Disturbances of emotional experience, and in particular a decrease in capacity to experience pleasure (anhedonia), have long been regarded as characteristic features of pathology in schizophrenia (
1), and they have been associated with functional impairment (
2). However, recent research on emotional disturbances in schizophrenia has raised fundamental questions about the nature of these deficits. Schizophrenia patients report experiencing lower levels of pleasure in their daily lives than nonpatients on measures of trait social and physical anhedonia (
3). In contrast, the patients' immediate (or “in the moment”) ratings of experienced emotion are intact, in both laboratory experiments and real life, and reflect the same two-dimensional structure (valence and arousal) as that of healthy subjects (
4).
A possible mechanism underlying the patients' deficits in trait reports of emotional experience can be formulated when one considers a model of cognitive control (
5) in which the dorsolateral prefrontal cortex provides top-down “guided activation” that biases representations toward context-appropriate responding and away from automatic but inappropriate ones. Together with data from affective science (
6), this model points toward integration of emotion and cognitive control when people make decisions based on affective states. Cognitive control processes supported by the dorsolateral prefrontal cortex have indeed been found to modulate the prediction, generation, interpretation, or regulation of emotions (
7) and to translate affectively valenced motivational states into goal-oriented behavior (
8). Thus, reporting on one's experienced emotion outside the presence of an eliciting stimulus, as is often the case with clinical interviews or anhedonia measures, likely relies on cognitive control processes that depend on the dorsolateral prefrontal cortex, such as establishing and maintaining representations that link and integrate affective states with task-relevant responses. Consequently, the patients' deficits as reflected in clinical interviews or anhedonia measures may stem in part from an impaired capacity to build and maintain such representations, which are necessary for generation and maintenance of emotional responses in the face of competing ones. This possibility is consistent with previous behavioral findings (e.g., as in references
9 and
10) and functional magnetic resonance imaging (fMRI) results (such as in references
