Common signatures of dysfunction across disorders are rapidly accumulating in indices as wide-ranging as neuroinflammatory cytokines (
1), resting regional cerebral blood flow (
2), genetics (
3), and white matter integrity (
4). Notably, across disorders, psychosocial functional impairment is better explained by shared as opposed to disorder-specific cognitive and affective dysfunction (
5). Thus, leveraging these common neurobehavioral phenotypes has promise in advancing therapeutics that could be flexibly applied across disorders.
Neuroimaging meta-analysis approaches such as activation-likelihood estimation (ALE) allow for testing of spatial convergence of findings across studies (
6), thus consolidating the neuroimaging literature into quantitative evidence for common dysfunction. While this provides limited information regarding small effects that may be unique to a specific disorder, it is a key approach for identifying those brain regions most likely to contribute to dysfunction across individuals, processes, and disorders (
7). Furthermore, psychiatric multimorbidity is far more common than unimorbidity. However, across studies, the procedure for identifying the primary disorder of interest varies. Hence, an explicitly transdiagnostic approach to interpreting meta-analytic results would likely be more reflective of the cumulative burden of psychopathology on neurocircuitry.
In a previous meta-analysis of 193 voxel-based morphometric studies of gray matter volume (N=15,892) (
8), we found that gray matter was reduced in three regions—the dorsal anterior cingulate and the left and right anterior insula—across a diverse array of disorders (schizophrenia, bipolar and unipolar depression, substance use, and anxiety disorders). Moreover, lower gray matter volume in these regions, even in healthy individuals, predicted worse behavioral performance on measures of higher-level cognition and executive function. We subsequently found, in a parallel meta-analysis of 283 cognitive control task–related activation studies (N=11,218), that similar disruptions were evident across disorders in these and related regions (
9) (i.e., the prefrontal cortex [from premotor to mid-dorsolateral prefrontal cortex], the anterior insula extending to the ventrolateral prefrontal cortex, the intraparietal sulcus, and the anterior midcingulate/presupplementary motor cortex), which together make up the “multiple demand” cognitive control network (
10,
11). These findings suggest a coordinated transdiagnostic structural and functional perturbation of an interconnected set of regions that are likely associated with transdiagnostic deficits in cognitive control performance (
12). Consistent with the work of others (
13–
15), it appears that networks required for adaptive cognition are structurally and functionally vulnerable to broad-spectrum psychopathology.
In this study, we examined whether there is a parallel transdiagnostic functional impairment in whole brain activation during emotional processing. Similar to the transdiagnostic findings on cognition, we expected that deficits would be most pronounced in regions that subserve adaptive emotional reactivity, regulation, and recovery. While the amygdala has long been the focus of disordered emotional processing, we expected that a distributed network of regions would converge in conjunction with amygdala hyperactivation (
16,
17). More specifically, we hypothesized that transdiagnostic dysfunction would additionally manifest in the hippocampal/parahippocampal gyri, thalamic nuclei, and fusiform gyri, particularly as hyperactivation in the patient group (
16). Furthermore, we hypothesized that lateral prefrontal regions associated with regulation of emotion and cognition (
18) would demonstrate dysfunction, particularly in tasks calling for suppression or control of affect. Finally, we hypothesized that disruption in emotional processing neurocircuitry would be inclusive of regions shown in our previous meta-analyses to be disrupted, especially the anterior-cingulo-insular network.
Methods
Experiment Inclusion Criteria and Identification
All procedures were consistent with the guidelines outlined by Müller and colleagues (
19). Articles were identified by searching PubMed and previous meta-analyses and review articles for functional neuroimaging experiments of emotional processing tasks published through May 2018 that compared patients with psychiatric disorders to matched control participants (see Figure S1 in the
online supplement). Experiments were eligible if they included emotional processing tasks with functional neuroimaging, whole-brain analysis, comparison between patients and matched healthy control participants, and coordinates in a defined stereotaxic space (e.g., Talairach or Montreal Neurological Institute [MNI] space).
Experimental procedures must have included a diagnostic interview of patients and control participants, with patient groups exceeding the threshold for diagnosis. A psychotic disorders category comprised schizophrenia and schizoaffective, schizophreniform, and delusional disorders. A nonpsychotic disorders category comprised bipolar, unipolar (major depression, dysthymia) depressive, anxiety (including obsessive-compulsive and posttraumatic stress), and substance use (mixed substance abuse and/or dependence) disorders.
Across disorders, patient participants included those with first-episode and chronic disorder manifestations, including interepisode states of bipolar and psychotic disorders. The substance use disorders included chronic users of a range of substances, currently active or abstinent but not in acute withdrawal. Experiments were selected to capture lifespan patterns and thus included participants ranging from childhood through older adulthood. Diagnoses presenting predominantly in childhood (e.g., attention deficit hyperactivity disorder) or those associated with altered developmental trajectories of brain structures inherent to expression of disorder phenotypes (e.g., autism spectrum disorders) were excluded.
Articles with experimental tasks probing emotional processing were included, broadly categorized into reactivity (e.g., passive face/picture viewing, fear conditioning), regulation (e.g., reappraisal/suppress), and compound (i.e., emotional targets in compound tasks such as n-back, go/no-go, delayed discounting, Stroop) (see Table S1 in the online supplement).
Peak coordinates for whole-brain between-group comparisons during emotional challenge (relative to a baseline/neutral condition) were required. Condition-by-group interactions were included if follow-up tests clarified patterns of patient hyper- versus hypoactivation during emotional processing. Experiments reporting results only for small-volume correction or within a region of interest were excluded. All coordinates were converted to MNI space (
20).
Activation Likelihood Estimation (ALE) Meta-Analysis
The revised ALE algorithm, implemented in MATLAB, was used to identify areas of spatial convergence of reported coordinates for differences between patients and control participants in activation during emotional processing tasks that were higher than expected under a random spatial association (
6,
21,
22) (see the Supplementary Methods section in the
online supplement). Resulting nonparametric p values were thresholded at a cluster-level family-wise-error-corrected p value of 0.05 (cluster-forming threshold at voxel-level p<0.005) and transformed into z scores for display. To avoid results dominated by one or two individual experiments and to have sufficient power to detect moderately sized effects, ALE was interpreted only for those analyses that included at least 17 experiments (
23).
