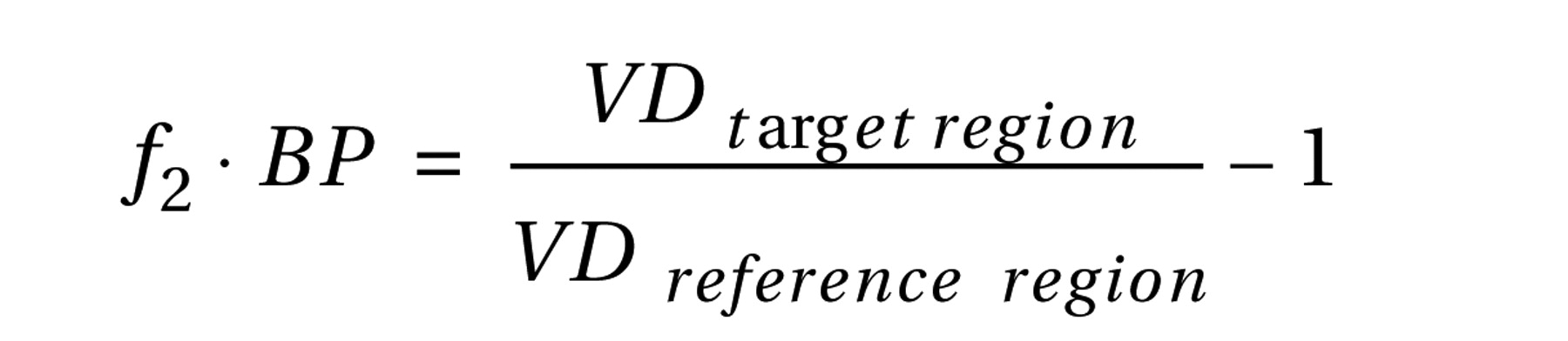For more than three decades, there has been intense interest in the pathophysiological role of the serotonin transporter (5-HTT) in depressive disorders. The methods available to study 5-HTT in humans have steadily become more sophisticated, and with the use of selective radioligands, it is now possible to image 5-HTT directly in the living human brain.
In vivo imaging studies of 5-HTT have generally been conducted in patients suffering from acute depression, and the results have been somewhat inconsistent, perhaps in part because of the differing methodologies used. Earlier studies employed [
123 I]-bCIT together with single photon emission computed tomography (SPECT). On balance, depressed patients appear to have decreased binding potential of 5-HTT in the midbrain and brain stem regions
(1 –
4) ; however, one study found increased 5-HTT binding potential in both regions in depressed adolescents
(5) . Of the studies using the newer SPECT agent [
123 I]-ADAM, one showed decreased binding potential in the midbrain region in depressed patients compared with healthy volunteers
(6), but another study found no difference
(7) .
A limitation of studies that utilize [
123 I]-bCIT is that it has similar affinity for the dopamine transporter and 5-HTT
(8) and it does not permit imaging of 5-HTT in certain brain areas relevant to depression, such as the anterior cingulate cortex and limbic structures. However, two more recently developed positron emitting ligands, [
11 C]McN5652 and [
11 C]DASB, allow quantification of 5-HTT in these brain regions outside the brain stem
(9,
10) . Once again, the data in acute depressive states are somewhat contradictory. Using [
11 C]McN5652 with positron emission tomography (PET), Parsey et al. found that unmedicated patients with major depression had lower 5-HTT binding potential in the amygdala and midbrain region than healthy comparison subjects
(11) . The same group reported similar but somewhat larger decreases in 5-HTT availability in patients with acute bipolar depression
(12) . However, two smaller studies employing the same ligand found increased 5-HTT binding in the thalamus and cingulate cortex and no change in the midbrain region in depressed patients
(13,
14) .
