Disruptive behavior disorders (DBDs), including oppositional defiant disorder and conduct disorder, are prevalent childhood psychiatric disorders and the most common reason for childhood mental health referrals (
1). DBDs are linked to a host of maladaptive social, emotional, and behavioral outcomes across the lifespan, and youths with DBDs incur significant financial costs through greater use of educational, criminal justice, and medical services (
2). DBDs are characterized by aggression and social dominance in the pursuit of rewards, often at the expense of others’ welfare (
3). An improved understanding of the motivational processes underpinning DBDs is crucial to the development of more effective treatments, including pharmacotherapeutic approaches and the incentivization of behavior change with psychotherapy.
An important focus for etiological research is the reward-related neurocircuitry underlying DBDs, including the highly interconnected brain regions of the mesolimbic dopamine system. A large body of research exploring reward processing has focused on the ventral striatum and nucleus accumbens (NAcc) (
4) as principal targets of dopamine neurons (
4). NAcc recruitment arises from the combination of emotion-related activation and instrumental motor-preparatory responsivity (
4,
5). Instrumental behavior is supported by NAcc connectivity with frontal brain regions, including the orbitofrontal cortex (OFC), which evaluates reward value and probability, the ventral medial prefrontal cortex (vmPFC), which monitors correct responses based on previous experience, and the dorsal anterior cingulate cortex (dACC), which monitors conflict (
5). In addition, the insula and thalamus increase arousal and promote communication between the ventral striatum and vmPFC, while the amygdala is primarily responsible for processing motivationally salient stimuli (
6).
Importantly, research has distinguished between neurocircuitry underpinning the willingness to expend effort (reward anticipation) and hedonic processing (reward receipt) (
7). During the anticipatory stage, reward-predicting cues elicit phasic release of striatal dopamine, leading to the urge to obtain and consume the reward (
7), whereas reward receipt is characterized by hedonic properties, such as feelings of pleasure. Temporal difference models of reinforcement learning (
8) posit that in the absence of a predictive cue, dopamine neurons respond to receipt of an unexpected reward (i.e., a positive prediction error); however, once an association between a predictive cue and reinforcer is learned, there is a shift in the dopaminergic response to the presentation of the cue associated with subsequent reward receipt (
7,
9). Thus, a central function of reward processing is to allow for adaptable and flexible responding to maximize reward gain.
Notably, youths with DBDs exhibit difficulties linking outcomes with antecedent events, consistent with their reported deficits in cognitive flexibility and behavioral adaptation (
3). Some of this inflexibility could arise because these youths show robust behavioral responses to receiving various rewards (
3), prefer large, immediate rewards (
10), and demonstrate more aggression in response to frustrative nonreward (
11). Concomitantly, youths with DBDs show impairments in the use of changing expected value information to optimize behavior (
12,
13), as well as reduced anticipatory recruitment of motivational neurocircuitry (
14). These findings suggest that youths with DBDs may have an exaggerated neural response during the emotional component of reward receipt coupled with minimal recruitment of neurocircuitry required for the effort to obtain it. However, few functional MRI (fMRI) studies have examined any such divergence in the motivational neurocircuitry underpinning DBDs. Two recent meta-analyses (
15,
16) retrieved only seven fMRI studies of reward-related neural processing in youths with DBDs, and only three studies distinguished between reward anticipation and receipt. Moreover, findings were equivocal, with evidence that DBDs are associated with both hypoactivity and hyperactivity in various regions, including the NAcc, amygdala, ACC, and OFC (
3).
One possible explanation for these mixed findings is the heterogeneity in emotional reactivity among youths with DBDs, which could obfuscate valuation-based differences in reward-related neural activation. A sizable subgroup of youths with DBDs have callous-unemotional traits (DBD+CU), defined by a lack of empathy, remorse, and guilt (
3,
17). These youths display a reward-dominant response style compared with typically developing youths and more severe and persistent antisocial behavior across development (
3,
18). Emerging evidence suggests that youths with DBD+CU show aberrant functioning and disrupted connectivity of the amygdala and OFC during stimulus-reinforcement learning, incentive motivation, and moral learning (
19). However, to our knowledge, only a single study has examined associations between callous-unemotional traits and distinct phases of reward processing, and it found reduced activation in the medial prefrontal cortex during reward anticipation (
20). The sparse and mixed body of available research underscores the need for more research in this area.
Three other gaps exist in our knowledge of reward-related neural functioning and DBDs. First, previous studies have typically assessed samples of fewer than 20 youths with DBDs, rendering them potentially underpowered to detect differences. Second, although DBD often emerges during childhood, previous studies have focused primarily on high-risk, male adolescents, a population for which there is a risk that substance use may confound reward-related neurodevelopmental abnormalities. Thus, larger samples of boys and girls are needed in studies that are conducted before the transition to adolescence. Third, although multiple regions of interest have been individually linked to reward-related neural processing differences in DBDs, these regions operate within an overarching reward network (
5). Understanding whether coactivation across a higher-order network or unique region-specific activation differentially contributes to brain-behavior associations may offer greater insight into the neuroetiology of DBDs.