We conducted the following analyses: We first pooled across coordinates for hypo- and hyperactivation in patients relative to control participants to identify transdiagnostic patterns (i.e., patterns observed when diagnostic category is not considered) of “aberrant activation,” thus pointing to prominent hubs of dysfunction. More refined analyses were then performed separately for activation coordinates associated with hyperactivation (patients > controls) or hypoactivation (patients < controls). Furthermore, we conducted these analyses on the following parcellations of the emotional processing literature:
1.
Across all patients and tasks.
2.
Separated by the type of emotional processing (reactivity, regulation, compound).
3.
Separated by psychotic versus nonpsychotic disorders.
4.
Separated by broad diagnostic spectra or groupings (i.e., schizophrenia, bipolar and unipolar depression, substance use, and anxiety disorders) and by individual disorders.
Follow-up ALE analyses were also performed to test for the contribution of demographic characteristics, medication status, and task- and stimulus-related factors.
Of note, the aim of this meta-analysis was to highlight transdiagnostic patterns. Nonetheless, while separate, diagnosis-specific analyses have been performed previously, each of these previous meta-analyses utilized different inclusion and exclusion criteria and thus included different experiments. We therefore performed subanalyses to aid in interpreting the primary transdiagnostic analyses. Moreover, because the subanalyses came out of an overall analysis that employed the same study selection process and data analysis methods across all diagnoses, the diagnosis-specific subanalyses can now be more meaningfully compared with each other.
Results
Final Selected Experiment Set
The final set of experiments consisted of 298 experiments from 254 articles (see Figure S1 and Tables S1 and S2 in the online supplement) covering 10,918 participants (5,427 patients and 5,491 control participants). (For a more detailed description of included experiments, see the online supplement.) The vast majority of experiments (N=283) used functional MRI (fMRI), followed by 14 positron emission tomography experiments and one using single-photon emission computed tomography. Mean ages ranged from 12.1 to 73.1 years (mean=32.5, median=33.2). Most experiments were with adults (N=273; age range, 18–50 years; mean age, 44.40 years), followed by children and adolescents (N=21; <18 years; mean age, 14.76 years), with very few studies with older adults (N=4; >50 years; mean age, 60.27 years).