In a comparative evaluation of 5-HTT radioligands, [
11 C]DASB compared favorably with [
11 C](+)McN5652, both in nonhuman primates
(15) and humans
(16), with the advantage of possessing higher specific to nonspecific binding. Using [
11 C]DASB and PET to measure 5-HTT, Meyer et al. found no differences in 5-HTT binding potential in unmedicated depressed patients relative to healthy comparison subjects
(17) . However, Cannon et al., also using [
11 C]DASB in depressed bipolar patients, found increased binding potential in the thalamus, cingulate cortex, and insula. In contrast, binding potential in the pons region was decreased; it did not differ from that of healthy comparison subjects in the midbrain region
(18) . Using PET in conjunction with [
11 C]Harmine, Meyer et al. recently found widespread increases in monoamine oxidase (MAO)-A density in unmedicated depressed patients
(19) . They suggested that increased MAO-A and the resulting decrease in monoamine levels might be the primary biochemical lesion in depressed patients. They also suggested that patients with normal or increased density of 5-HTT would be more at risk for depression in the presence of elevated MAO because synaptic serotonin (5-HT) levels would be lowered even further by an effective 5-HT reuptake process. Clearly, this idea contrasts with the notion that patients with recurrent depression may have long-standing deficits in 5-HTT availability
(11,
12) .
We have used radioligand imaging to study 5-HT-related receptors in recovered depressed patients withdrawn from medication. The aim is to identify relatively enduring abnormalities in brain 5-HT mechanisms in people at risk for recurrent illness. An advantage of this approach is that any changes seen are unlikely to be attributable to epiphenomena of acute depressive illness or its treatment. Thus far, we have found that recovered depressed patients exhibit diminished cortical binding potential of 5-HT
1A receptors measured by [
11 C]-WAY 100635
(20), while cortical 5-HT
2A receptor binding potential, measured by [
11 C]-MDL 100907, is increased
(21) . The purpose of the present study was to use [
11 C]DASB with PET imaging to measure 5-HTT binding potential in recovered depressed patients. Low levels of 5-HTT binding in recovered subjects would suggest a persistent deficit in 5-HTT and perhaps in presynaptic 5-HT innervation. In contrast, normal or increased levels of 5-HTT would be consistent with the modified monoamine theory proposed by Meyer et al.
(19) .
Method
Participants
The study was approved by the research ethics committee at Hammersmith Hospital, London, and the Administration of Radioactive Substances Advisory Committee, United Kingdom. All participants gave written informed consent. We recruited 20 healthy men (age: mean=38.9 years [SD=11.9]), with no current or past psychiatric history, and 24 recovered depressed men (age: mean=35.7 years [SD=9.8]) who had experienced at least one episode of unipolar major depression in the past (number of episodes: mean=2.4; range=1–5 [87.5% had >2 episodes]). All patients had been euthymic for a mean duration of 31 months (SD=31) (range=3–108 months), nine had never received any antidepressant medication, and the rest were medication free for 23 months (SD=30) (range=3–120 months). The mean age of onset of a first episode of depression was 24.6 years (SD=9.3) (range=16–52 years), and the overall duration of illness was 11.5 years (SD=7.6) (range=1–30 years). Four patients had attempted suicide in the past during their depressive episodes, nine had melancholic symptoms during at least one of their episodes, and eight had a family history of major depressive disorder.
Subjects were diagnostically assessed by a clinical psychiatrist. All patients met Structured Clinical Interview for DSM-IV (SCID) disorders criteria for major depressive disorder in the past and were fully recovered. Subjects were rated to be recovered from depression based on the following four criteria: self-reported euthymia following the last episode of major depression, clinician-rated euthymia, absence of criteria for a major depressive episode as determined by SCID, and a Hamilton Depression Rating Scale (HAM-D) score of less than 7
(22) . Only subjects who met all four criteria for recovery in addition to being euthymic and medication free for more than 3 months were included in the study and subsequently scanned. Subjects with any serious medical or neurological (current or past) illness, alcohol or illicit substance dependence, or other axis I or II disorders were excluded. All subjects were administered HAM-D and Beck Depression Inventory. They also completed the Dysfunctional Attitude Scale because a previous study showed that Dysfunctional Attitude Scale scores correlated positively with regional 5-HTT binding potential in acutely depressed patients
(17) . We included only male subjects in this study because of a lack of toxicology data for [
