Attention deficit hyperactivity disorder (ADHD) is one of the most common but poorly understood developmental disorders of childhood. A defining characteristic of ADHD is ineffective control of behavior in cognitive, emotional, and social domains
(1). Cognitive control, the ability to voluntarily constrain actions in a goal-directed manner, comprises at least two operations: the inhibition of inappropriate but prepotent responses (response inhibition) and the suppression of interfering responses (interference suppression). Response inhibition, as measured by errors of commission on tasks requiring cue-guided inhibition of a prepotent response, is reduced in subjects with ADHD
(2,
3). Interference suppression, as measured by erroneous or slower response to a target stimulus because of interference from competing responses, is also reduced in subjects with ADHD
(4). The neural basis of reduced cognitive control in ADHD is not well understood.
Delayed or abnormal maturation of the frontal cortex and related circuitry is suspected to underlie reduced cognitive control in ADHD. Improved response inhibition
(5) and interference suppression
(6) during typical development is paralleled by changes in myelination and synaptic organization
(7,
8) and in functional activation of the frontal cortex
(9–
11). Volumetric studies have shown abnormally reduced right frontal lobes
(12) and striatal
(13) and cerebellar
(14) structures in ADHD subjects. Further, functional activation of frontal-striatal structures was atypical in ADHD. During interference suppression, ADHD adults have shown reduced anterior cingulate activation but greater frontal-striatal activation relative to comparison subjects
(15). During response inhibition, ADHD children exhibit reduced frontal or striatal activation
(2,
16,
17), but in one study frontal activation was greater or did not differ from that of comparison children
(2). Patterns of group differences across studies likely relate to variable task demands. Atypical activations likely reflect anatomical abnormalities, since response inhibition performance has been shown to correlate with volumes of right frontal lobe and caudate in ADHD and comparison children
(18).
The present study addresses significant gaps that remain in our understanding of the neural basis of impaired cognitive control in ADHD. First, the neural basis of impaired interference suppression has been examined in adult subjects with ADHD
(15) but not in preadolescents. We hypothesized that findings from adults would not extend to children with ADHD, since the neural basis of interference suppression differs substantially between typically developing children and adults
(11). Second, most subjects in past studies were chronically medicated. We minimized contribution of prolonged medication to any differences between ADHD and healthy children by including primarily children who had never been medicated. Third, neuroimaging studies of ADHD have examined response inhibition in isolation
(2,
17) rather than in concert with other cognitive control operations. Tasks that evoke flexibly multiple control operations may provide a different, and perhaps ecologically valid, characterization of cognitive control in ADHD. Fourth, most neuroimaging studies of ADHD have used block designs that cannot selectively visualize successful performance. Thus, functional brain differences between ADHD and comparison subjects may have reflected greater frustration or arousal that may accompany error-prone performance of ADHD subjects. We included a rapid event-related design that varied the order and timing of trials in an optimal manner for selective visualization of individual trials. One prior study also selectively visualized correct trials but without an optimized trial design such as ours
(17). Selective visualization of accurate cognitive control allows for better characterization of differences in the neural substrates of cognitive control.
The present study examined the neural basis of both response inhibition and interference suppression within a modified flanker task in a single group of preadolescent children, most of whom were chronically unmedicated. Our study of typical development has revealed that adults recruit the ventral frontal cortex during both cognitive control functions, whereas regional recruitment within the frontal cortex varies for the two operations in children
(11). On the basis of past studies highlighting frontal abnormalities in ADHD, we hypothesized that ADHD children would recruit the same frontal regions as healthy children but to a lesser extent. However, no prior study of ADHD has examined more than one cognitive control function within the same task. It is likely that ADHD children’s cognitive impairments may be enhanced under those circumstances, revealing functional abnormalities that are not limited to the frontal cortex.