The psychotic disorders category comprised experiments exclusively with schizophrenia patients (N=59). The more heterogeneous nonpsychotic disorders category comprised 239 experiments investigating patients with anxiety disorders (N=94; posttraumatic stress disorder [PTSD], N=28; obsessive-compulsive disorder [OCD], N=20; social anxiety disorder, N=17; specific phobia, N=11; generalized anxiety disorder, N=10; panic disorder, N=5; mixed anxiety disorders, N=3), unipolar depressive disorders (N=79), bipolar disorders (N=47), and substance use disorders (mixed substance abuse and/or dependence disorders, N=19; cocaine, N=8; alcohol, N=6; methamphetamine, N=3; cannabis, N=2). Across disorders, patient participants included those with first-episode and chronic disorder manifestations, as well as interepisode expressions in the case of chronic bipolar and psychotic disorders (11 of 47 and four of 59, respectively). Reports of comorbidity were inconsistent across studies, precluding quantification.
On the task level, experiments represented three broad types of emotional processing: reactivity (N=230), compound emotional/cognitive (N=56); and regulation (N=18). Tasks spanned multiple domains: picture viewing (N=161), script reactivity/imagery (N=20), Stroop (N=15), conditioning (N=12), and diverse tasks, none of which exceeded more than 11 studies. A majority of experiments included medicated (N=174) as opposed to unmedicated patients (N=119); information on medication was lacking for five experiments.
Meta-Analysis Results Across Disorders
Aberrant Activation Patterns During Emotional Processing
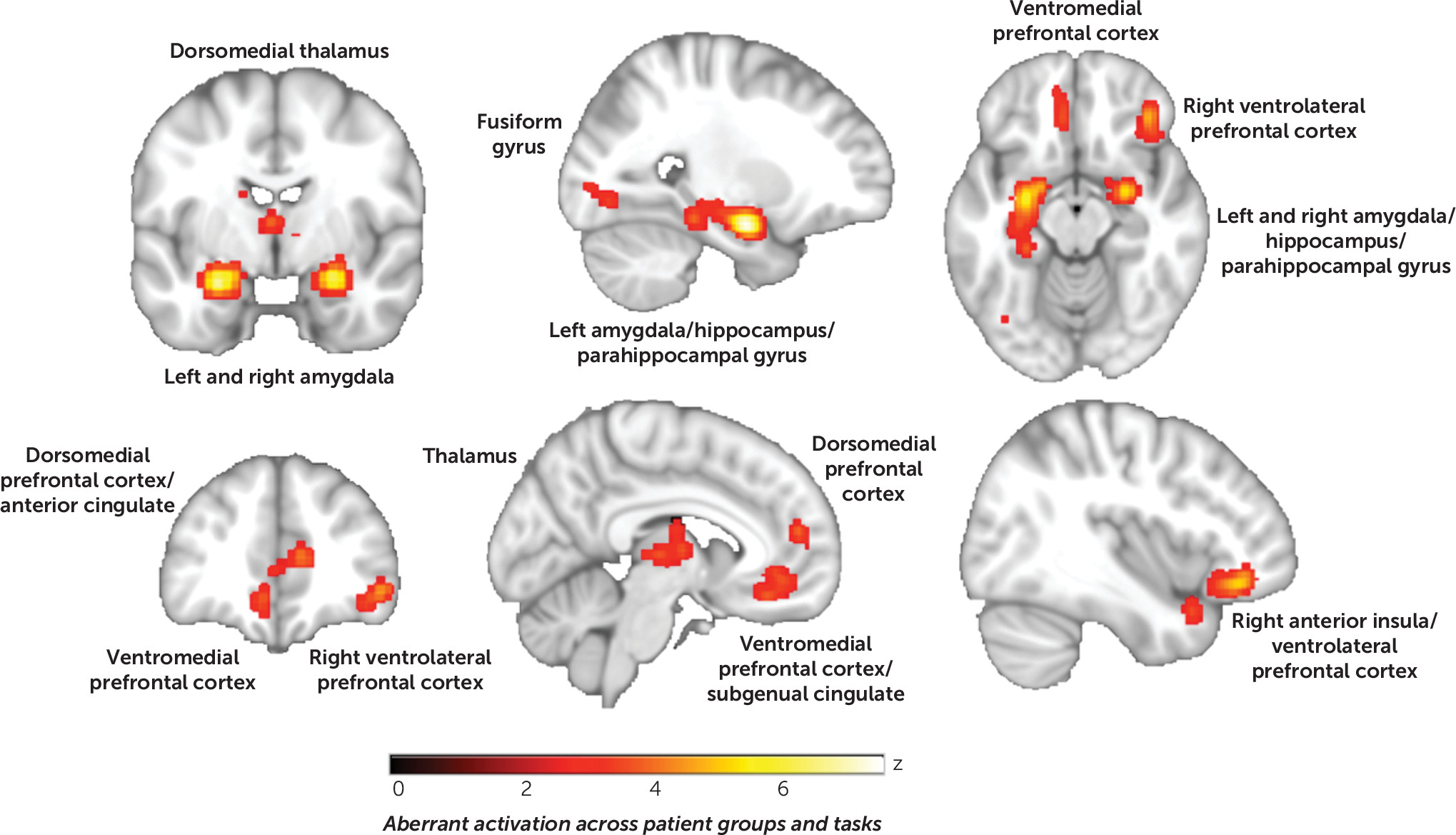
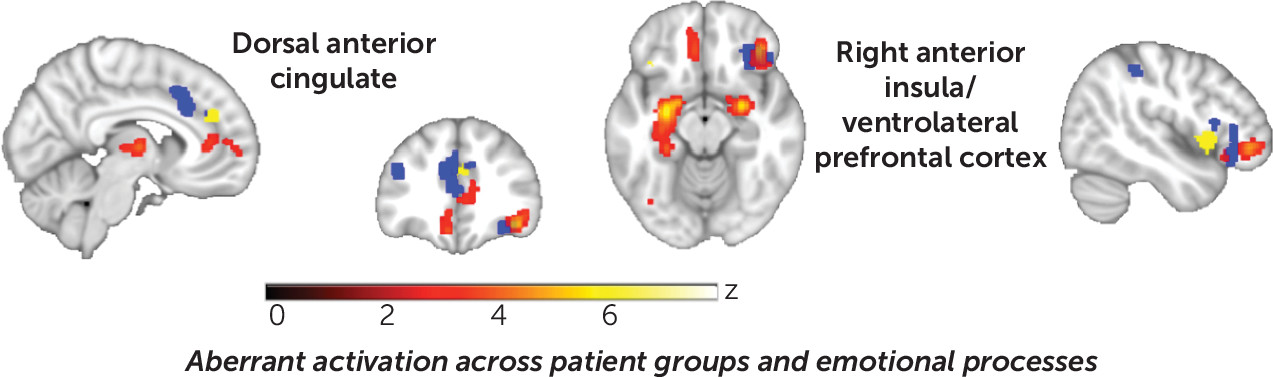
Aberrant activation across disorders and processing types.
The first ALE entailed pooling across 1) disorders, 2) patterns of patient hyper- and hypoactivation, and 3) processing types to assess “aberrant activation.” We found consistent abnormalities in the ventromedial prefrontal cortex extending to the subgenual cingulate cortex, the right insula extending to the ventrolateral prefrontal cortex, the thalamus (dorsomedial extending to pulvinar nuclei), the left and right amygdala extending to hippocampal and parahippocampal gyri, and the inferior occipital cortex (
Figure 1; see also Table S2 in the
online supplement). The contributions of individual experiments to the observed clusters of convergence according to sample and design characteristics are reported in detail in Table S3 in the
online supplement. In summary, consistent with a common signature shared to varying degrees across diagnoses, a broad distribution of disorders contributed to each of the clusters of convergence. This was confirmed by follow-up region-of-interest analyses on extracted data from the six significant clusters, which demonstrated similar probabilities of voxelwise activation from the modeled activation maps across disorder groupings (Kruskal-Wallis test, H values, 7.86–0.56, n.s.) (see Figure S2 in the
online supplement). Additionally, experiments with adults (ages 18–50), fMRI, and unpleasant emotional reactivity overwhelmingly contributed to convergence, as did face and scene processing tasks. Results from medicated and unmedicated samples contributed similarly across clusters. Corresponding ALE results stratified by sample and design characteristics are provided in Figures S3–S9 in the
online supplement.