11 C]DASB in women.
PET Scanning
PET scans were carried out at the Cyclotron unit, Hammersmith Hospital, London, on an ECAT EXACT three-dimensional scanner (CTI, Knoxville, Tenn.), with an axial field of view of 23.4 cm and 95 reconstructed transaxial image planes
(23) . A 5-minute transmission scan using a [
137 ]Cs point source was carried out prior to each study for subsequent attenuation correction and scatter correction. The 90-minute three-dimensional dynamic emission scan was acquired in list mode. In the postacquisition frame rebinning, 28 time frames of increasing length were generated (30-second background frame prior to the injection, then 1×15-second-frame, 1×5-second frame, 1×10-second frame, 3×30-second frames, 3×60-second frames, 3×120-second frames, 3×180-second frames, 8×300-second frames, and 4×450-second frames). The spatial resolution of the images reconstructed using the reprojection algorithm with the ramp and Colsher filters set to Nyquist frequency is close to isotropic: 5.1 mm full width at half maximum transaxially and 5.9 mm full width at half maximum axially averaged over a radius of 10 cm from the center of the field of view
(23) .
Radiotracer
The radiotracer [
11 C]DASB was synthesized as previously described
(24) . The standard DASB and the precursor desmethyl DASB were obtained from Target Molecules Ltd. (Southampton, United Kingdom). [
11 C]DASB was injected into an antecubital vein as a smooth bolus over 30 seconds. The injected radioactivity dose was between 366 MBq and 566.54 MBq (mean=540.11 MBq [SD=31]). The radiochemical purity of the injected [
11 C]DASB was high and ranged from 95%–99% (mean=97% [SD=1]). The injected mass of cold DASB varied between 0.69 and 10.5 μg (mean=3.3 mg [SD=1.9]). The specific activity was on average 61,021 MBq·μmol
–1 (SD=34,608).
Magnetic Resonance (MR) Scans and Definition of Volumes of Interest
All volunteers had a structural T1 magnetic resonance imaging (MRI) scan performed on either 0.5T (0.5 Apollo System, Marconi Medical Systems, Cleveland) or 1.5T (1.5 Eclipse System, Marconi Medical Systems, Cleveland) MR scanners (TR=30 seconds, echo time=3 seconds, flip angle=30, number of signal averages=1, voxel dimensions=0.98 mm×1.6 mm ×1.6 mm, acquisition time=13 minutes) or 3T (3T Intera Philips Medical Systems, Zurich, Switzerland) MR scanners (TR=9.6 seconds, echo time=4.6 seconds, flip angle=8, number of signal averages=1, voxel dimensions=0.94 mm×0.94 mm×1.2 mm). The scans were inspected by an independent clinical radiologist and found to be normal.
MRIs were resliced to a voxel size of 1 mm×1 mm×1 mm, centered on the anterior commissure and aligned to the anterior-posterior commissure line. MRIs were coregistered to the individual summated PET images using Statistical Parametric Mapping (SPM)2, which adopts a rigid body transformation using a normalized mutual information method. Eight target regions of interest (amygdala, anterior cingulate cortex, caudate nucleus, frontal cortex, hippocampus, insula, dorsal raphe, and thalamus), chosen on the basis of previous studies, and a reference region, the cerebellum, were defined on the coregistered MRI, with the help of a probabilistic brain atlas template
(25) . Standard Montreal Neurological Institute T1 template (available in SPM2) was normalized to the coregistered, individual MRI, and the deformation parameters were applied to the probabilistic atlas. This normalized brain atlas was resliced to PET space and segmented to obtain data from gray matter only. Cerebellar gray matter was used as a reference region because of the negligible density of 5-HTT in the cerebellum
(26) . The dorsal raphe region was manually defined as a fixed-size region (900 mm
3 ) on the summed PET images of each individual. Dynamic PET scans were sampled by applying the individual region-of-interest object maps.
