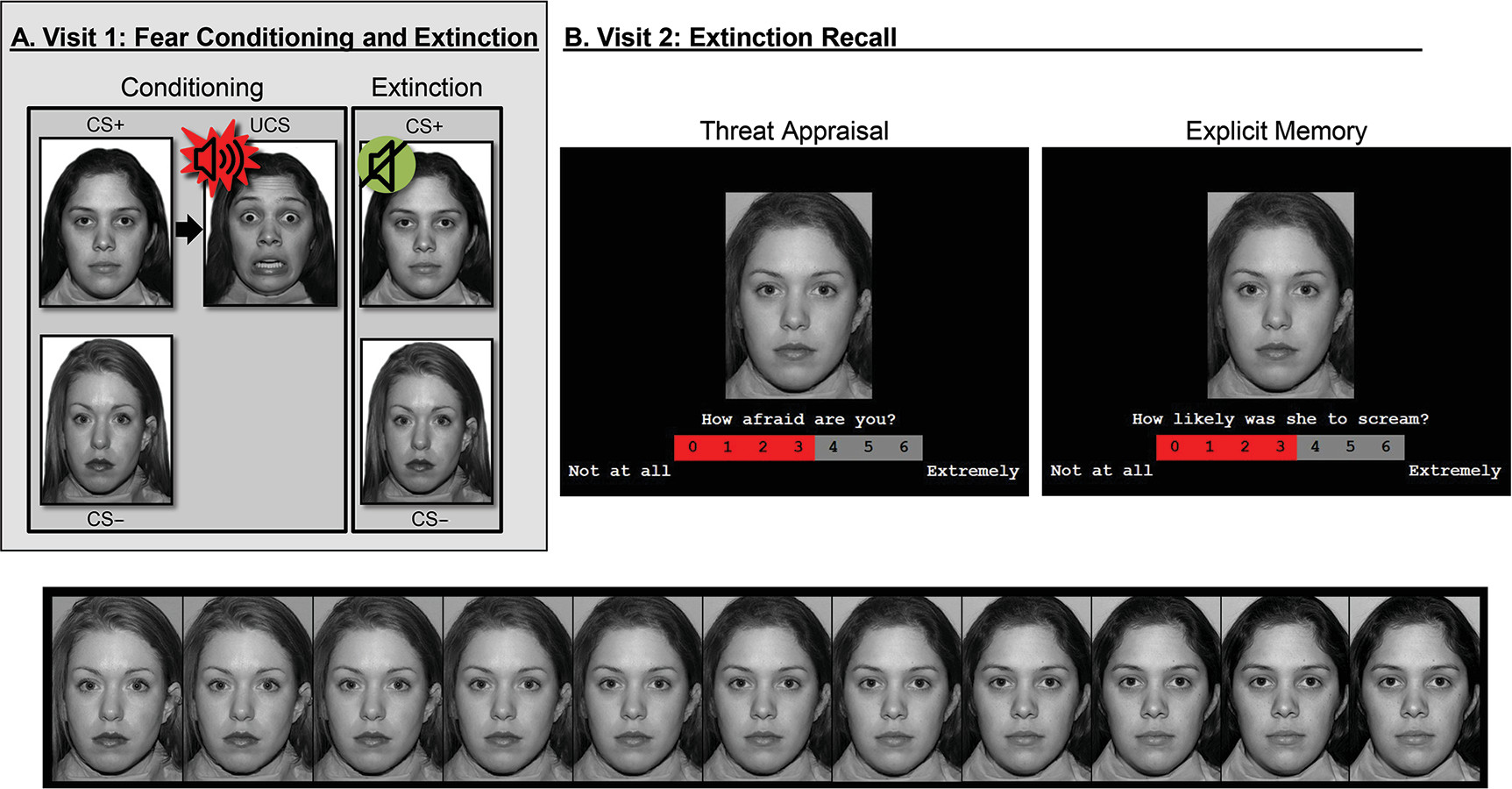
Threat conditioning provides a useful context for this research (
6). Threat conditioning occurs when neutral stimuli are paired with aversive unconditioned stimuli, leading the neutral stimulus to acquire the capacity to evoke defensive behavior (
7,
8). Developmentally and across species, threat conditioning research shows preadolescent maturation of some amygdala-related functions, including aspects of learning and attention (
9–
12). In youths with anxiety, previous studies have suggested that perturbation in such circuitry begins early in life and endures (
1,
2). In contrast, consistent with the late maturation of the prefrontal cortex (PFC), research has revealed differences between pediatric and adult anxiety disorders for PFC-supported functions, such as the sustained maintenance of extinction (
4,
5,
10–
13). In addition, late-maturing circuits support functions unique to primates, such as the efficient categorization of extinguished threats and other ambiguous cues (
14–
16). Thus, distinct amygdala-PFC circuitry dysfunctions may manifest in pediatric and adult anxiety disorders during categorization processes, such as subjective appraisal or declarative memory, triggered by ambiguous threat cues. To evaluate this possibility, fMRI paradigms can parametrically model ambiguity, using stimuli that blend features of extinguished threat and safety cues (
4,
17). Such paradigms are well suited (
18) for mapping age differences in the neural correlates of anxiety disorders.