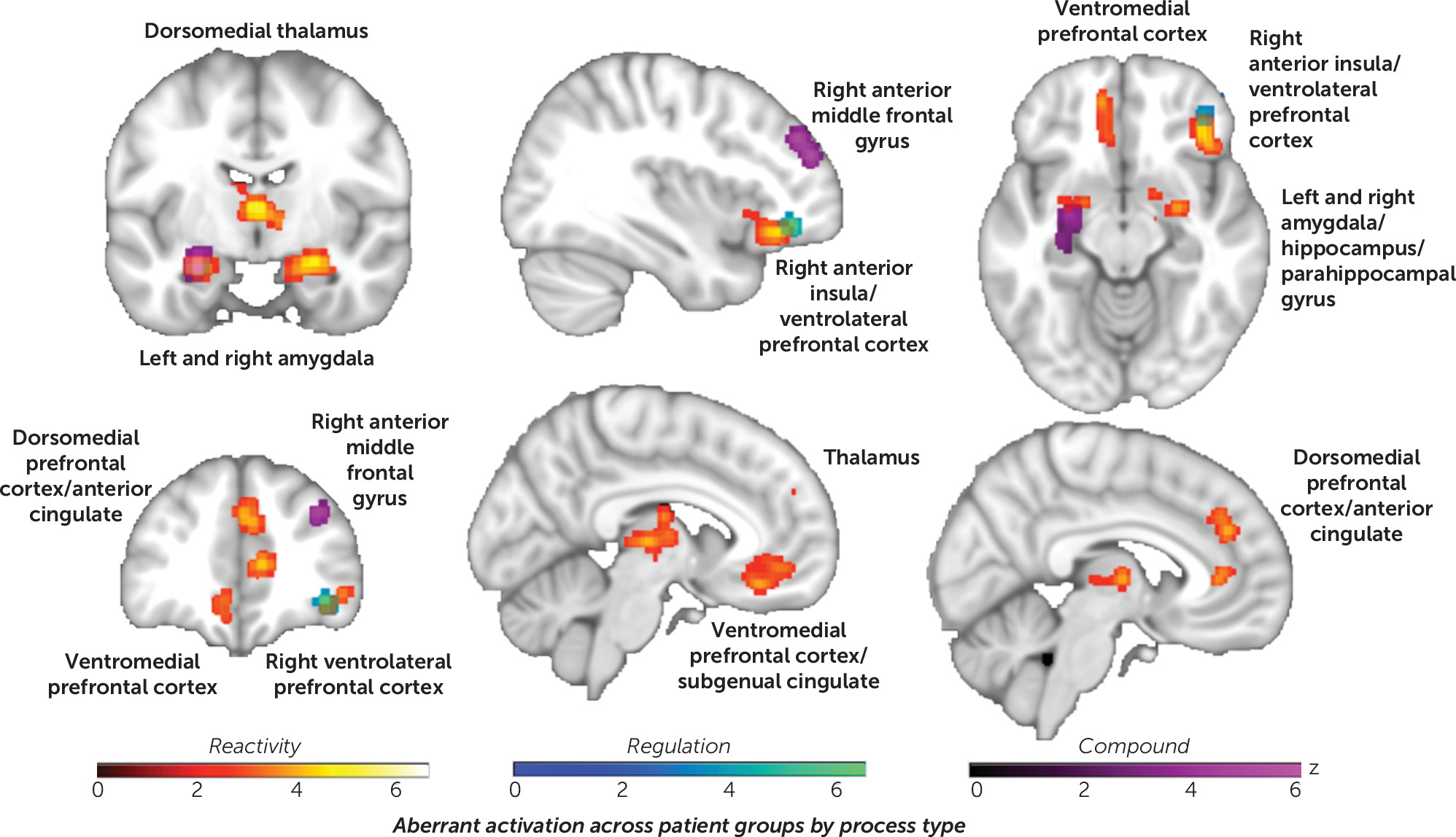
Aberrant activation across disorders separated by processing type.
Separating ALE by the emotional process involved (i.e., reactivity, regulation, or compound) (
Figure 2; see also Table S4 in the
online supplement) revealed that passive reactivity was characterized by convergent disruption in the dorsal anterior cingulate, the dorsomedial/pulvinar thalamic nuclei, the ventromedial, dorsomedial, and right ventrolateral prefrontal cortex, as well as the left and right amygdala and the hippocampal and parahippocampal gyri. Compound tasks were predominantly characterized by disruption in the left amygdala/hippocampus and the right anterior middle frontal gyrus. Regulation tasks, by contrast, showed convergent disruption specific to the right ventrolateral prefrontal cortex. Contrasts confirmed these distinctions between processes (see Figure S10 in the
online supplement).
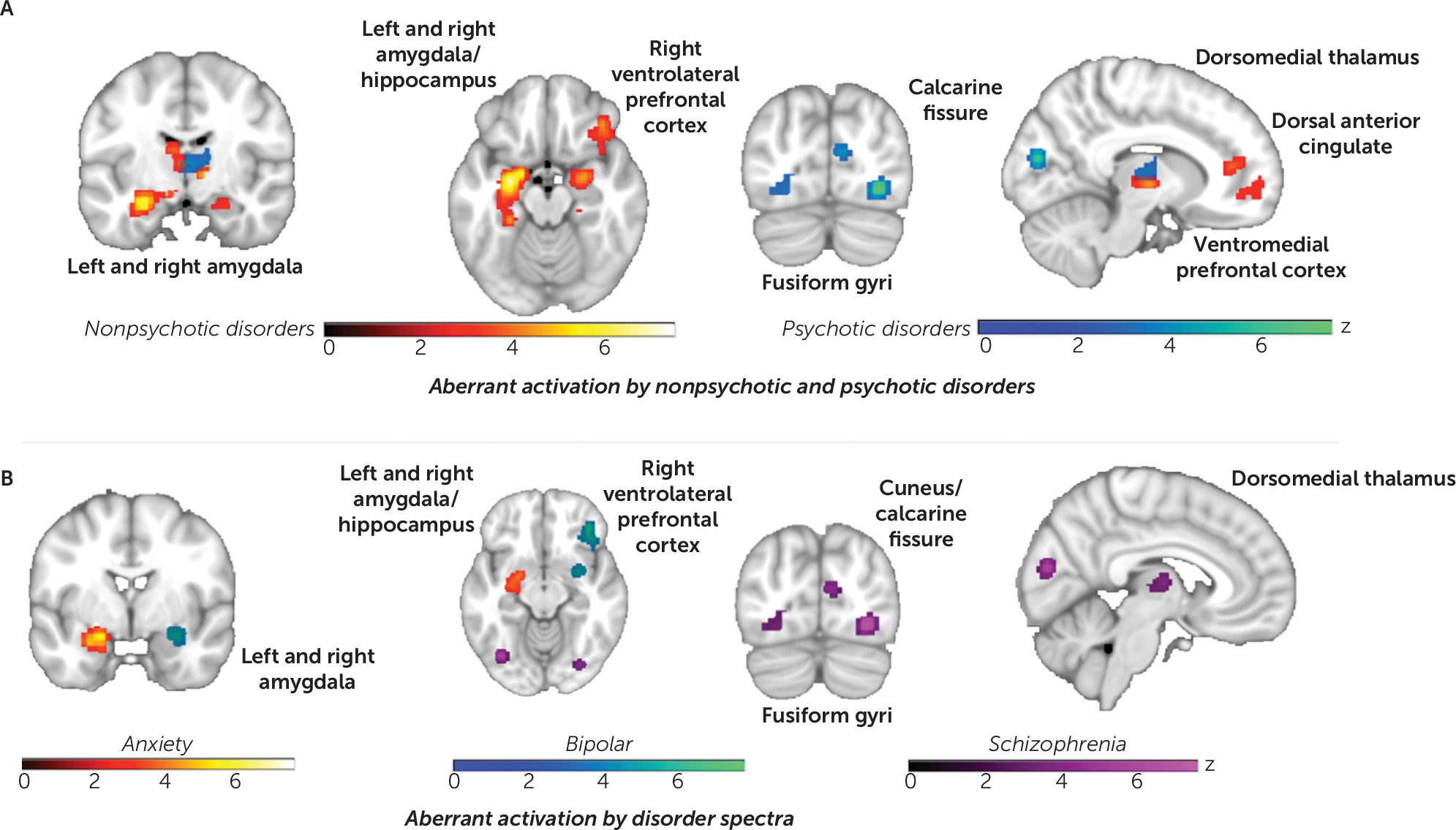
Aberrant activation by disorder groupings.