Input Function
The arterial plasma input function was derived from continuous, online whole-blood monitoring (initial 28 minutes) and 10 discrete blood samples, in eight of which the fraction of unmetabolized parent compound was determined. The time course of the plasma-to-blood ratio, obtained from the first five discrete arterial samples at 3-, 9-, 15-, 21-, and 28-minute scan times, was first fitted to a sigmoidal curve. Then, the measurement of the arterial whole-blood activity obtained from the continuous detector system
(27) was multiplied with that ratio to obtain a total plasma activity curve for the first 28 minutes of the scan. This curve was then combined with the discrete plasma activity concentration measurements at 35, 42, 50, 70, and 90 minutes to an input function describing the total plasma activity concentration for the entire scan. Finally, the input function of the activity concentration, because of unmetabolized [
11 C]DASB in plasma, was created by multiplying the total plasma activity input function with the function obtained from the fit of the model for the parent fraction in plasma to the eight measurements of the parent compound during the scan. Calculations were performed using Matlab (MathWorks, Inc., Natick, Mass.) on Sun Ultra 10 workstations (Sun Microsystems, Inc., Santa Clara, Calif.).
Quantification of DASB Binding
To quantify the binding of [
11 C]DASB in brain tissue, the graphical analysis of reversible radioligand binding using the plasma input function
(28) was applied to obtain regional estimates of the total volume of distribution from the slope of the linear part of the plot. The threshold was set at 35 minutes, i.e., the transformed data from frames 20 to 28 were fitted with a straight line.
Using the definition of binding potential introduced by Mintun et al.
(29), estimates of binding potential in target regions of interest (with specific binding) were obtained from the estimated volumes of distribution in those regions and in the cerebellum (reference region without specific binding)
(30) . The following equation was used to determine the estimates of binding potential:
To also assess [
11 C]DASB binding on a pixel level, spectral analysis was used to generate parametric maps of volume of distribution
(31,
32) . A set of 100 basis function, logarithmically spaced between β
min =0.0007s
–1 and β
max =0.1s
–1, was chosen to obtain unbiased estimates of volume of distribution for the specific kinetics of [
11 C]DASB
(33) . Individual volume of distribution images were then normalized to the standard Montreal Neurological Institute T1 template, and average volume of distribution images for the two cohorts were calculated.
Statistical Analysis
Statistical analyses were performed using Statistical Package for Social Sciences, Version 12 (SPSS Inc., Chicago). Demographic data were analyzed using an independent samples t test. We tested our hypothesis that regional 5HTT receptor binding potential would be reduced in patients recovered from depression using a repeated measure analysis of variance (ANOVA), with “diagnosis” (patients versus comparison subjects) as a between-subjects factor and “region” (brain region of interest) as a within-subjects factor, with Huynh-Feldt correction. Significant interactions or main effects were explored using post hoc, independent, two-tailed t tests. Correlations were analyzed using Pearson’s correlation coefficient, and significance was set at p<0.05.
Results
There was no significant difference between recovered depressed patients and healthy comparison subjects in either Beck Depression Inventory (healthy volunteers: 4.0 [SD=7.8] versus patients: 5.8 [SD=5.0]; t=0.88, df=42, p=0.39) or HAM-D ratings (healthy volunteers: 0.4 [SD=0.7] versus patients: 1.0 [SD=1.5]; t=1.8, df=42, p=0.075). However, patients scored significantly higher on the Dysfunctional Attitude Scale relative to healthy comparison subjects (healthy volunteers: 104 [SD=27] versus patients: 128 [SD=0]; t=2.8, df=42, p=0.008).
There was no significant difference in the specific activity (healthy volunteers: 55,788 MBq/μmol [SD=42,361] versus patients: 65,383 MBq/μmol [SD=26,705]; t=0.9, df=42, p=0.4) or amount of cold ligand injected (healthy volunteers: 3.7 μg [SD=1.8] versus patients: 3.1 μg [SD=2.1]; t=1.1, df=42, p=0.3).