11–13) indicating that schizophrenia patients exhibit impairments of dorsolateral prefrontal cortex functioning necessary for maintaining task-relevant informa-tion needed to override prepotent responses. It also accords with findings that better working memory performance in patients is associated with a smaller discrepancy between in-the-moment and delayed reports of experienced emotion (
14).
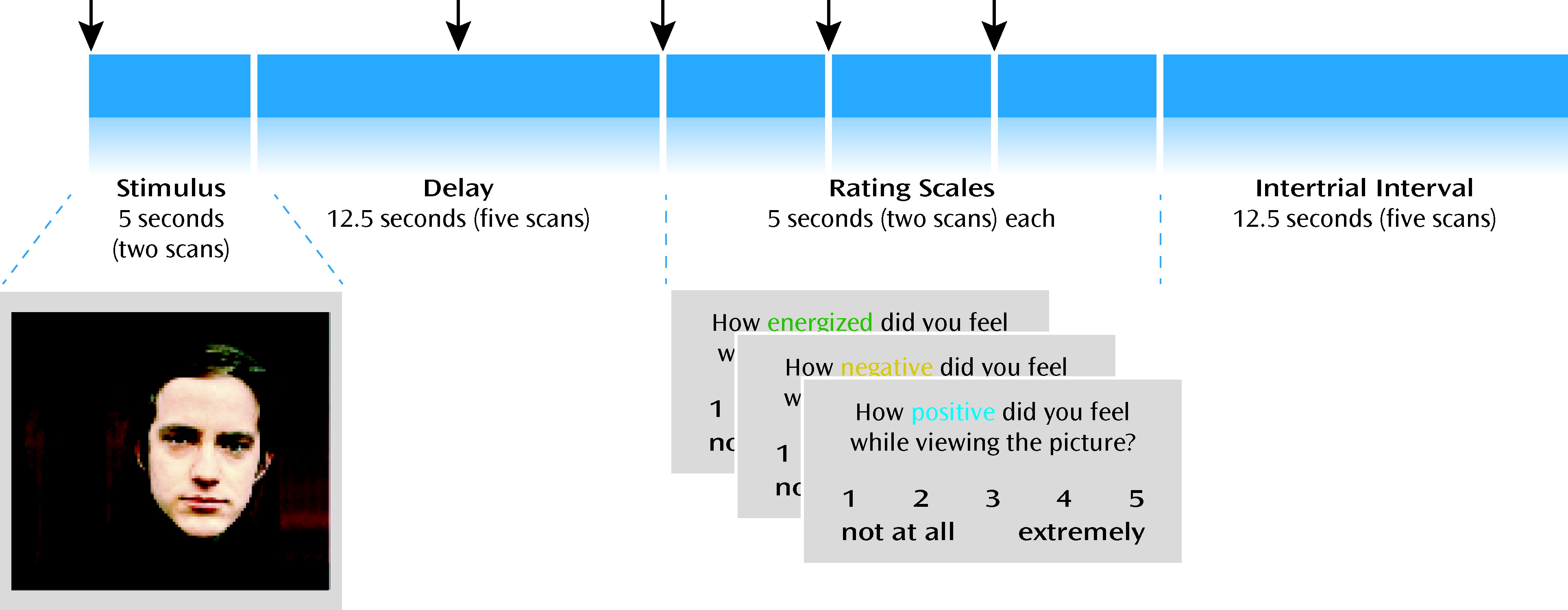
fMRI studies examining brain activation during presentation of emotion-eliciting stimuli in schizophrenia (
15–17) have generated variable findings, including limbic and prefrontal increases and decreases in activity in patients relative to healthy subjects. To our knowledge, no fMRI study has specifically examined the neural correlates of cognitive control processes related to maintaining and reporting on emotional experience in schizophrenia. We used event-related fMRI to explore brain activity during trials in which subjects viewed affective images and, after a delay, rated how “positive,” “negative,” and “energizing” the emotion was while they were viewing the image. Performing this task required controlled processes depending on the dorsolateral prefrontal cortex, such as active maintenance of affective representations and effective manipulation of valenced aspects of the emotional experience in order to accurately rate it on dimensions both consistent and inconsistent with its overall valence. This design allowed us to test the hypothesis that schizophrenia patients' in-the-moment emotional reactivity is spared but that they have a deficit in cognitive control processes needed to maintain and report on emotional experience after the eliciting stimulus is removed. In support of this hypothesis, we sought evidence of 1) robust brain activation during the presence of emotional stimuli, in particular in limbic and paralimbic structures, consistent with an intact ability to generate an emotional response in the presence of an emotional stimulus, and 2) altered delayed ratings of experienced emotion and delayed brain activation, in particular in areas critical for active implementation of cognitive control (e.g., dorsolateral prefrontal cortex). In contrast, the alternative hypothesis (that schizophrenia patients are characterized by a fundamental impairment in emotion generation) would be supported by impaired brain activation even during the presence of the emotional stimuli in structures involved in affective representations.