Separating studies by nonpsychotic versus psychotic disorders (
Figure 3A; see also Table S5 in the
online supplement) revealed that nonpsychotic disorders were characterized by aberrant activation of the left and right amygdala, the hippocampal and parahippocampal gyri, the dorsal anterior cingulate, the ventromedial and dorsomedial prefrontal cortex, and the dorsomedial thalamus. Psychotic disorders were characterized by convergent aberrations in the dorsomedial thalamus as well as the calcarine fissure and the left and right fusiform gyri. The reliability of these apparent differences between psychotic/control and nonpsychotic/control patterns was confirmed in direct between-group contrasts (see Figure S11 in the
online supplement).
Finer resolution by disorder spectra (i.e., anxiety, depression, bipolar, schizophrenia, and substance use) (
Figure 3B) highlighted convergent aberrant activation in the left amygdala and hippocampus in anxiety disorders. Bipolar disorder showed convergence in the right amygdala and right ventrolateral prefrontal cortex, whereas schizophrenia was characterized by disruption in the dorsomedial thalamus, the calcarine fissure, and the fusiform gyri. In contrast, similar to a previous meta-analysis and despite ample power to detect effects (
24), unipolar depressive disorders showed no convergent patterns. Substance use disorders also showed no convergence. Contrasts as well as refinement to individual disorder categories were consistent with these patterns (see Figures S12 and S13 in the
online supplement).
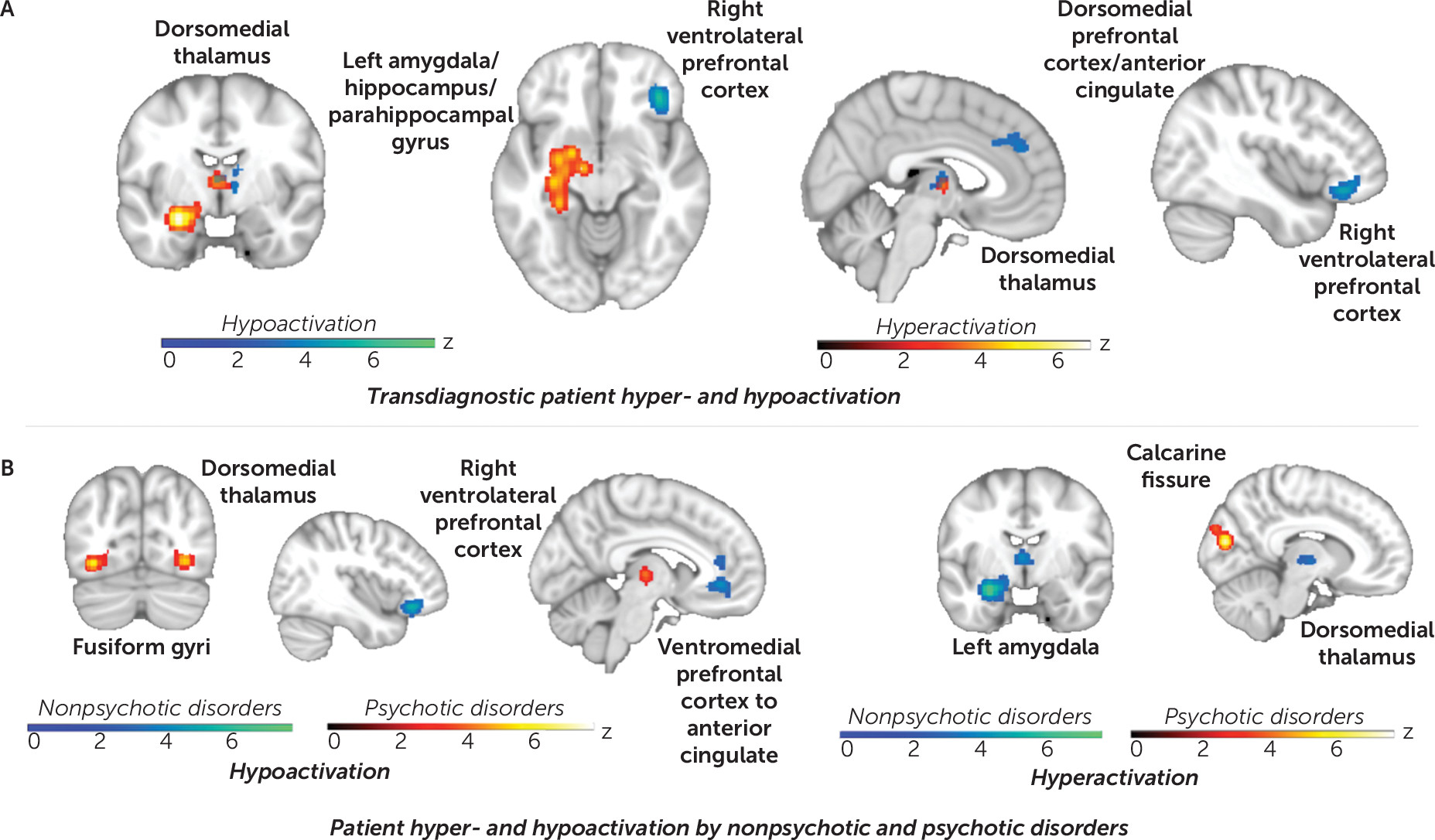
Hyper- versus hypoactivation across disorders and processing type.