Method
Subjects
Ten children (seven boys and three girls; mean age=8.8 years, SD=0.9) with combined-type ADHD and 10 age- and gender-matched comparison subjects (mean age=9.2 years, SD=1.3) participated for monetary compensation. The comparison subjects were a subset of a previously described comparison group
(11). After complete description of the study to subjects, written informed consent from parents and assent from children were obtained. At the time of scanning, seven of the ADHD children were medication naive. In the other three ADHD children, medication had been withheld for 4 weeks (N=1) or 36 hours (N=2). Exclusion criteria were 1) full-scale IQ below 85 (estimated from performance on Vocabulary and Block Design subtests of the WAIS-III [ADHD subjects: mean=104.3, SD=16; comparison subjects: mean=128.4, SD=16.7]); 2) history of neurological or affective disorder; or 3) language disorder (subtest scores below 85 on the Wechsler Individual Achievement Test or Woodcock Mastery Scale). Inclusion criteria for the ADHD group were 1) DSM-IV criteria met as determined by the Diagnostic Interview Schedule for Children
(19) (symptoms: mean=16.3, SD=1.7); 2) parent and teacher endorsements (i.e., ratings of “pretty much” or “very much”) on five out of nine items from the Swanson, Nolan, and Pelham (SNAP) Checklist; and 3) Child Behavior Checklist and Teacher Report Form
(20) Attention Problem scores >60, a cutoff validated by Chen et al.
(21) (ADHD: parent mean=71.3 [SD=7.1]; teacher mean=67.1 [SD=10.7]; comparison subjects: parent mean=50.2 [SD=0.48]). ADHD children with symptoms of conduct disorder (mean=0.78, SD=1.6) and oppositional defiant disorder (mean=4.9, SD=3.0) per the Diagnostic Interview Schedule for Children were not excluded. Among the 10 ADHD children, there was at least one symptom of conduct disorder reported in two children and at least one symptom of oppositional defiant disorder in seven children.
Task Procedure
Stimuli were generated in Psyscope
(22) and viewed via a magnet-compatible projector. Responses were recorded via right-hand held optical buttons. Head movement was minimized using a bite-bar formed with each subject’s dental impression.
Subjects performed two runs of a modified Eriksen flanker task (see reference
11 for details). Each trial consisted of a display comprising a central arrow and two flanking stimuli on either side. Subjects were instructed to press one of two buttons indicating the direction of the central arrow (left/right) while ignoring the flanking stimuli. On congruent and incongruent trials the flankers were arrows pointing in the same or different direction, respectively, relative to the central arrow. On neutral trials the flankers were diamonds that were not associated with a response. On no-go trials, the flanking stimuli were Xs that signaled that subjects should withhold their response.
Trials were presented in a rapid event-related design with a 3-second intertrial interval. On each trial, the display (800 msec) was followed by a blank screen (300 msec) and then a cross hair (1600 msec); intertrial interval was 300 msec. On fixation trials, the cross hair was presented for 2700 msec. The trial sequence followed a stochastic design in SPM 99, in which the probability of each trial type varied sinusoidally between 0 and 1 over a 30-second period. Each trial type had a probability function with a different phase; over time, all trial types occurred with equal probability. Across the two runs, there were 46–58 trials of each type (across subjects 51–52 trials per type) in addition to 44 fixation trials. The trial sequence was specified by one set of lists for half the subjects and another set for the remaining half; the order of the two runs was counterbalanced across subjects.
Imaging Procedure
A 3-T MRI (GE 8.2.5, GE Medical Systems, Milwaukee) was used to acquire T1-weighted flow compensated spin-echo anatomy images (TR=500 msec; minimum TE) in 16 contiguous 7-mm axial slices, parallel to the plane of the anterior commissure-posterior commissure. Functional acquisition included the same slices with a T2*-sensitive gradient echo spiral pulse sequence with parameters of TR=1000 msec, TE=30 msec, field of view=24 cm, flip angle=60°, 64×64 matrix.
Data Analysis
Using SPM 99 (Wellcome Department of Cognitive Neurology, London), images were corrected for differences in slice acquisition time and motion. All subjects displayed less than 1 mm of motion in x, y, and z directions throughout the course of each scan. Estimated motion parameters did not differ between ADHD children (mean=0.139 mm, SD=0.008) and comparison subjects (mean=0.159 mm, SD=0.005) (t=0.55, df=18, p=0.59). Images were normalized into a standard space
(23) and interpolated to 2×2×2 mm cubic voxels. Normalized image volumes were spatially smoothed (6-mm full width at half-maximum Gaussian kernel) and temporally filtered (low-pass filter: 4-msec Gaussian; high-pass filter: calculated on the basis of trial frequency). fMRI responses were modeled by canonical hemodynamic response function and its temporal derivative for correct trials only. For each subject, activation maps were generated using linear contrasts identifying regions that were more active during incongruent relative to neutral trials and during no-go relative to neutral trials.