Discussion
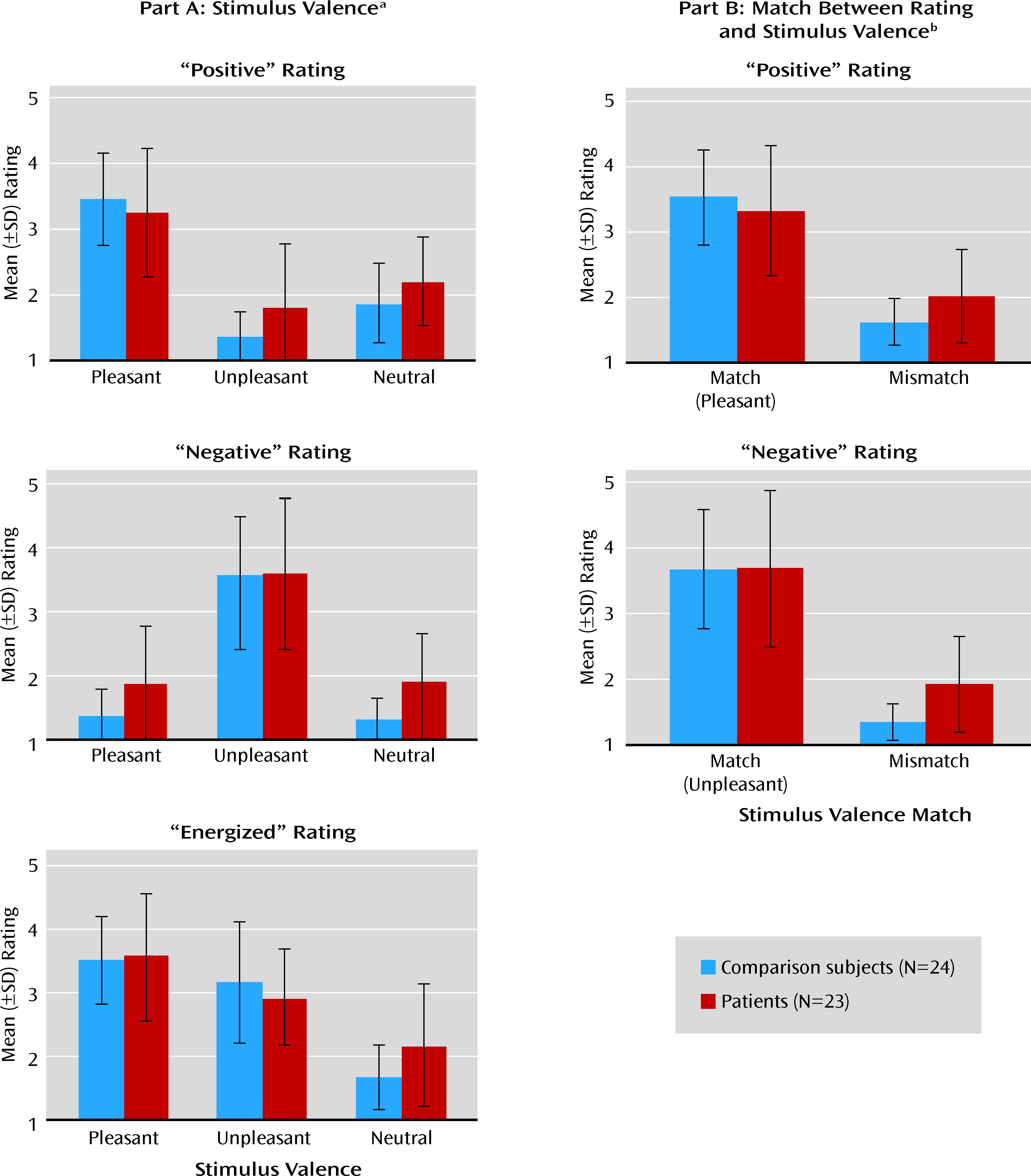
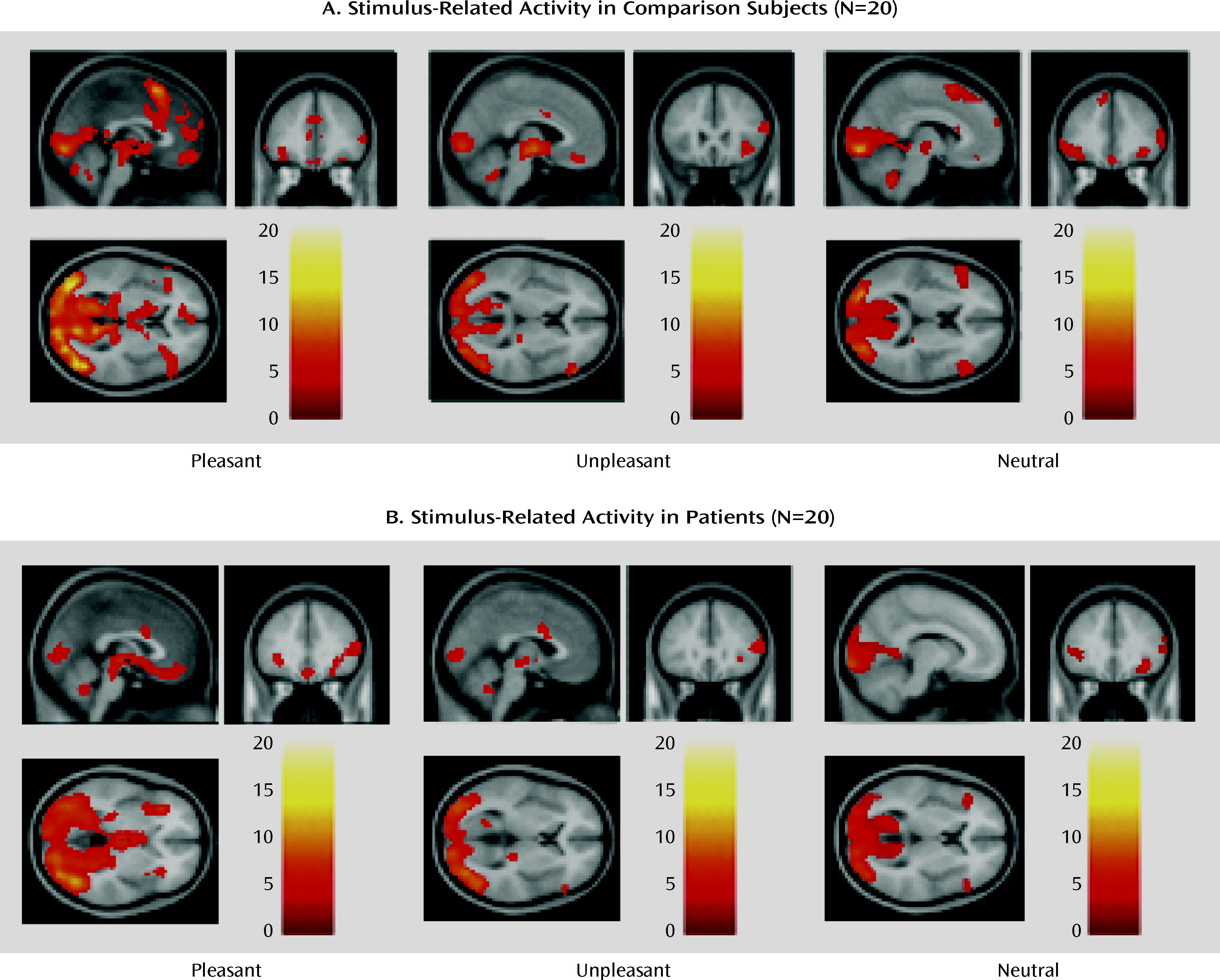
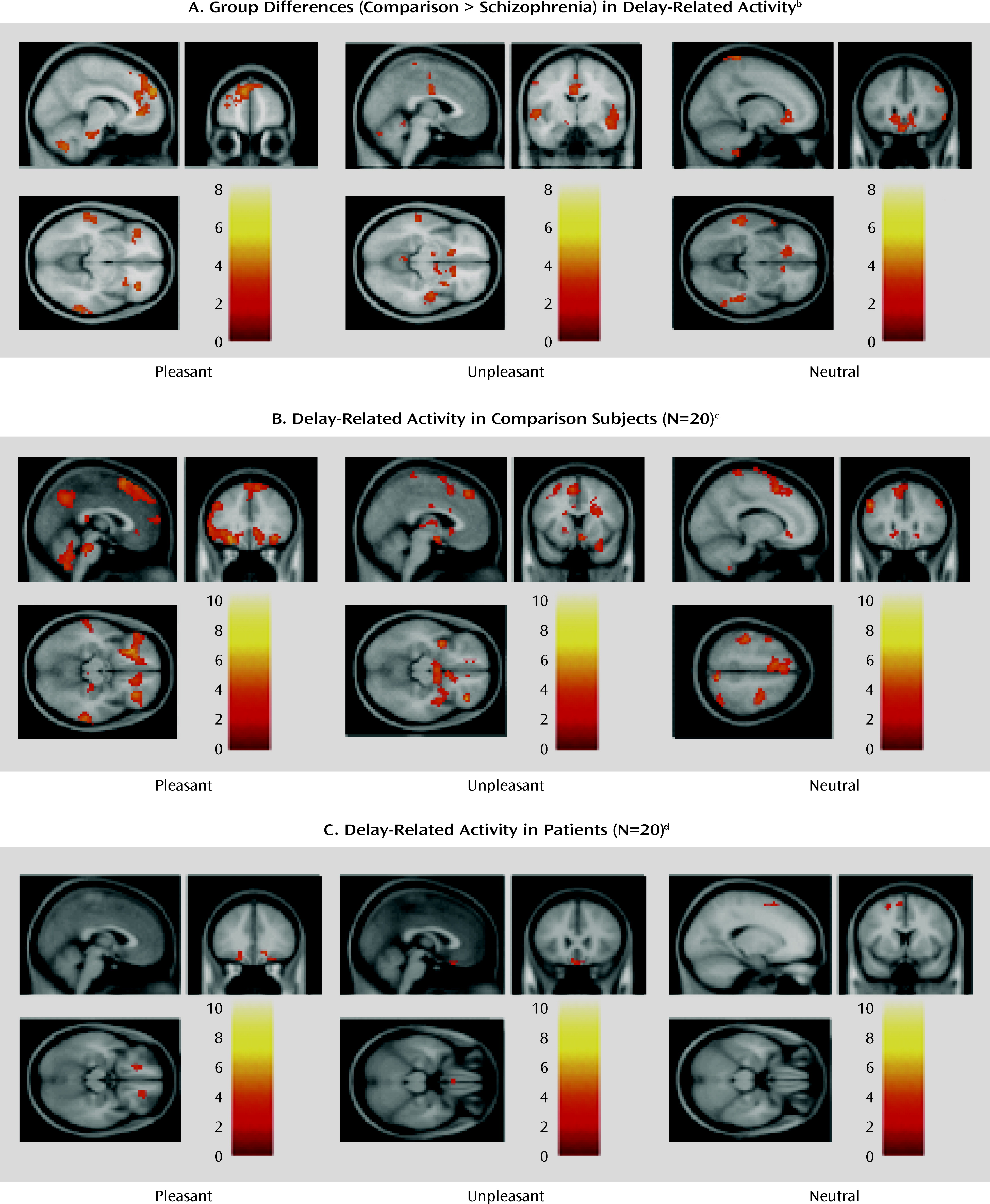
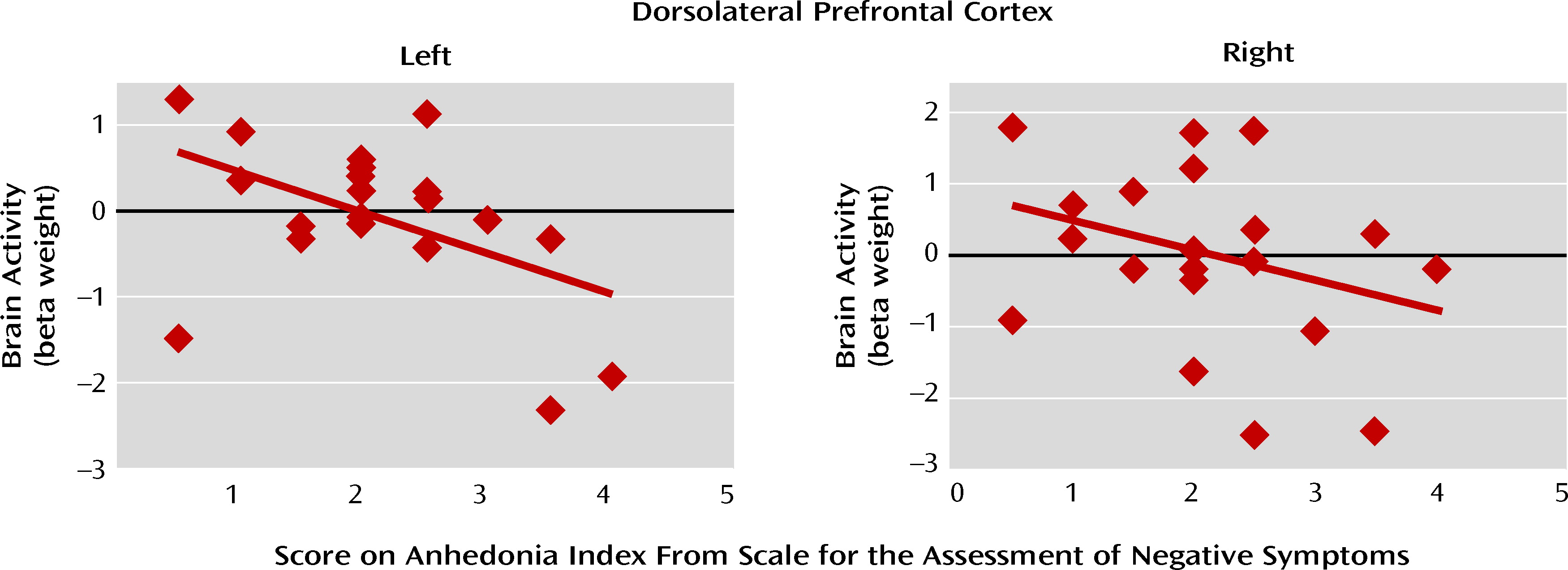
The main findings of this study were 1) in the presence of stimuli with affective content, the brain activity of schizophrenia patients was similar to that of comparison subjects in a network of prefrontal, limbic, and paralimbic structures, 2) during the delay between stimulus offset and emotion ratings (when maintenance and evaluation of past emotions required significant support from cognitive control), patients showed less activation in this network, which included the dorsolateral prefrontal cortex, and furthermore, 3) the patients' ratings of emotional experience showed evidence of impairment when the emotion was rated on dimensions incongruent with the stimulus valence, and 4) the delay-related activity elicited in the patients' dorsolateral