Testing separately for convergent hyper- versus hypoactivation in patients demonstrated that patient hyperactivation (pooled across disorders and processes) emerged in the amygdala, the hippocampal/parahippocampal gyri, and the dorsomedial thalamus. In contrast, patient hypoactivation was more evident in medial and lateral prefrontal regions, as well as in the dorsomedial thalamus (
Figure 4A; see also Tables S6 and S7 in the
online supplement). The specific contributions by broad disorder grouping are presented in Figure S14 in the
online supplement. (Regarding the laterality of amygdala and ventrolateral prefrontal effects, if the statistical threshold is reduced, the resulting network disruptions are bilateral and symmetrical, consistent with the structure of the canonical networks.)
Hyper- versus hypoactivation across disorders by processing type.
Separating ALE by patient hyper- versus hypoactivation as well as emotional process involved (i.e., reactivity, regulation, or compound) (see Figure S15 and Table S8 in the
online supplement) revealed that passive reactivity was characterized by patient hyperactivation in the amygdala, the hippocampus, the parahippocampal gyrus, the dorsomedial thalamus, and the ventromedial prefrontal cortex/subgenual cingulate. Hypoactivation during passive reactivity converged in the dorsal medial thalamus and the right ventrolateral prefrontal cortex, a region shown to activate during intact emotion regulation and confirmed by conjunction with networks identified in a meta-analysis of intact emotion regulation (
18) (see Figure S16 in the
online supplement). Compound tasks were predominantly characterized by patient hyperactivation in the left amygdala, the hippocampus, and the parahippocampal gyrus and hypoactivation in the caudate/putamen and the right anterior middle frontal gyrus. A conjunction with the canonical networks identified by Shirer and colleagues (
25) demonstrated that the hypoactive right anterior middle frontal gyrus cluster overlaps with a node of the anterior salience network (see Figure S17 in the
online supplement) and is more anterior than the cluster typical in emotion regulation (
18). Regulation tasks showed no convergence when patient hypo- and hyperactivation were analyzed separately, likely because of the limited number of included contrasts.
Hyper- versus hypoactivation by disorder groupings.
Separating ALE by patient hyper- versus hypoactivation as well as psychotic and nonpsychotic disorders (
Figure 4B; see also Table S9 in the
online supplement) demonstrated that nonpsychotic disorders showed convergent hyperactivation in the left amygdala and the dorsomedial thalamus, coupled with hypoactivation in the ventromedial and right ventrolateral prefrontal cortex as well as the dorsal anterior cingulate. Psychotic disorders showed hyperactivation in the cuneus/calcarine fissure. Hypoactivation was evident in the dorsomedial thalamus as well as the fusiform gyri, overlapping with regions particularly responsive to faces (
26). Contribution statistics indicated that, in fact, experiments with faces contributed to 83% of the contrasts (see Table S7 in the
online supplement).
Finer group resolution by disorder spectra (i.e., anxiety, depression, bipolar, schizophrenia, and substance use) (see Figure S18 and Table S10 in the online supplement) highlighted overlapping hyperactivation in the left amygdala and the hippocampus in anxiety and unipolar depressive disorders. Bipolar and substance use disorders showed overlapping hypoactivation of the right ventrolateral prefrontal cortex. Schizophrenia was equivalent to the psychotic disorders group. Region-of-interest analyses further confirmed these findings (see Figure S19 in the online supplement). Refinement to individual disorder categories (e.g., specific phobia, alcohol use disorder, OCD) showed convergence consistent with these patterns (see Figure S20 in the online supplement), although numerous categories had fewer contrasts than warranted for valid inference.
Overlap of structural and functional disruptions across disorders.
Figure 5 illustrates the close correspondence of the regions shown here to be disrupted during emotional processing (red) to those we previously demonstrated to be marked by gray matter reduction (
8) as well as functional disruption during cognitive control tasks across disorders (
9). In the medial wall of the prefrontal cortex to the anterior cingulate, a gradient is evident, with emotional more anterior, cognitive disruption more posterior, and gray matter loss at the intersection. Disruptions of emotional processing and cognitive control showed the most correspondence in the right ventrolateral prefrontal cortex, with gray matter loss slightly more posterior in the anterior insula.
Discussion
In a meta-analysis of emotional processing tasks across psychiatric disorders, we observed a common pattern of aberrant brain activation, shared to varying degrees across diagnoses. The pattern emerged in regions typically associated with emotional reactivity and regulation, including the left and right amygdala extending to the hippocampal and parahippocampal gyri, the ventromedial prefrontal cortex extending to the subgenual cingulate, the right anterior insula extending to the ventrolateral prefrontal cortex, the dorsomedial and pulvinar thalamic nuclei, and the inferior occipital cortex. Owing to the overrepresentation of passive reactivity tasks, this overall pattern largely reflected passive viewing, primarily of unpleasant scenes and faces. Regulation of emotional reactions prompted convergence limited to the right ventrolateral prefrontal cortex, while compound tasks showed convergence in both the amygdala/hippocampus and the right anterior middle frontal gyrus. The latter may reflect the switching and updating required in tasks such as the Stroop and n-back (
27). Considering patient hyper- versus hypoactivation separately across disorders and processes revealed excessive activation of the amygdala and the hippocampal and parahippocampal gyri coupled with hypoactivation in the medial and lateral prefrontal regions. Interestingly, clusters of both hyper- and hypoactivation converged in the dorsomedial thalamus.