All analyses were generated by using a voxel-level height threshold of p<0.001 uncorrected for multiple comparisons and spatial extent threshold of 5 voxels; in the event that no activation reached threshold, successively liberal height thresholds, with a more stringent extent threshold, were employed to identify regions of subthreshold activation (e.g., uncorrected p<0.005 or p<0.01 with extent threshold of 10 voxels). First, to identify brain regions most consistently engaged during interference suppression (incongruent > neutral) and response inhibition (no go > neutral) within a group, individual activation maps for each contrast were averaged across subjects separately for the ADHD and comparison groups in a random effects model. Second, group differences were examined with group-by-condition repeated-measures analysis of variance (ANOVA) on signal intensity in functional regions of interest defined by the group average of the comparison group. Third, to identify regions in which the magnitude of activation was correlated with performance, simple regression analyses were performed separately for each group for 1) activation during interference suppression (incongruent > neutral) as a function of subjects’ ability to suppress interference (measured by the accuracy or reaction time difference between neutral and incongruent trials) and 2) activation during response inhibition (no go > neutral) as a function of subjects’ ability to withhold inappropriate responses (measured by the accuracy difference between no go and neutral trials). In the event that similar regions were identified across groups, regression analyses were also performed including all subjects.
Results
Performance
Incorrect or absent responses on congruent, incongruent, and neutral trials and failure to withhold responses on no go trials were classified as errors. Subjects’ mean accuracy rates and response latencies were statistically analyzed separately in group (ADHD, comparison) by trial type (incongruent, neutral) repeated-measures ANOVAs including conditions that were relevant to the fMRI analysis. Overall accuracy was lower in ADHD than in comparison children (main effect of group: F=10.13, df=1, 18, p<0.01). Further, accuracy was lower for incongruent relative to neutral trials (main effect of trial type: F=9.44, df=1, 18, p<0.01). However, this result differed between ADHD and comparison children (group-by-trial type interaction: F=4.89, df=1, 18, p=0.04). Planned contrasts indicated that accuracy was significantly lower for incongruent relative to neutral trials in ADHD children (incongruent: mean=87.4%, SD=10.5; neutral: mean=92.3%, SD=6.7) (t=2.88, df=9, p=0.02) but not in the comparison subjects (incongruent: mean=98.1%, SD=2.1; neutral: mean=98.9%, SD=2.0) (t=1.07, df=9, p=0.31). Thus, comparison children were more successful at suppressing interference relative to ADHD children. Response latencies of correct trials did not differ between groups. Response latencies were longer during incongruent relative to neutral trials (main effect of trial type: F=23.62, df=1, 18, p<0.01), and this finding did not differ between groups, indicating that both groups were susceptible to interference. The mean magnitude of interference (incongruent relative to neutral trials) was 49.2 msec (SD=52.8) in ADHD children and 43.3 msec (SD=28.9) in comparison children. Thus, while failures of interference suppression were more frequent in ADHD children, temporal characteristics of successful interference suppression were similar in the two groups. For response inhibition, accuracy on no go trials was numerically higher for the comparison children (mean=92.2%, SD=4.0) than for ADHD children (mean=78.1%, SD=24.0). This effect was marginally significant (t=1.83, df=18, p=0.08) because of the large variability in performance within the ADHD group.