Based on previous research, we hypothesized that fMRI response profiles would manifest differently in pediatric and adult anxiety disorders in late-maturing, recently evolved circuitry, encompassing the heteromodal association cortex (
16). We expected greater engagement of this circuitry in healthy, older individuals compared with similarly aged patients with anxiety disorders, thereby extending findings from previous studies on adult anxiety. We further hypothesized that functional connectivity between the amygdala and ventromedial prefrontal cortex (vmPFC) would differ between healthy adults and adults with anxiety, also extending findings from previous investigations (
5,
20,
21). Finally, limited previous data have generated less precise hypotheses for children than for adults, although the available data reveal different patterns in pediatric than adult anxiety (
4,
5). Thus, given sparse and inconsistent findings, support appears strongest for an overall expectation of differing patterns in pediatric and adult anxiety disorders, both for heteromodal association cortex activation and amygdala-vmPFC functional connectivity, with unclear expectations regarding diagnostic differences in youths.
Discussion
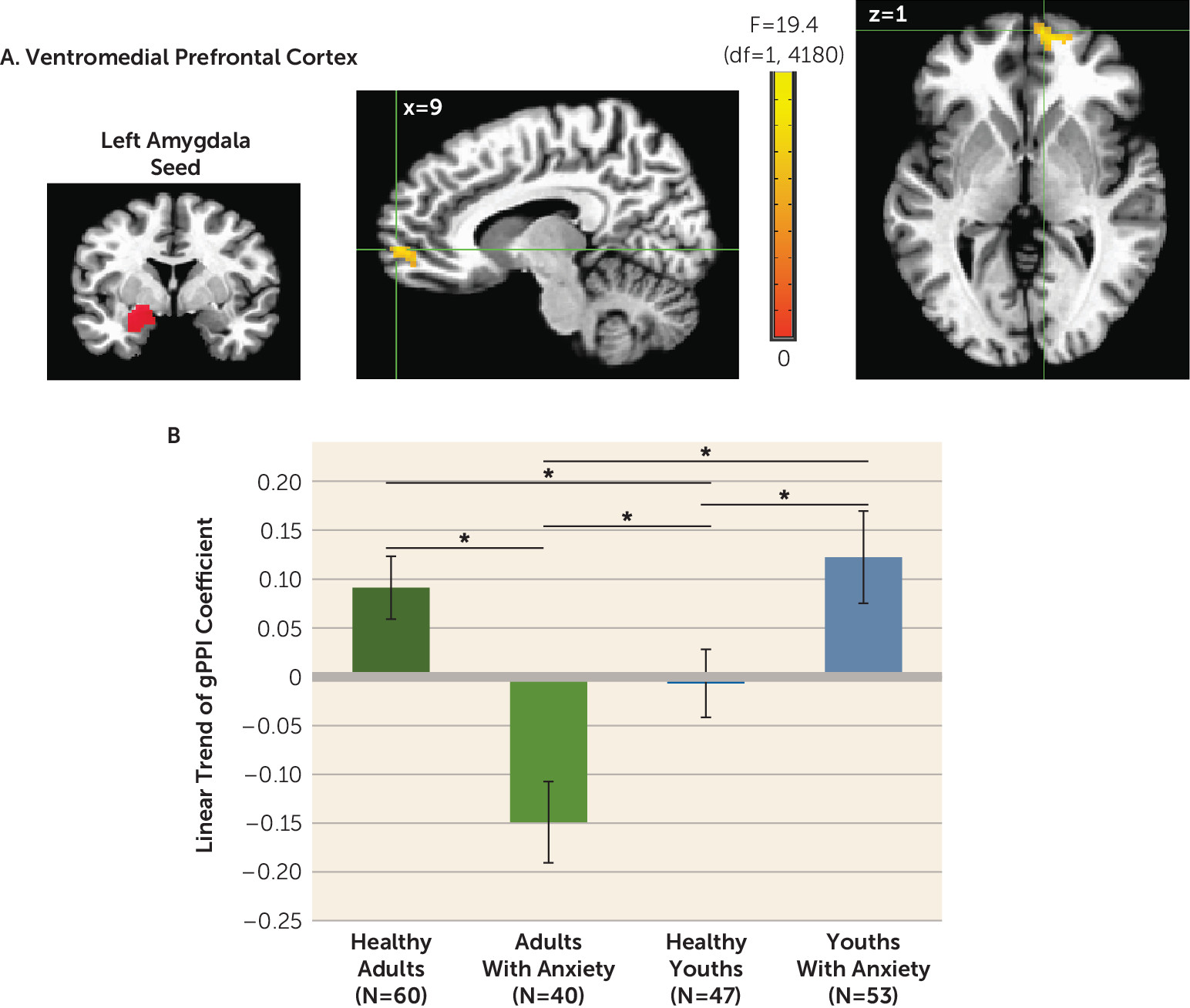
Two of our study results could influence neurodevelopmental theory. First, compared with adults with anxiety, healthy adults exhibited more positive amygdala-vmPFC connectivity as stimuli increasingly resembled safety cues, and the opposite pattern was observed in healthy youths compared with youths with anxiety. Second, age moderated anxiety-related differences in activation in ways that varied across attention conditions and brain regions. These implications extend beyond neurodevelopmental theory to highlight methodological complexities in fMRI research as well.
Considerable previous research employed face viewing tasks to examine amygdala and PFC functioning in typical development. In these studies, as rigor increased, notable age differences failed to replicate, including for PFC and valence-specific activation (
26,
27). However, findings for amygdala-PFC functional connectivity occurred with some consistency (
5,
21), possibly because of stronger reliability data for amygdala-PFC connectivity than valence-specific activation (
28,
29). Among healthy volunteers in the present study, adults but not youths exhibited increasingly positive amygdala-PFC coupling as levels of safety information increased in ambiguous face cues.
These data extend findings from previous research on amygdala-PFC development. Previous human studies demonstrated age differences in amygdala-PFC connectivity during face emotion viewing, whereas previous rodent studies linked age differences in amygdala-PFC function to age differences in threat learning (
12,
13). In the present study, we used conditioning and extinction with face cues depicting neutral expressions, which revealed age differences in amygdala-PFC connectivity during extinction recall. This connects previous face emotion imaging research in humans with threat-learning research in rodents, all revealing age differences in amygdala-PFC functioning.