prefrontal cortex by pleasant stimuli correlated negatively with a clinical measure of anhedonia. Taken together, these results suggest that schizophrenia may be characterized by a failure of the prefrontal circuitry supporting the critical link between affect and goal-directed behavior.
These results help shed new light on the precise mechanism behind disturbances of emotion-cognition interactions in schizophrenia, and they may provide a parsimonious account of the variability noted in previous studies. The experimental design used here allowed for a clearer separation than in previous studies between the immediate emotional experience and the control processes necessary for ratings of that experience. This has allowed us to detect robust in-the-moment affect-related brain activity in patients, as in another recent study (
24), followed by prominent hypoactivity during the delay in much of the same affect-related circuitry, as well as in control-related structures (necessary for the maintenance and reporting of affective experience after the stimulus offset). The significant negative correlation between delay-related dorsolateral prefrontal cortex activity and anhedonia supports the idea that much of the disconnect between emotional experience and behavior observed in patients, at least in laboratory contexts, may be related to dysfunctional top-down support from cognitive control processes (
14,
25). Furthermore, the variability in previous results may be accounted for, at least in part, by disturbances in the patients' neural substrates of cognitive control. It is likely that these deficits may have contributed, by means of dynamic interactions with affect-related systems (
7,
26), to manifestations of group differences in emotion-related brain activity, especially since the majority of previous studies used designs that led to significant overlap between brain activity related to experiencing emotion and that elicited by cognitive control (
24).
The translation of affective-motivational states into goal-directed behavior is thought to depend critically on effective implementation of top-down controlled processes (
8,
27). Therefore, deficits in cognitive control may also contribute to impaired representations of the value of outcomes (
28) and may account for a hypothesized decoupling of affect from motivated behavior (
29). In our study, the patients' dorsolateral prefrontal hypoactivity was weakly correlated with ratings of avolition, but the correlation did not reach statistical significance. However, since deficits in striatal responses to primary reinforcers correlated with avolition in other studies (
30), the specific role of cognitive control dysfunction in schizophrenia patients' avolition symptoms needs further exploration.