Imaging
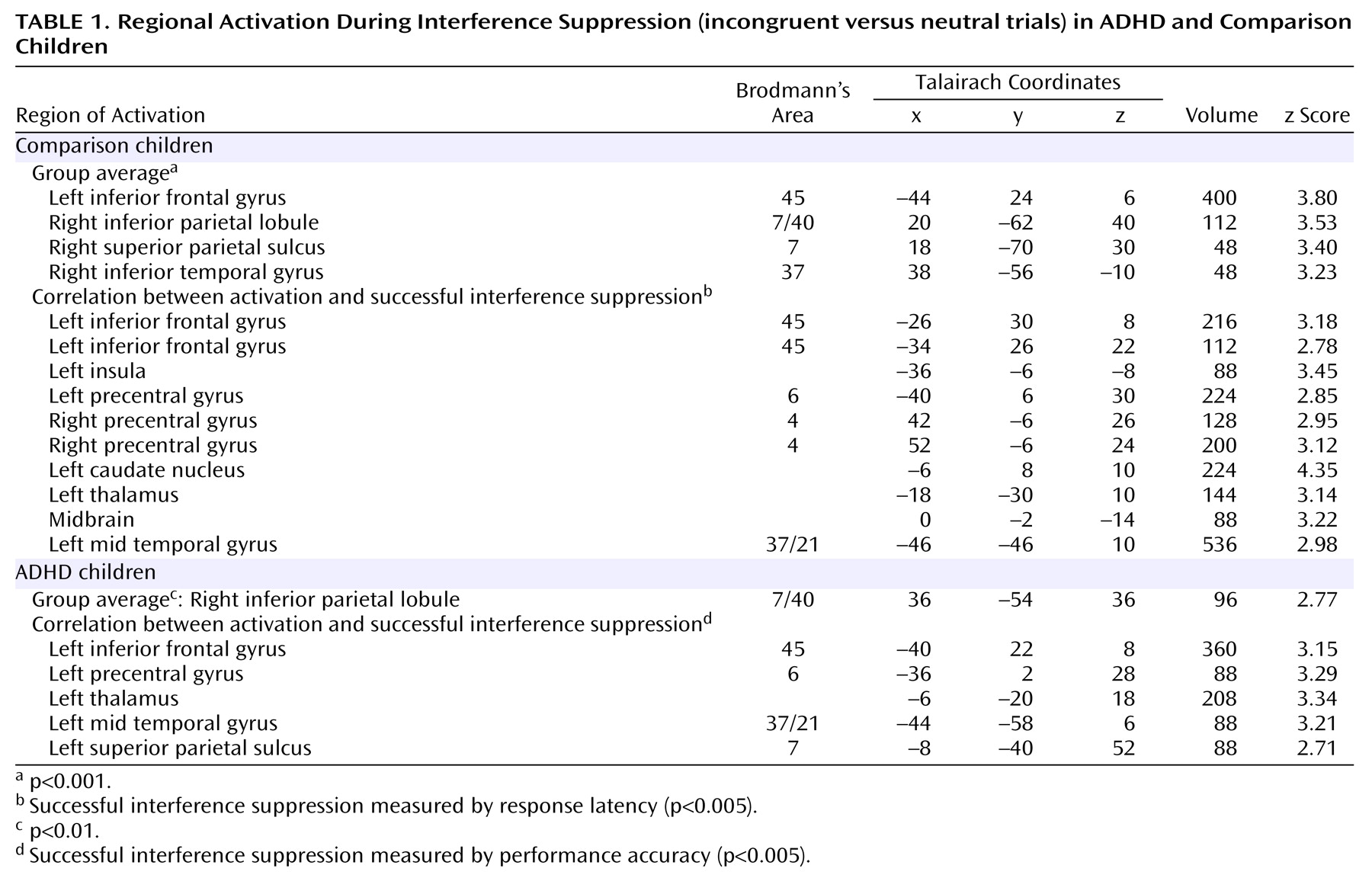
Interference suppression
As seen in
Table 1 and
Figure 1, comparison children activated the left inferior frontal gyrus (Brodmann’s area 45) and adjoining insula and right inferior parietal lobule near the supramarginal gyrus (Brodmann’s area 7/40). Smaller activations were observed in the right superior parietal cortex (Brodmann’s area 7) and inferior temporal cortex (Brodmann’s area 37). ADHD children did not activate any region at a significance threshold of p<0.001 or 0.005. A right inferior parietal region (Brodmann’s area 40), anterior and lateral to that in comparison subjects, was weakly activated in ADHD children. For the left inferior frontal region of interest, a group-by-condition interaction (F=4.44, df=1, 18, p=0.05) was obtained, indicating significant activation during incongruent relative to neutral trials in comparison children (mean=0.049) (t=5.6, df=9, p<0.001) but not ADHD children (mean=–0.019) (t=0.61, df=9, p=0.56). For the right inferior parietal region of interest, the group-by-condition interaction was not significant, but significant activation during incongruent relative to neutral trials was observed in the comparison children (mean=0.059) (t=3.1, df=9, p=0.01) but not the ADHD children (mean=0.036) (t= 0.81, df=9, p=0.44).