Functional connectivity results also extend findings from previous research on anxiety disorders, particularly in adults. In this age group, we found lower amygdala-vmPFC connectivity in participants with anxiety compared with healthy subjects, particularly to face cues containing high degrees of safety-related information. These findings were consistent with theory suggesting that anxiety disorders involve deficient safety cue signaling (
20,
30). Such theory implied that failure to appropriately maintain safety cue representations in experimental settings also manifested in the everyday lives of individuals with anxiety, where they experience fear when failing to recognize safety information. Notably, such theory highlighted experimental data for adults. In the present study, compared with adults, data for youths demonstrated opposite trends, with greater amygdala-vmPFC connectivity in participants with anxiety compared with healthy subjects.
In disorder subgroups, such as pediatric and adult anxiety disorders, such opposite-appearing trends are termed crossover interactions. Compared with a crossover pattern, interactions more frequently involve disorder-related differences in only one of two subgroups. Relative to our findings on connectivity in youths with anxiety, our data for adults with anxiety more closely replicate findings from previous research. Thus, these new connectivity findings for youths should be viewed with caution and require replication, given our observation of crossover patterns. Recent neurobiological studies have, however, demonstrated crossover interactions in pediatric anxiety with increasing frequency (
31,
32), including longitudinal studies showing age-related crossover interaction in neural correlates in pediatric anxiety (
32,
33). Finally, it is notable that connectivity profiles in this study differed among younger and older patients and healthy participants. Thus, replication is needed in medication-free youths and adults both with and without acute anxiety.