The precise nature of the dorsolateral prefrontal-dependent control processes supporting delayed reporting of emotional experience cannot be unequivocally resolved by this experiment. For instance, the timing of control processes may vary across subjects, although it is unlikely that the two groups differed in this respect. Notably, post hoc analyses of correlations between brain activation and task performance (Table S1 in the online data supplement) provided indirect evidence consistent with a critical role of sustained dorsolateral prefrontal cortex activity in delayed ratings of emotional experience, in particular for ratings that did not match the valence of the stimulus (i.e., mismatch ratings). Furthermore, the patients' abnormalities in the delayed mismatch ratings appeared related primarily to dysfunction of the dorsolateral prefrontal cortex, rather than other brain areas previously proposed to be dysfunctional in schizophrenia, such as the hippocampus, or other structures involved in affective representations, such as the orbitofrontal cortex (see “Comparison of DLPFC and Other Brain Areas…” in online supplemental text file). Considering the numerous previous reports that dysfunction of the same prefrontal area (Brodmann's area 9) correlates with failure of cognitive function in schizophrenia, even when emotional states are irrelevant to task performance, this dorsolateral prefrontal functional deficit is probably not valence specific or even emotion related. Instead, it is likely that prefrontal representations are the actual contextual link between affective states and task-appropriate responses, and these representations appear to degrade faster in patients, similar to previously described deficits in task context processing (
11). This may also explain why the patients' performance was similar to that of the healthy subjects when such representations were more readily available (i.e., for ratings matching the stimulus valence). Nevertheless, these mechanisms remain to be definitively characterized by more studies involving systematic manipulations of the affective aspects and the cognitive demands of the task.
Another potential limitation is that most patients were chronically ill and, consequently, medicated. It is unlikely that medication status can fully account for our findings, as previous results have shown emotional responding not to be strongly affected by medication (
31) and deficits in prefrontal cognitive processes have been documented in first-episode and never-medicated schizophrenia patients (
11,
12). Nevertheless, future studies of individuals at risk for developing schizophrenia or of medication-naive first-episode patients would be helpful in understanding what, if any, role medication or illness duration has in this type of task.
It is interesting that in the within-group statistical maps of stimulus-related activity, some activations in the presupplementary motor and supplementary motor areas appeared to be more extensive in healthy subjects than in patients (
Figure 3), and these areas have been implicated in various aspects of motor control (
32). While this apparent difference was not statistically significant, it could, theoretically, reflect a real but small effect. Indeed, such an effect would be consistent with the hypothesized reduced cognitive control in schizophrenia patients. Another aspect worth noting is that the delay between stimulus offset and experience rating was evidently short enough so that the patients and comparison subjects did not differ in their matched ratings, a finding that has been repeatedly reported when ratings were not delayed (
15,
25,
33,
34). However, differences did emerge in ratings of experience that did not match the stimulus. These findings extend previous reports of stronger aversion reported by patients in response to positive and neutral stimuli (
35), as well as stronger arousal in response to neutral stimuli (
4), but their precise mechanisms remain elusive. Previous results suggest that encoding of the valence of emotional stimuli, at least negative stimuli, depends on controlled processes supported by the dorsolateral prefrontal cortex (
36). Thus, rating emotional experiences on dimensions inconsistent with the stimulus valence may require support from control processes in order to overcome prepotent responding to a valence that is a match between stimulus and rating type. Our additional correlational analyses support this idea (see Table S1 in the online data supplement). Nevertheless, further research is necessary to confirm that this mechanism contributes to the higher mismatch ratings in patients (see also reference
35).
In conclusion, the results presented here suggest that a disturbance of prefrontal cognitive control processes may account to a greater degree than previously thought for the hypothesized dysfunctional interaction of cognition and emotion in schizophrenia. As such, they provide a basis for formulating and testing hypotheses in future research aimed at investigating the nature of neural deficits in this disorder, their clinical correlates, and even possible therapeutic approaches.