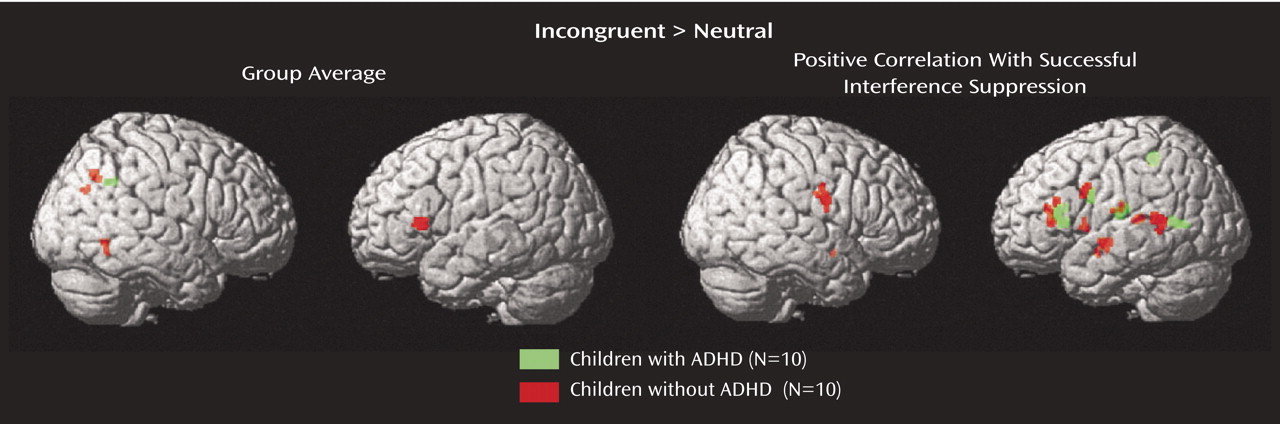
Regression analyses identified regions for which magnitude of activation correlated negatively with magnitude of interference (defined as reduced performance on incongruent relative to neutral trials) for comparison and ADHD children, separately (
Table 1,
Figure 1) and together (
Figure 2). In light of group differences in performance, optimal indices of interference differed between groups. Accuracy was at ceiling in comparison children; therefore, increased response latency during incongruent relative to neutral trials was a better index than accuracy of the magnitude of interference. In contrast, accuracy was significantly reduced in incongruent relative to neutral trials in ADHD children; therefore, accuracy but not response latency best characterized the magnitude of interference. Therefore, regression analyses were performed with latency measures for comparison children and with accuracy measures for ADHD children. Collectively, these analyses revealed that better interference suppression was associated with greater activation in a common network of distributed regions in the left hemisphere in the two groups. Specifically, regions associated with effectiveness of interference suppression were in the inferior frontal gyrus (Brodmann’s area 45) extending into the insula, premotor cortex (Brodmann’s area 4, 6), middle temporal cortex (Brodmann’s area 21), and the thalamus. Better interference suppression in ADHD children was also related to greater recruitment of the medial bank of the superior parietal sulcus (immediately posterior to the postcentral gyrus). Finally, better interference suppression in comparison children was also related to recruitment of the right premotor cortex (Brodmann’s area 4), left caudate, and midbrain. In light of the substantial overlap in brain regions that were positively correlated with successful interference suppression between the two groups, regression analyses with latency and accuracy measures of interference suppression were computed including all subjects. These analyses identified a region in the left inferior frontal gyrus that included the insula and the thalamus (
Figure 2).