Although we found age to moderate both connectivity and activation, the patterns differed. Whereas connectivity findings related to cue features, activation findings related to viewing contexts. Moreover, unlike connectivity findings manifesting as a crossover interaction, interactions for activation resembled more typical findings in previous research (i.e., associations in only one age-delimited subgroup).
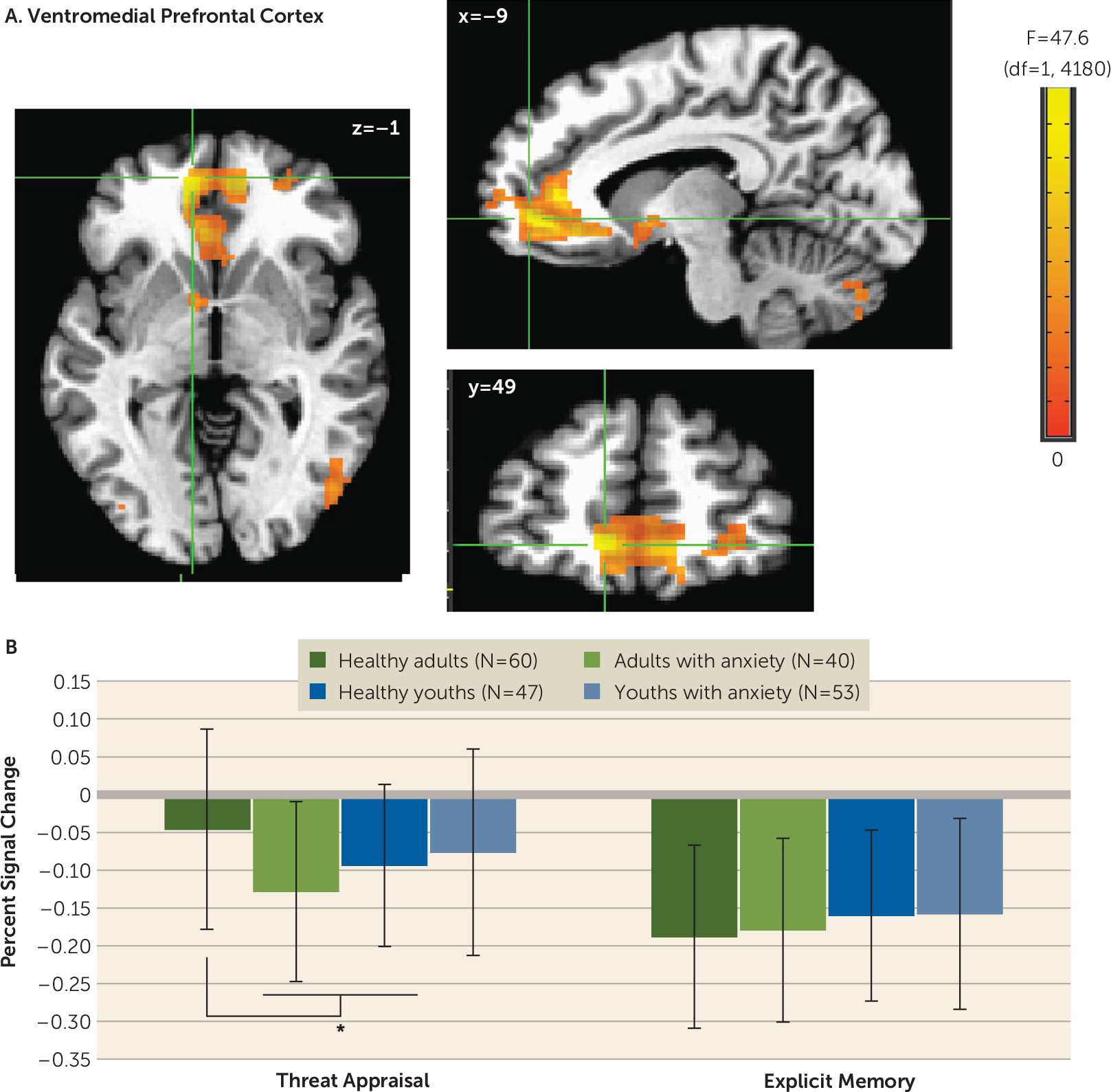
Differences in vmPFC activation manifested in adults with anxiety but not youths with anxiety. These findings resemble patterns in previous studies of emotional disorders, in which group differences involved mPFC deactivation (
20,
34,
35). In such studies, less deactivation or greater activation in healthy adult volunteers compared with adult patients may reflect default-mode dysfunction, which can generate apparent task deactivation through group differences in baseline conditions (
36,
37).
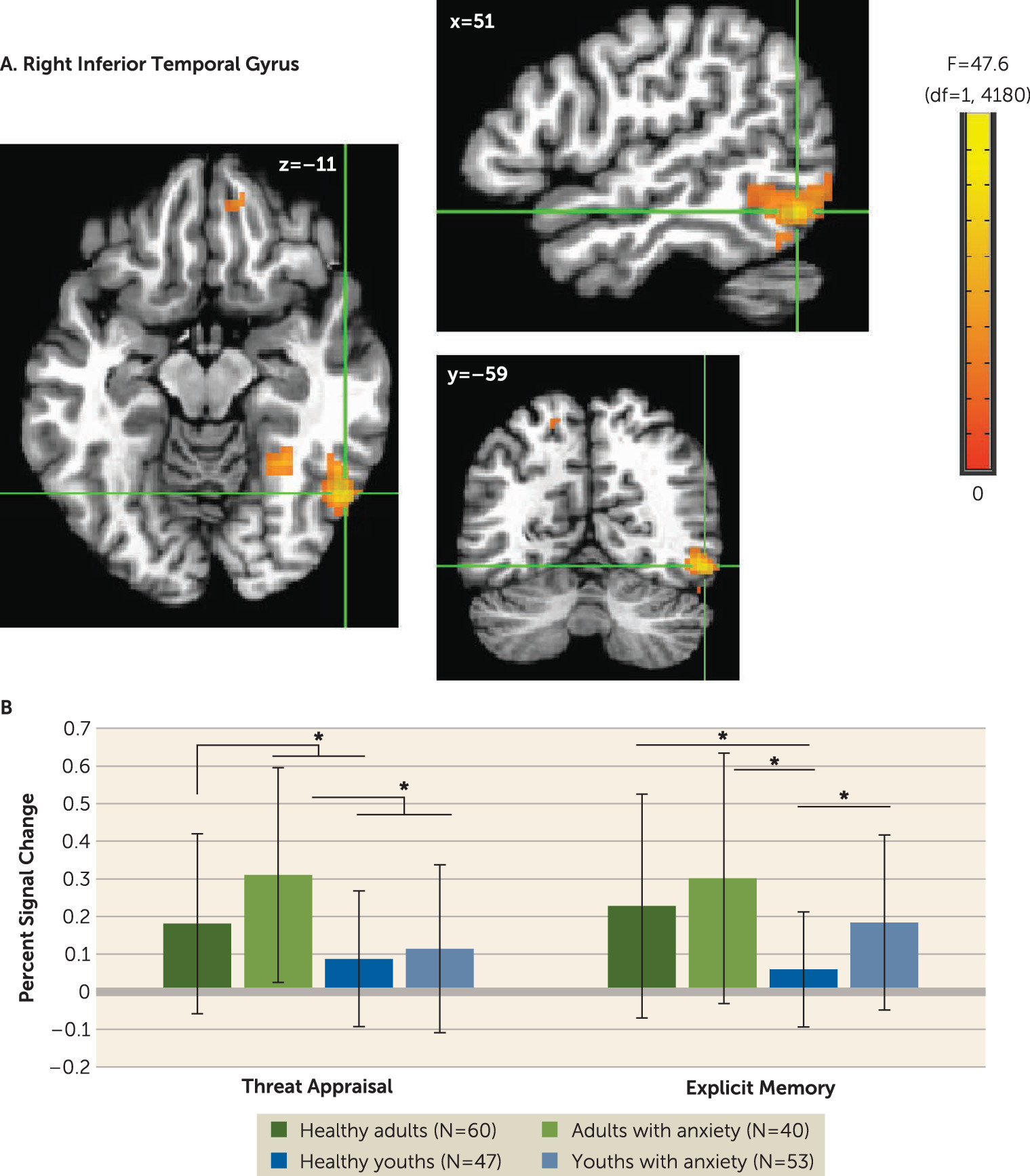
In our study, group differences in inferior temporal activation manifested with different patterns in youths with anxiety compared with adults with anxiety, as a function of task instructions. These differences occurred in the context of cue-related activation in all participants, replicating studies of temporal responses to faces. Patterns among youths resembled patterns found in another study, which showed group differences during memory tasks (
19), and which extended ample work on developmental differences in face representation within the inferior temporal cortex (
26,
27,
38).
Given the novelty of these developmental findings, replication is needed before their significance can be determined. However, these results do provide clues regarding processes underlying anxiety disorder development. For example, the observed mechanisms mediating age differences in brain activation could arise from earlier maturation in circuitry supporting children’s capacity to remember events rather than to appraise their