Ventrolateral Prefrontal Cortex Abnormalities
The right ventrolateral prefrontal/orbitofrontal cortex demonstrated hypoactivation in patients across disorders. Consistent with its proposed role in signaling “nonreward” (
28), dysfunction was most pronounced during unpleasant stimuli/paradigms as well as during both reactivity and regulation processes. A conjunction with our previous voxel-based morphometry and cognitive control transdiagnostic meta-analyses demonstrated that disruptions of emotional processing and cognitive control showed the most correspondence in this region, with gray matter loss located slightly more posteriorly within the anterior insula. The ventrolateral prefrontal cortex and the anterior insula together constitute central bilateral nodes in the “multiple demand network” that subserves domain-general cognitive control (
10,
11). Consistent with the involvement of the ventrolateral prefrontal cortex in domain-general cognitive regulation is its demonstrated role in intact emotion and action regulation (
18). In fact, this region has been shown to be central to affective and nonaffective inhibitory control among the same participants and to show concurrent disruptions in psychopathology (
29). Regarding its convergence in both reactivity and regulation tasks, the ventrolateral prefrontal cortex has both afferent and efferent connections to the amygdala and sensory cortices, as well as afferent connections from the anterior insula (
30). Additionally, it is heavily interconnected to regions involved in emotional expression (e.g., the frontal pole) and regulation (the dorsolateral prefrontal cortex). Dysfunction of this region has been linked to perseverative negative bias in depression (
31), and the present findings suggest that this region may contribute more broadly to perseverative rumination observed across disorders (
32).
The ventrolateral prefrontal cortex and its afferent and efferent connections appear to be a hub in the coordination of emotional experience. Accordingly, neuromodulation-based interventions that target this region have shown promise across disorders. A recent study of intracranial electroencephalography in epilepsy patients showed that phasic fluctuations in alpha and theta band power specifically in the orbitofrontal cortex corresponded to simultaneous fluctuations in subjective dysphoria (
33). Furthermore, high-frequency electrical stimulation of this area suppressed theta and alpha power, corresponding to immediately improved mood, specifically among patients with moderate to severe comorbid depression. In OCD, 1-Hz therapeutic repetitive transcranial magnetic stimulation (rTMS) to the ventrolateral prefrontal/orbitofrontal cortex has reduced subjective obsessions and compulsions as well as metabolism in the left and right orbitofrontal cortex (
34). Among patients with major depressive disorder who failed to respond to a trial of rTMS to the dorsomedial prefrontal cortex, 24% achieved remission after a subsequent course of rTMS to the ventrolateral prefrontal/orbitofrontal cortex (
35).
Ventromedial Prefrontal Cortex Abnormalities
The ventromedial prefrontal cortex/subgenual cingulate also showed convergence in ALE across disorders. Accumulating work, including a recent systematic review and meta-analysis by Hiser and Koenigs (
36), demonstrates that the ventromedial prefrontal cortex is activated during perception of both appetitive and aversive cues, as well as value-based decision making, inhibition or regulation of emotions, and processing of social and self-relevant cues. Furthermore, lesions to the ventromedial prefrontal cortex have impaired performance on such tasks (
37). Hiser and Koenigs (
36) and others (
38,
39) have further highlighted transdiagnostic impairments in flexible and efficient recruitment of this region during a wide range of emotional tasks, suggesting a potentially domain-general impairment in determining motivational contingencies.
Dorsomedial Prefrontal Cortex Abnormalities
The dorsomedial prefrontal cortex, extending to the dorsal anterior cingulate, also demonstrated convergence as a region of aberrant activation in ALE across disorders. Functional parcellation of the medial prefrontal cortex in healthy samples has suggested that the more ventral portion observed here is more characterized by bottom-up, stimulus-based evaluation, whereas the dorsal portion observed here is more representative of top-down, meta-cognitive, self-oriented processes such as mental imagery, rumination, and complex social processes such as empathy (
40–
45). Subparcellation of the dorsomedial prefrontal cortex has further specified that the observed cluster of convergence comprises two smaller contiguous regions, with the left strongly connected to the fronto-parietal network and the right strongly connected to the salience network (
23,
43) (see Figure S21 in the
online supplement). As such, it is well positioned as a hub for promoting higher-order affective processing and appraisal (
46). Regarding activation, both hyper- and hypoactivation of the dorsomedial prefrontal cortex during tasks like mental imagery have been observed across and even within disorder categories, such as PTSD and depression (
47,
48). Notably, here a cluster of hypoactivation converged when considering the directionality of contrasts, with no convergent cluster of hyperactivation. However, inspection of the contribution to the dorsomedial prefrontal cluster of aberrant activation irrespective of directionality (
Figure 1; see also Table S3 in the
online supplement) revealed that patient hyper- and hypoactivation contrasts contributed equally (51% and 49% of all contrasts, respectively), suggesting substantial heterogeneity in response profiles. This heterogeneity has implications for interventions, particularly those targeting perseverative self-focus, rumination, and reflection.
In addition to proximal and/or corresponding disruption in structure and cognitive and emotional function in the right ventrolateral prefrontal cortex and anterior insula, a gradient was evident in the dorsomedial prefrontal cortex/anterior cingulate, with emotional more anterior dorsal cingulate and cognitive disruption more midcingulate—a rostral-to-caudal finding well established in functional segregation of task-based and resting-state studies (
49). Subregions from anterior to posterior cingulate have also shown distinctions in glucose metabolism, limbic and cortical afferents, cingulospinal efferents, and electrical stimulation responses (
50). Gray matter loss in the cingulate was observed specifically at the intersection of regions of emotional and cognitive disruption. This corresponds approximately to the cytoarchitectonic conjunction of Brodmann areas 24 and 32. Haber and colleagues (
51) recently used retrograde tracing in macaques to assess afferent inputs from all areas of the frontal cortex to six sites in the anterior cingulate and found this area to be the sole, prominent connectional hub. Haber and colleagues observed a similar hub in human diffusion imaging data. While there was no complete overlap for gray matter loss and cognitive and emotional dysfunction across our three transdiagnostic meta-analyses, the regions of overlap between structural and/or functional disruptions were all prominent information-processing hubs for integrating information across cortical and subcortical networks. Essentially, the convergence of structural and functional dysfunction in the anterior cingulate subregion (as well as the ventrolateral prefrontal cortex and anterior insula) may underscore the centrality of these regions in determining network disconnection and its related cognitive and affective impairment in mental illness.