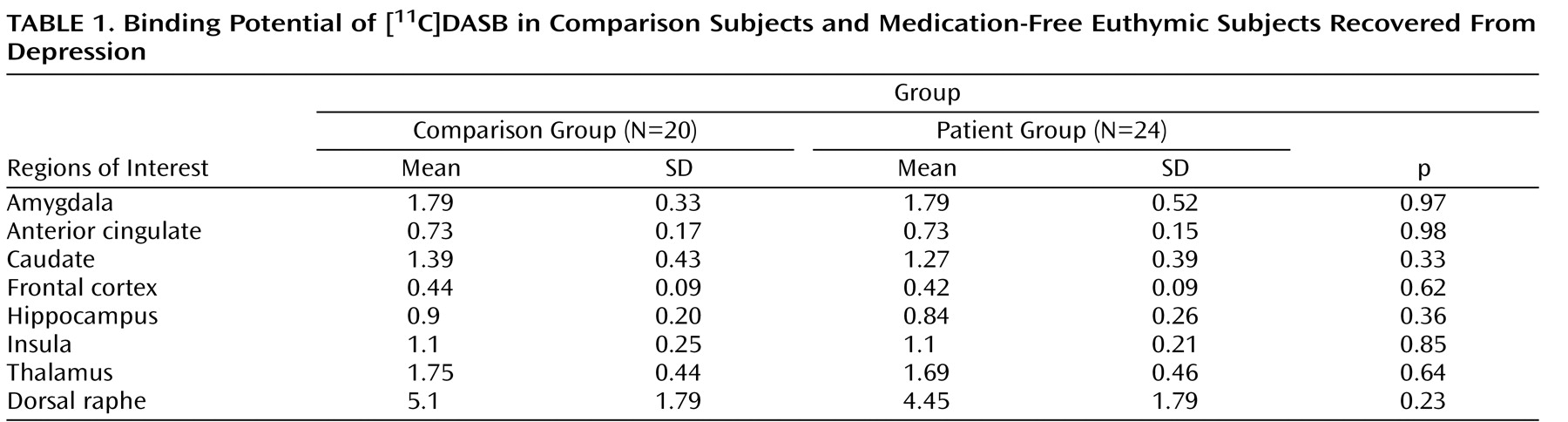
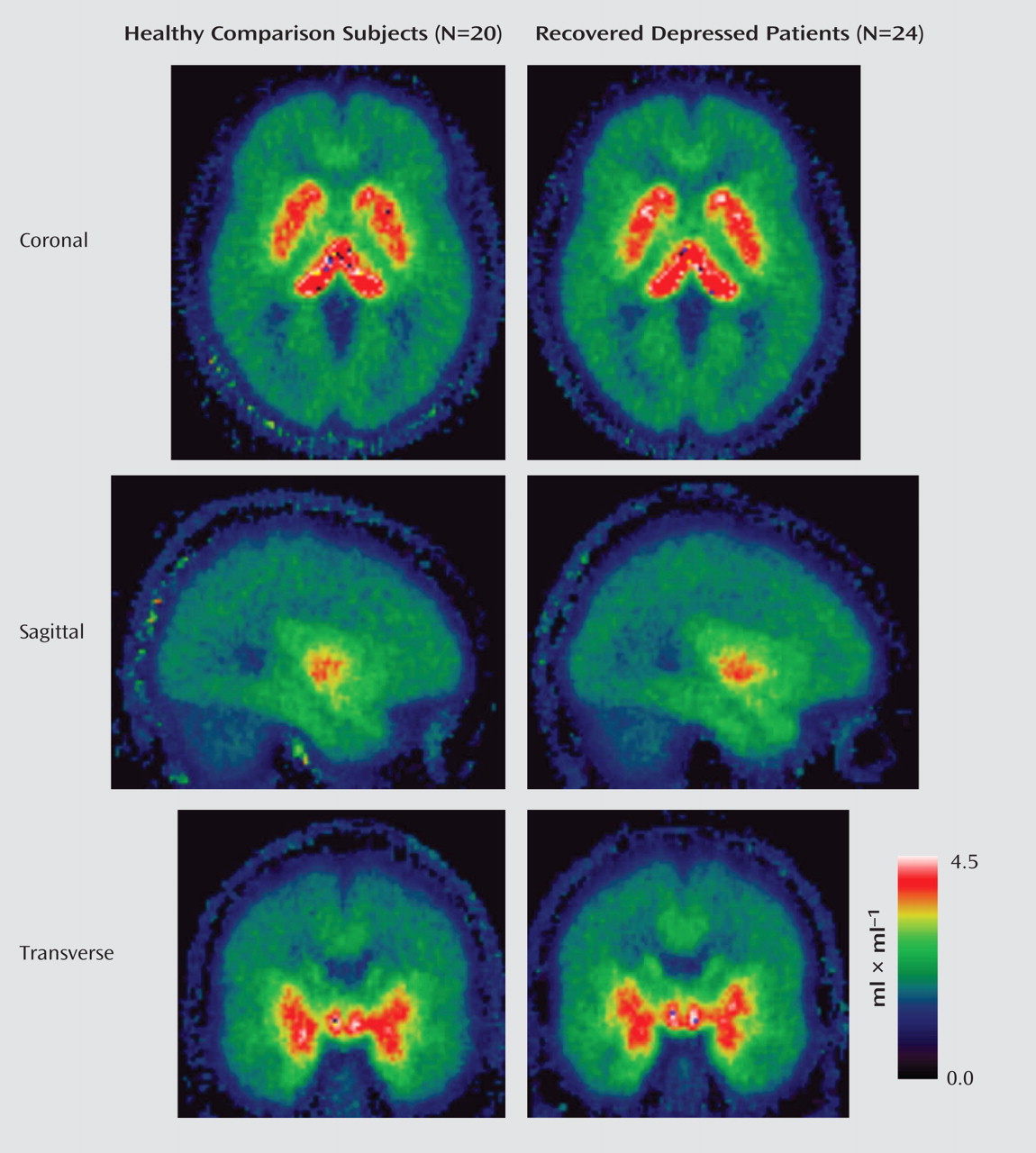
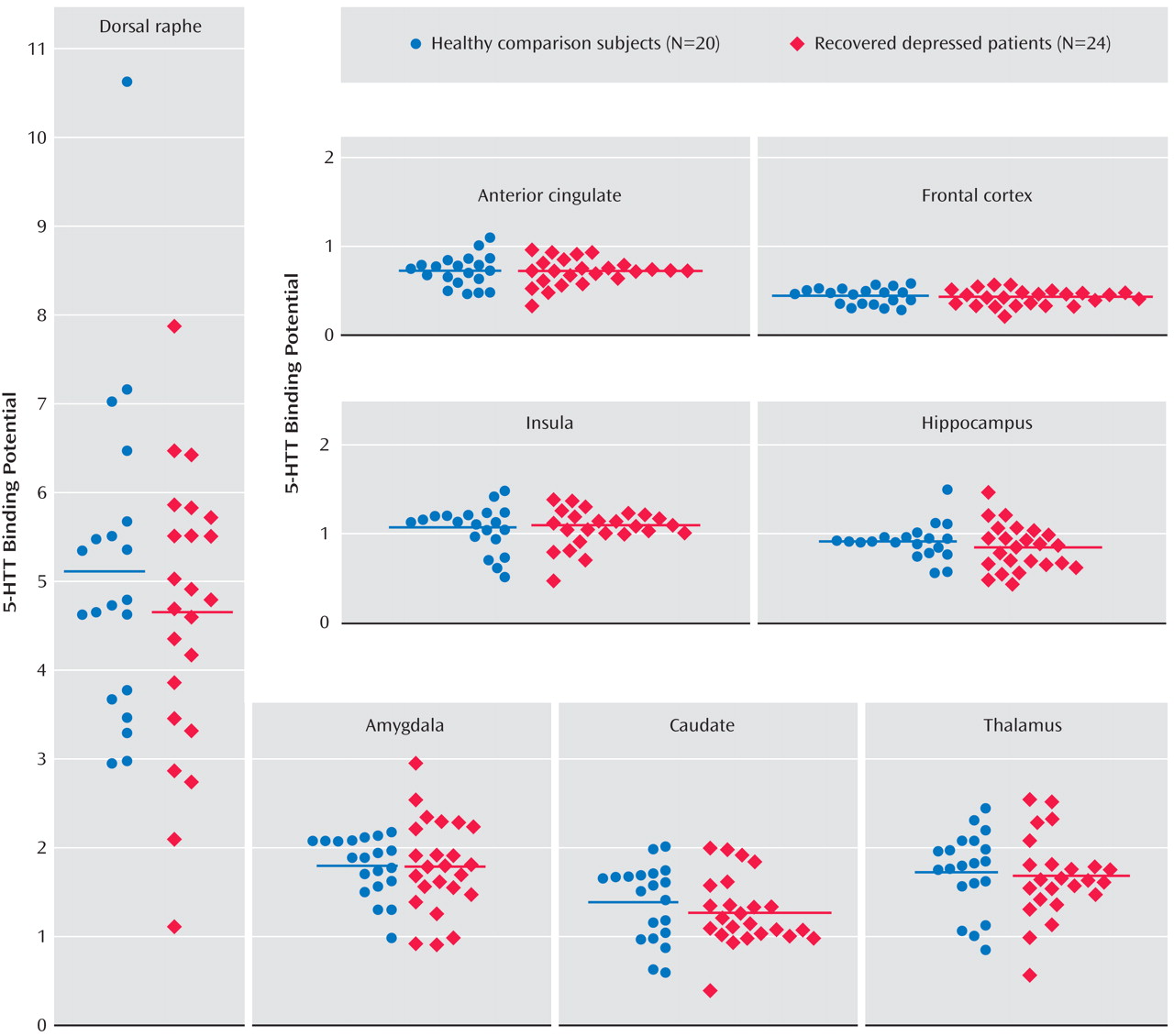
The repeated measures ANOVA of the binding potential data showed a main effect of region (F=178.9, df=1.3, 53.1, p<0.001), with the highest binding density apparent in the dorsal raphe; intermediate levels in the thalamus, anterior cingulate, and hippocampus; and lowest density in the frontal cortex. However, there was no significant main effect of group (F=1.68, df=1, 42, p=0.2) or group-by-region interaction (F=1.23, df=1.3, 53, p=0.28) (
Table 1,
Figure 1,
Figure 2 ). There were also no group differences between healthy comparison subjects and patients with previous episodes of melancholia (F=2.1, df=1, 27, p=0.15), medication-naive patients (F=1.97, df=1, 27, p=0.17), or patients with a positive family history (F=1.2, df=1, 26, p=0.29). Scores on the Dysfunctional Attitude Scale in patients did not correlate with binding potential values in any of the eight regions of interest studied (all p values>0.1). There was also no significant correlation between binding