emotional impact. Emerging but immature memory circuit function may create vulnerabilities reflected in our results, including more prominent findings in youths than adults on the threat memory task. Earlier maturation may arise for memory rather than appraisal as a result of greater objectivity in the process of memory encoding. In contrast to subjective fear judgments, the verifiable nature of events encoded in memory may create opportunities for social scaffolding. In this scenario, children’s and adolescents’ discussions of their memory for threatening events with other people witnessing the events could sculpt their threat-related memory circuitry. Such mechanistic neurodevelopmental hypotheses may guide clinical research on anxiety disorders.
For activation, the main analysis compared groups at average reaction time in order to generate reaction time-adjusted findings. To a degree, this accounts for influences of reaction time on activation. Notably, these analyses revealed no four-way interactions, possibly reflecting reduced statistical power in tests of higher-order interactions. Alternatively, previous research suggested that cross-group heterogeneity in brain physiology coupling can obscure findings in higher-order interactions (
16,
19). To address this problem and thereby enhance sensitivity to higher-order interactions, one study on adolescent social reticence used subject-specific regressors for skin conductance responses collected during extinction recall (
19). Another study on typical adolescent development used ratings during threat conditioning (
16). Although we did not measure skin conductance response in our study, analyses presented in the
online supplement applied the approach from these previous studies using subject-specific and event-specific reaction time regressors. Future work might measure both reaction time and skin conductance responses while examining higher-order interactions and relations among activation, skin conductance response, and reaction time.