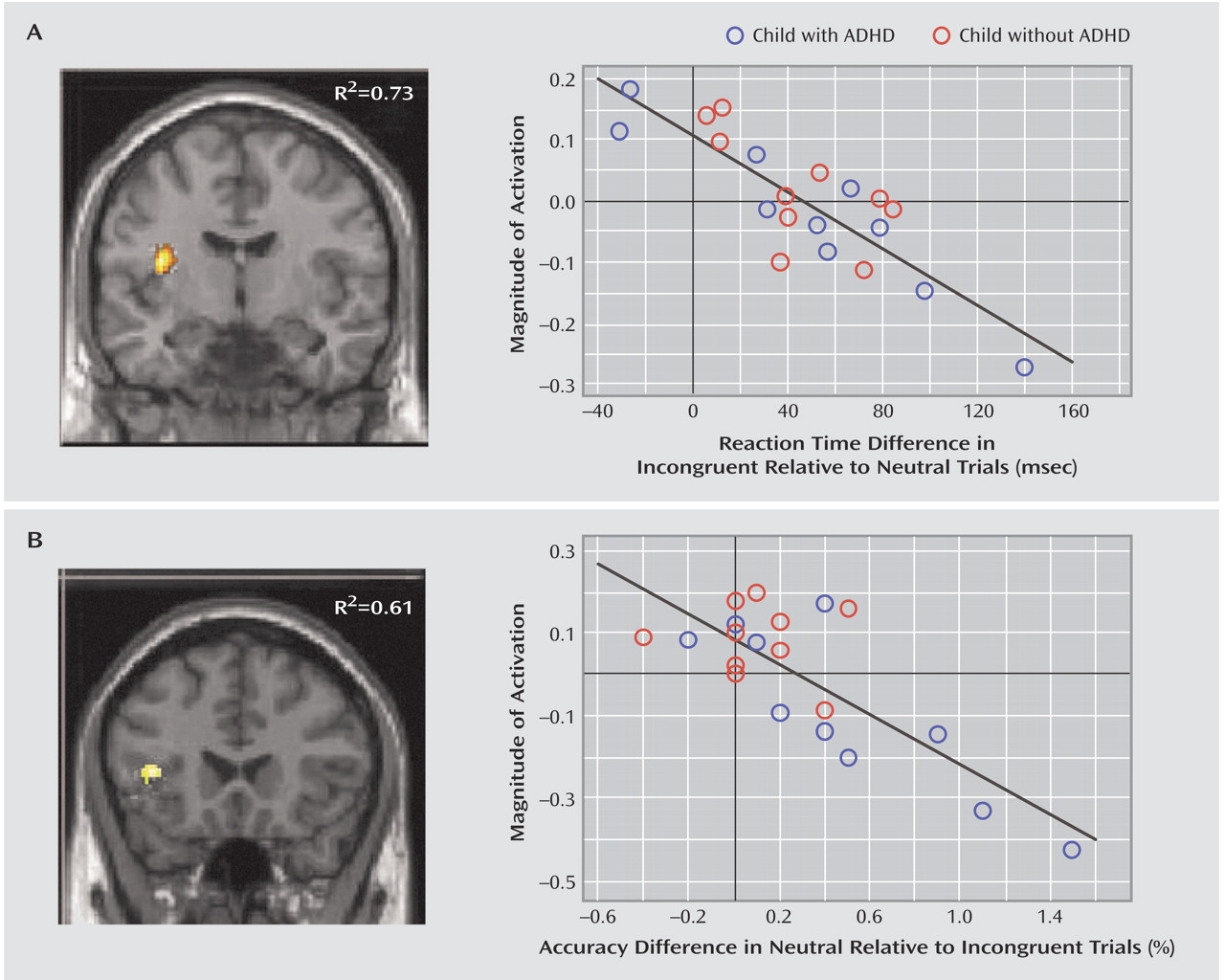
Response inhibition
Comparison children had significant activation in a dorsal region on the border of the right precentral and inferior frontal gyri (Brodmann’s area 6/44) (
Table 2,
Figure 3). For this region of interest, the group-by-condition interaction did not reach statistical significance, but activation was significantly greater during no go relative to neutral trials in the comparison children (mean=0.06) (t=6.2, df=9, p<0.001) but not the ADHD children (mean=0.05) (t=1.0, df=9, p=0.33). No regions were activated at p<0.001 in the ADHD group. A right frontal region in the insula was activated in ADHD children at p<0.005.
Regression analyses identified regions for which magnitude of activation correlated significantly with magnitude of response inhibition (defined as reduced accuracy on no go relative to neutral trials) separately for healthy and ADHD children (
Table 2,
Figure 3). This analysis revealed that better response inhibition was associated with greater activation in the right hemisphere but in different locations in the two groups. Superior response inhibition was associated with activation in the right premotor cortex (Brodmann’s area 8) and bilateral caudate in comparison children, whereas it was associated with the right posterior superior temporal gyrus (Brodmann’s area 22) in children with ADHD. Correlational analyses at a lenient threshold (p<0.005) revealed further differences in the two groups: posterior foci in bilateral superior parietal cortex (Brodmann’s area 7) in comparison children and in the left medial frontal gyrus (Brodmann’s area 8) and postcentral gyrus (Brodmann’s area 43) in ADHD children.