potential in any region and the number of episodes of depression, duration of illness, age of onset of depressive episode, or duration of euthymia in the patient group (all p values>0.1).
Discussion
The results of our study indicate that 5-HTT binding potential is not altered in a number of pathophysiologically relevant brain regions in male recovered depressed patients. This finding argues against the idea that depressed patients have persistent deficits in the availability of 5-HTT. In addition, insofar as 5-HTT provides an index of 5-HT neuronal density
(34), our results suggest that the presynaptic 5-HT innervation of limbic and cortical areas is unchanged in recovered depressed patients. Our data therefore are more consistent with the modified monoamine theory proposed by Meyer et al.
(19) .
As noted earlier, the results of ligand imaging studies of 5-HTT in patients with acute depression have yielded contradictory findings. There are a number of factors that could explain these discrepancies, including psychiatric comorbidity, differences in ligand specificity, previous drug treatment, and effects of acute illness (e.g., weight loss, insomnia). Studying recovered depressed patients avoids some of these potential confounds. In addition, the ligand we used for our study, [
11 C]DASB, binds with high affinity and selectivity to 5-HTT and has a number of advantages over other 5-HTT tracers, including better specific-to-nonspecific binding
(15,
16) .
A previous study by Meyer et al. in patients with acute unipolar depression found no difference in [
11 C]DASB binding potential between depressed patients and healthy comparison subjects
(17) . However, in the latter investigation, there was a positive correlation between self-ratings on the Dysfunctional Attitude Scale and 5-HTT binding potential in the depressed patients. This relationship was not seen in the present study, despite patients showing significantly higher Dysfunctional Attitude Scale scores than healthy volunteers. However, because the patients were clinically recovered, their mean Dysfunctional Attitude Scale scores were lower than those found in the acutely depressed patients in the study conducted by Meyer et al.