Beyond extending previous fMRI data, our findings carry more general implications. Comparing brain function in youths and adults broadly informs developmental perspectives. Other research on anxiety disorders has shown developmental continuities (
39–
42), often for processes that basic research localizes to early-maturing circuits with strong cross-species conservation. In contrast, our results suggest that age-related discontinuities exist in the neural correlates of anxiety disorders. Age moderated anxiety-related differences on tasks that engage late-maturing capacities, supported by areas of the heteromodal association cortex unique to primates. Thus, longitudinal work may consider whether developmental changes in the neural architecture of these capacities accounts for age-related discontinuities in anxiety disorders.
Our findings also broadly extend perspectives relating normative and pathological development. Structural imaging has shown maturation throughout adolescence in the heteromodal association cortex (
43,
44). Moreover, children’s capacity to appraise their internal state and perform memory tasks matures on similar time scales, consistent with changes in brain regions engaged by tasks engaging these capacities (
4,
5,
45,
46). Our study extends this research in healthy youths to youths with anxiety in our observation of age differences in late-maturing cortex on tasks that engage late-maturing psychological capacities.
Longitudinal research could evaluate the clinical significance of our findings by connecting neural and clinical data. Such research may find that youths with anxiety who develop relatively strong appraisal or memory capacities overcome aspects of their anxiety disorder. Conversely, such research may find that healthy youths who do not develop these capacities have a high risk for adult anxiety. If such longitudinal findings do emerge, prospective studies might use the associated insights to locate biomarkers that identify youths with anxiety at risk for persistent emotional problems or healthy youths at risk for new ones.
Beyond such clinical extensions, other studies could examine pathophysiology. Longitudinal genetic research has shown anxiety to involve complex developmental processes (
47). Some processes are age invariant, while others are age specific. In tandem with previous imaging research, our data suggest that neural correlates may reflect similarly complex multicomponent processes. Thus, as in genetic studies, prospective imaging studies might seek to differentiate age-invariant from age-specific correlates in pediatric and adult anxiety disorders.
Strengths of this study include a relatively large sample and observation of clinically relevant findings. However, these findings should be considered in the context of four limitations. First, cross-sectional data only provide initial insights on development, as findings may guide longitudinal research. Second, while patients were medication-free and had acute anxiety, there were group differences (i.e., the gender ratio in adults). Only pediatric patients were required to enter a treatment study, and more youths than adults asked whether they would be exposed to the unconditioned stimulus during imaging. Results remained unchanged after statistically accounting for these factors. Third, evoked levels of fear generally were mild, with mean levels in the lower half of a 0–6 rating scale. Future research might use more evocative stimuli. Lastly, although this study was large in the context of clinical imaging, the sample size may still be small, because we examined complex questions with correspondingly complex methods. Statistical thresholds were more conservative than those in other studies, but few effect sizes were large, some interactions reflected unexpected patterns, and larger samples would support more powerful tests and more conservative thresholds. In addition, simpler designs would generate more powerful tests but also could lack sensitivity to multiplex clinical features, such as complex relations among task conditions, diagnosis, and age. Replication is important, given the complexity of interactions. Beyond addressing limitations with cross-sectional data, such replications might examine very large samples to appropriately power conservative statistical tests of complex interactions, with expectations of moderate effect sizes.
In conclusion, discontinuities exist between pediatric and adult anxiety disorders. Our findings suggest that differences in brain function manifest on measures of amygdala-vmPFC functional connectivity and task-related activation in the heteromodal association cortex. Such differences occur when viewing ambiguous, extinguished threat cues and engaging late-maturing psychological capacities. These results could shape refinements in developmental perspectives on anxiety disorders.