ADHD Symptoms
We examined whether regions identified by group averaging and the regression analyses correlated with ADHD symptom severity as indexed by scores on the attention problems subscale of the Child Behavior Checklist. Magnitude of activation in the regions of interest identified by group averaging in the left inferior frontal and right inferior parietal region for interference suppression and right precentral region for response inhibition did not correlate significantly with symptom severity. Similarly, none of the regions that correlated with successful interference suppression and response inhibition (identified by regression analyses) correlated significantly with symptom severity. These analyses suggest that variability in symptoms, as indexed by a common diagnostic instrument, are limited in sensitivity to cognitive control task performance and associated activation. However, our ADHD group was selected to be highly homogeneous (all combined subtype) and thus may have lacked the range of variability necessary to reveal correlational relationships.
Group Differences in IQ
In light of differences in average IQ between the ADHD and comparison groups, we examined whether a subset of the comparison children (N=5, mean=115) matched in IQ to the ADHD group showed significant activation in the regions of interest identified by group averaging (using two-tailed t tests). For interference suppression, magnitude of activation during incongruent relative to neutral trials was significantly greater in the left inferior frontal region (t=5.4, df=4, p=0.005) but marginally so in the right inferior parietal region (t=1.7, df=4, p=0.15). For response inhibition, magnitude of activation during no go relative to neutral trials was significantly greater in the right precentral region (t=3.6, df=4, p=0.02). Thus, despite a lower IQ, the subset of healthy children showed robust activation within frontal regions. It is unlikely, therefore, that reduced activation in these regions in the ADHD group was related to their lower IQ.
Discussion
Rapid event-related fMRI revealed that two cognitive control operations, the inhibition of inappropriate responses (response inhibition) and the suppression of interfering responses (interference suppression), were associated with different patterns of atypical brain activation in a single group of preadolescent ADHD children. ADHD children failed to suppress interference more often and exhibited reduced engagement of a frontal-striatal-temporal-parietal network that subserved healthy performance. ADHD children showed reduced response inhibition relative to comparison children, and the two groups recruited different anterior brain regions (frontal cortex and caudate nucleus in the comparison subjects but not those with ADHD) and posterior brain regions (superior temporal cortex in ADHD subjects but not comparison subjects). It is unlikely that the observed altered neural basis of cognitive control in ADHD resulted from stimulant exposure, since most children had not been chronically medicated.
Functional activation associated with interference suppression and response inhibition, however, did exhibit common atypicalities in ADHD, namely failure to activate the frontal cortex and caudate nucleus in an age-appropriate manner. The present findings are the first to reveal atypical frontal-striatal involvement during two control operations in the same ADHD children. Atypical activation was not associated with symptom severity of attentional problems. Magnitude and extent of left inferior frontal (Brodmann’s area 45) activation and premotor (Brodmann’s area 6/4) activation associated with interference suppression was reduced in ADHD subjects relative to comparison children. Right frontal (Brodmann’s area 6/44, 8/9) activation associated with healthy response inhibition was not observed in ADHD children; instead, they weakly activated the right insula. Atypical frontal engagement in ADHD is consistent with prior findings regarding a variety of control operations, including response inhibition
(2,
16,
17), working memory
(24), and interference control
(15). Further, caudate activation was associated with both cognitive control operations in healthy children but not in those with ADHD, even at lenient thresholds. Caudate activation has been associated with sensitivity to changes in the probability of events
(25,
26), a property of the present task that varied the frequency of inhibitory events. Atypical caudate engagement across a variety of cognitive control operations in ADHD
(2,
16,
17) suggests deficient encoding of event probability, perhaps leading to impoverished response preparation, as a core deficit of the disorder. Reduced caudate engagement is consistent with findings of reduced caudate volume in ADHD
(13). Convergent evidence, therefore, supports that frontal-striatal abnormalities underlie ineffective cognitive control in ADHD.
