnosed with autism spectrum disorder
(41). Accordingly, development of cerebellar tissue could be compromised in co-twins who overexpress neurotrophins, undermining the development and efficiency of the neuronal connectivity necessary for adequate language and social development. In support of this notion is the finding that brain-derived neurotrophic factor, one of the neurotrophins found at high levels in children who later were diagnosed as autistic, is a growth factor for neuronal populations such as cerebellar-based Purkinje cells
(41). (However, levels of brain-derived neurotrophic factor were also high in children later diagnosed with mental retardation without autism, so the association may not be specific.)
As suggested earlier, an alternative explanation is that in order for severe autism to develop in one twin, genetic susceptibility to the disorder must be accompanied by deleterious environmental events that do not necessarily affect the child’s co-twin
(8). These environmental events could include prenatal or perinatal factors. Previous studies have been inconsistent in their support of an association between adverse prenatal and perinatal events and autistic spectrum disorders
(42–
45), although a more recent study supports such an association
(46). Although this association is not robust, or specific to severe autism, these environmental factors could include nutritional variation due to malposition of one co-twin
(47), prenatal trauma in the form of reduced blood flow or oxygen, or twin-to-twin transfusion resulting in significantly lower birth weight in the more severely affected co-twin. These conditions could affect the extent to which certain brain regions develop concordantly in co-twins
(48). Postnatal factors could include delivery complications or respiratory distress affecting one co-twin to a greater extent than the other. Although the co-twins in our study did not appear to differ in the incidence of these events, our data were based on retrospective parental responses to a general checklist of perinatal and postnatal problems as well as medical interventions required. These responses were most likely not as reliable as review of hospital birth records and pose a limitation to this study.
The finding that cerebellar volumes diverge in discordant twins and are associated with the severity of the behavioral phenotype for autism suggests that variability in cerebellar morphometry may be mediated by nongenetic risk factors. Although this interpretation would not be consistent with findings from previously cited studies of the cerebellum
(37,
39), which has been found to be under significant genetic control in normal subjects, it is possible that the genes that underlie the phenotype for autism affect the cerebellum in a way that increases its susceptibility to environmental risk factors. Moreover, insofar as cerebellar development is experience dependent
(49,
50), the less affected (broad-phenotype) co-twin’s capacity to adapt to environmental stimuli may result in further “experience-guided” brain development
(51), including greater cerebellar-cortical connections. Accordingly, preservation of cerebellar integrity in one co-twin may result in a cascade of events that permits the development of a relatively mildly affected neuroanatomic and behavioral phenotype in that co-twin.
Neuroanatomic differences between concordant and discordant twin pairs can be elucidated further with studies that use larger groups of not only monozygotic but also dizygotic twin pairs. Such studies, which should also include unaffected twin pairs, would help identify the relative contributions of genetic liability and environmental risk factors for autism. In addition, environmental risk factors, including potential prenatal, perinatal, and postnatal disruptions in development, should be integrated systematically into such studies. Moreover, the inclusion of comparison subjects with IQs in the average range would reduce the potential contribution of IQ to findings of anatomic differences. Continuing to clarify and differentiate the neuroanatomic pathways that subserve the entire spectrum of autistic disorders ultimately will permit specific and appropriately timed clinical interventions to children who display the full range of phenotypic expression of this disorder.