Thalamic Abnormalities
Although the left and right amygdala (and adjacent hippocampus/parahippocampus) showed prominent convergent hyperactivation in ALE across disorders, this occurred in conjunction with abnormalities in a distributed network sensitive to emotional arousal. This is noteworthy given the predominant focus on amygdala hypersensitivity to threat in animal and human models of psychopathology. For example, the pulvinar and dorsomedial thalamic nuclei showed strong convergence and have been implicated as governors of cortico
-cortical connections in deploying selective attention (
46,
52,
53), possibly via network synchronization. The pulvinar is densely connected both structurally (
54) and functionally (
55) to various cortical regions, including occipital, parietal, temporal, and prefrontal cortices (
56,
57). The dorsomedial thalamus has similar cortical connections as well as both direct and indirect projections to the amygdala (
58,
59). Taken together, disrupted thalamo-cortical and thalamo-subcortical-cortical connections may feature prominently in determining dysregulated emotional experience by disrupting adaptive selective or motivated attention.
Disorder-Specific Patterns of Abnormalities
The regions of prominent emotional disruption within this broader network differed by disorder type—foremost in the directionality of patient hyper- and hypoactivation. Nonpsychotic disorders as a group were characterized more by amygdala, hippocampal/parahippocampal gyri, and dorsomedial thalamic hyperreactivity as well as prefrontal hyporeactivity. Psychotic disorders showed more prominent disruptions in the thalamus and occipital cortex, with a hyperactive calcarine fissure concurrent with hypoactive fusiform gyri and dorsomedial thalamus. Psychotic disorders, particularly schizophrenia, may be marked by hyperreactivity of primary visual processing coupled with impaired fronto-temporal-parietal-thalamic re-entrant modulation of the fusiform gyrus to adaptively tune visual processing to motivationally relevant cues (
28,
60–
62). Anxiety disorders, not surprisingly, were marked by pronounced amygdala/hippocampal hyperactivation, bipolar disorders by ventrolateral prefrontal cortex hypoactivation, unipolar disorders by amygdala hyperactivation, and substance use disorders by ventrolateral prefrontal cortex hypoactivation. While across disorders disruptions were observed in a coordinated network of regions sensitive to emotional expression and regulation, these findings suggest that therapeutics tuned to specific disruptions within the network may be preferentially effective for some neurobehavioral phenotypes.
Limitations
The number of included experiments in each of the emotional process, task, and stimulus domains differed substantially, as did the distribution by disorder. We observed strong evidence of a transdiagnostic and domain-general emotional neurocircuit disruption, with limited diagnosis-specific effects. The latter may reflect a lack of power due to the limited corpus of published papers for some disorders and/or the fact that ALE probes spatial convergence without accounting for individual effect sizes. Similarly, the cross-disorder aberrant activation analyses were inherently better powered than those specific to hyper- and hypoactivation. Thus, interpretations that directionality of neurocircuit disruption is not central to understanding psychiatric dysfunction are not warranted based on these findings. Additionally, polythetic diagnostic schemes, comorbidity, and the inherent difficulty in establishing consensus on principal disorder could hamper detection of emotional processing impairments more unique to a given disorder and instead bias the detection of common patterns. Any coordinate-based meta-analysis must rely on published findings, which in turn may entail publication biases that favor positive findings and those that are located in a priori hypothesized regions. However, the present whole-brain analysis should minimize these biases, and indeed, previous neuropsychiatric meta-analyses have provided negative results in spite of a potentially biased literature (
24,
63). Potentially influential factors such as medication types, illness duration, and comorbidity could not be assessed because of inconsistent reporting across studies. Further, our findings are most applicable to (younger) adults owing to there being few published studies in children and older adults. Additionally, it was not possible to assess effects of gender, as virtually all studies investigated more or less gender-balanced samples. On average, 51% of experimental samples were female and 49% male. As such, the results represent convergence in roughly equally mixed samples. Less than 8% of studies assessed a single sex, and most of those were studies of males with first-episode psychosis or schizophrenia. Lastly, while this is the most comprehensive meta-analysis of psychiatric disorders on emotional processing, the included studies do not represent the whole of the neuroimaging literature, including the vast number of studies focused on specific regions of interest.
Conclusions
In summary, we observed a common network of emotional disruption across disorders that largely mirrored the network(s) utilized in adaptive emotional expression and regulation (
18,
46). In particular, the identified abnormal pattern corresponds largely to the ventrostriatal/ventromedial prefrontal “reward” network, the lateral orbitofrontal “nonreward” network, and the salience network. In turn, the functions of these networks correspond to the proposed Research Domain Criteria dimensional neurobehavioral domains of positive valence, negative valence, and cognitive control (
64,
65). Taken together, these findings support the concept that psychiatric illness may be productively formulated as dysfunction in transdiagnostic neurobehavioral phenotypes, such as neurocircuit activation, as opposed to discrete diagnoses.
These findings underscore the fact that coordinated network disruptions, including but also distal to the amygdala, underlie observed impairment and may be productive targets for intervention. More specifically, these findings, in conjunction with our previous transdiagnostic gray matter reduction and cognitive control meta-analyses, highlight the right ventrolateral prefrontal/orbitofrontal cortex, anterior insula, and dorsal anterior cingulate as especially promising targets for interventions to remediate cognitive and affective dysfunctions common across mental illnesses.
Acknowledgments
Dr. McTeague was supported by NIMH grant K23 MH104849. Mr. Rosenberg was supported by National Science Foundation Graduate Research Fellowship under grant DGE-1650604. Dr. Huemer was supported by the Max Kade Fellowship provided by the Austrian Academy of Sciences. Dr. Eickhoff was supported by the Deutsche Forschungsgemeinschaft (DFG, EI 816/4-1; and LA 3071/3-1), NIMH (grant R01-MH074457), the Helmholtz Portfolio Theme “Supercomputing and Modeling for the Human Brain,” and the European Union Seventh Framework Programme (FP7/2007–2013) under grant agreement 604102. Dr. Etkin was supported by NIMH grant DP1 MH116506 and the Sierra-Pacific Mental Illness Research, Education, and Clinical Center at the VA Palo Alto Health Care System.