(17), in which patients with Dysfunctional Attitude Scale scores >190 had significantly greater DASB binding than comparison subjects. Therefore, it is possible that only patients with particularly high levels of dysfunctional attitudes might demonstrate increased 5-HTT binding. A recent study by Cannon et al.
(18), also using [
11 C]DASB, found several interesting changes in 5-HTT in acutely depressed bipolar patients. While increased 5-HTT binding potential was seen in the cingulate cortex and thalamus, 5-HTT binding was lowered in the brainstem at the level of the pontine raphe. No change was seen in the midbrain region.
In general, studies with [
123 I]-βCIT have indicated a lowering of 5-HTT in the midbrain region and hypothalamus of acutely depressed patients, although there are inconsistencies. The most recent study with [
123 I]-βCIT, conducted by Staley et al.
(3), also found a decrease in 5-HTT availability in diencephalon in unmedicated depressed patients. However, this reduction was limited to female subjects. Because of the lack of availability of suitable toxicity data with [
11 C]DASB, we were unable to include female participants in the present study. Therefore, our negative findings are applicable only to male participants, and it is possible that 5-HTT availability might be persistently decreased in recovered female depressed patients. However, Cannon et al. found no clear effect of gender in their study of [
11 C]DASB in bipolar depressed patients
(18) .
Previous studies in acutely depressed patients have used graphical methods of analysis with a reference-region approach and simplified reference-tissue models. We found no detectable differences in [
11 C]DASB binding potential between recovered depressed patients and healthy volunteers using the following three additional methods of analysis: 1) graphical analysis of reversible binding using a reference-region (cerebellum) input function
(35) ; 2) a reversible two-tissue compartment, four-rate constant compartmental model with free blood-volume term; and 3) spectral analysis (data not shown). The mean parametric maps of volume of distribution obtained with spectral analysis shown in
Figure 1 also illustrate the similarity in binding potential between recovered depressed patients and healthy comparison subjects. In these circumstances, the sensitivity of our analysis to detect changes in binding potential between the two participant groups becomes a critical issue. Power analysis suggests that with this study population we would have been able to detect a change of 15%–20% in the binding potential estimates in most of the brain regions studied at 80% power and p<0.05. This suggests that our study was sufficiently powered to detect differences of the order usually reported between depressed patients and comparison subjects in ligand imaging studies of 5-HTT.
A number of other possible explanations need to be considered for the observed lack of difference between 5-HTT binding potential in recovered depressed patients and comparison subjects. For example, while our patients had been unmedicated for long periods of time, the long-term effects of previous antidepressant treatment in the human brain are not known. Studies in animals have shown that treatment with different classes of antidepressants can produce adaptive changes in 5-HTT
(36,
37), but the duration of these effects after treatment cessation has not been clarified. Conceivably, however, persistent treatment-induced adaptive changes in 5-HTT might have obscured an underlying pathophysiological abnormality in our subjects. However, consideration of treatment-naive subjects alone did not reveal any tendency toward a difference in binding potential.
While our findings appear to exclude an abnormality in 5-HTT availability in recovered depressed patients, there is increasing evidence from animal studies suggesting that altered 5-HT function early in life can have profound effects on neuronal development, and these effects can in turn influence responses to stress and adversity in adulthood
(38) . Hence, normal levels of 5-HTT in adult life do not exclude important effects of altered 5-HTT activity on the development of the neural circuitry supporting emotional regulation. Indeed, our other PET studies in recovered depressed patients have revealed altered availability of cortical postsynaptic 5-HT
1A and 5-HT
2A receptors
(20,
21) . These receptors are often colocalized on cortical neurons where they have opposing physiological roles
(39) . Thus, even in the presence of normal levels of 5-HTT, recovered depressed patients appear to have changes in postsynaptic 5-HT-receptor expression, which could lead to abnormalities in the excitability of cortical networks. The notion that such changes could be developmental in origin is intriguing and will require appropriate studies in high-risk subjects before the onset of depressive disorder.