Interference Suppression
Despite differences in the magnitude and extent of activation, loci of frontal-temporal activation during interference suppression were similar between groups. Studies of adult interference control suggest that the lateral prefrontal cortex mediates selection from competing responses
(26,
27). Superior interference suppression was associated with greater left inferior frontal engagement in both groups. Lateralization of the activation to the left hemisphere is consistent with verbal mediation of interference suppression. Indeed, fluid verbal ability was negatively correlated with magnitude of interference in healthy children
(11). Verbal mediation is further suggested by activation of other language-related regions such as the left middle temporal cortex in both groups. The extent of frontal-temporal activation, however, was reduced in ADHD children, which may relate to reduced gray matter in those regions
(28). Greater recruitment of the premotor cortex, a region known to be involved in response planning, was associated with better interference suppression in both groups, but premotor activation was more extensive in comparison children than in those with ADHD. Thus, similar frontal and temporal activation loci suggest that both groups engaged similar response selection and planning processes that contributed to interference suppression but that ADHD children engaged these areas to a lesser extent.
In contrast to group similarities in the location (but not magnitude) of activation in frontal and temporal cortices, the two groups differed in terms of parietal activation. Studies of adult interference control suggest that the parietal cortex represents visuospatial maintenance and attentional processes
(26,
27). Right parietal activation was robust in the comparison children but weak in those with ADHD, suggesting reduced visuospatial attentional processing. Better interference suppression in ADHD children, however, was associated with recruitment of the medial bank of the left superior parietal sulcus. This region, extending posteriorly, has been associated with visuomotor attention
(29). Greater recruitment of that region in ADHD children with better interference control may reflect a compensatory strategy that relied upon greater visuomotor attention in the face of reduced visuospatial attention.
Response Inhibition
The functional anatomy of response inhibition differed between the ADHD and comparison children. Response inhibition in the healthy children was related to premotor activation in the right hemisphere. ADHD children revealed frontal activation—only with lenient thresholds—in the right insula and supplementary motor cortex, regions that were not observed in the comparison children with lenient thresholds. Better response inhibition in ADHD children was associated with robust activation of the right posterior superior temporal cortex. This region has been associated with awareness of one’s own motor actions
(30) and may reflect alternative, perhaps compensatory, processes in ADHD children. Those alternative processes were not evoked by the healthy children because temporal activation was not observed even at a lenient threshold. In other studies, regions that were more activated in ADHD than in comparison subjects were in the right inferior parietal cortex
(16,
17). The loci of the putative compensatory activation across studies, right inferior parietal and superior temporal regions, correspond closely to regions exhibiting increased gray matter in ADHD relative to comparison children
(28). The temporal rather than parietal locus of the posterior activation in the present study may reflect processes specific to current task demands. Our task was novel because it interleaved response inhibition (no go) with a variety of response selection trials (neutral, congruent, and incongruent). The variety of go conditions may have reduced the potency of the go response, yielding a marginal group difference in no go accuracy. Prior ADHD studies have reported significant group differences in performance and also more widespread frontal activation than that in our study
(2,
16,
17). Those studies did not employ designs optimized to visualize selectively successful performance, as done in our study. Thus, design, task, and performance characteristics may account for differences across studies.
Conclusions
Our findings refine and extend current understanding of cognitive control in ADHD in four ways. First, our findings provide the first evidence for multiple, rather than unitary, patterns of functional neural abnormality in ADHD. Greater overlap between ADHD and comparison children in regions mediating interference suppression than response inhibition suggests differential sensitivity to ADHD. Indeed, methylphenidate, a stimulant effective for alleviating ADHD symptoms, had selective effects on cognitive control: ADHD children exhibited improved response inhibition but not interference suppression
(4). Methylphenidate enhances catecholaminergic systems innervating frontal-striatal regions. Enhanced frontal-striatal function may be insufficient to improve interference suppression because parietal-temporal contributions are necessary
(26,
27). However, posterior activations during response inhibition vary across studies
(16,
17, present study) and thus may reflect participatory but unnecessary processes. Second, the differences in activation between ADHD and comparison children could not have resulted from prolonged medication exposure, since most of the ADHD children were medication naive. Third, findings of past ADHD studies have been ambiguous regarding the extent to which functionally atypical regions reflect lower task performance or disorder. These factors were disambiguated by our study because we identified brain regions that were typically and atypically related to intragroup variability. Fourth, our results strengthen the view that the caudate nucleus is a region of functional abnormality related to ADHD rather than a specific task because it was abnormal across two cognitive control operations within the same preadolescent ADHD children.