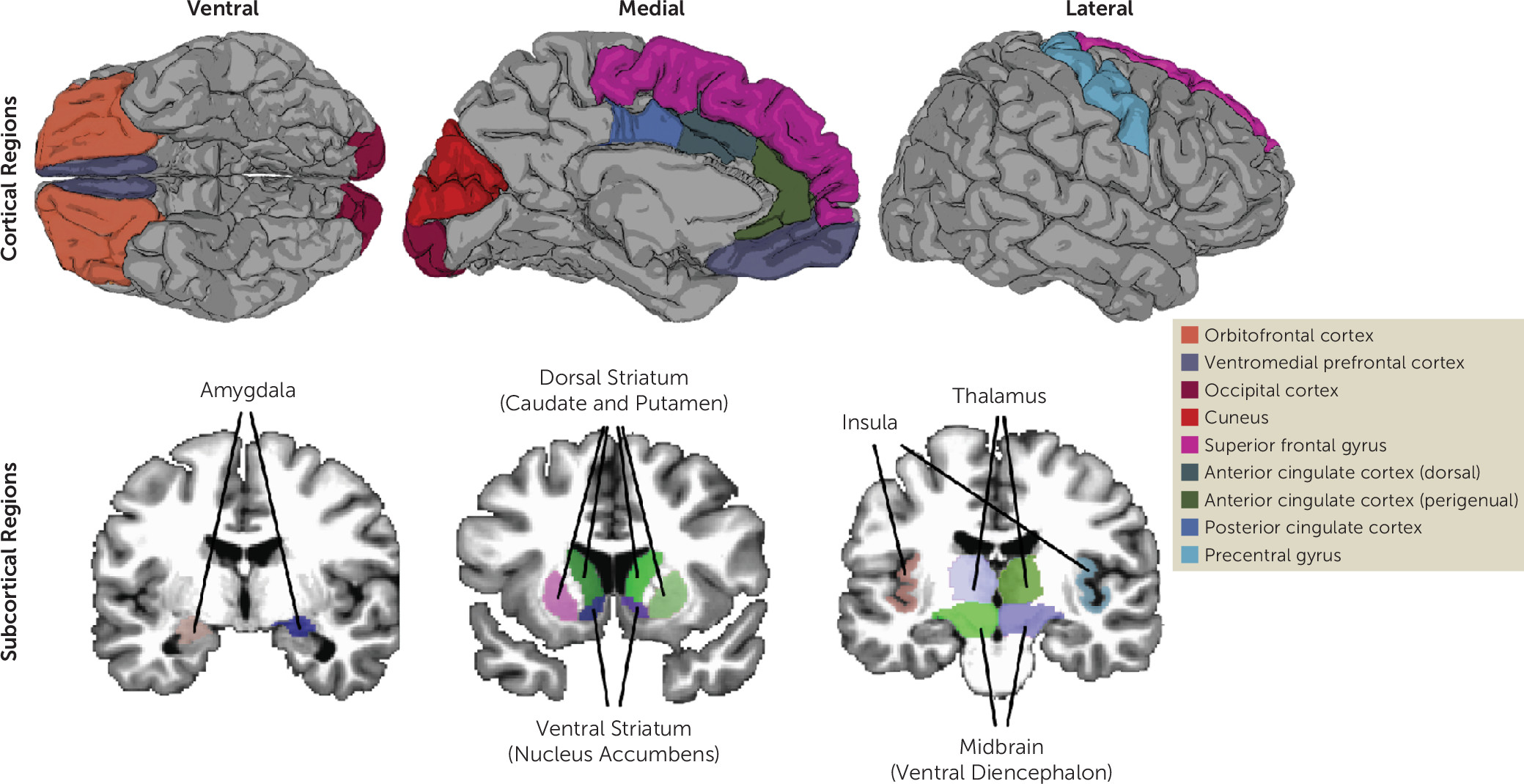
Using the full baseline data release from the Adolescent Brain and Cognitive Development (ABCD) Study (N=11,874; mean age=9.51 years [SD=0.50]) in the largest DBD-focused fMRI study to date, we tested for group-based differences in reward-related brain activation between the following groups: youths with DBDs compared with typically developing youths, youths with DBD only compared with typically developing youths, youths with DBD+CU compared with typically developing youths, and youths with DBD only compared with youths with DBD+CU. In addition, we tested whether sex moderated these associations. We also used latent variable modeling to examine network-level reward-related activation using a latent reward network factor. We focused on 16 cortical and subcortical regions (
Figure 1; see also Table S1 in the
online supplement) consistently identified in fMRI meta-analyses of reward processing and DBD (
15,
16,
21). We hypothesized that compared with typically developing youths, youths with DBD (irrespective of callous-unemotional traits) would exhibit reduced anticipatory reward activation and increased activation to reward receipt. Because of the limited number of previous studies, and the likely similar utility of monetary rewards among boys and girls, we made no a priori hypotheses about sex differences. Finally, we hypothesized that regions of interest would form a well-fitting higher-order latent reward network factor during reward anticipation and receipt similarly across groups.
Methods
Participants
Participants were enrolled in the ongoing, longitudinal ABCD Study, and the study data were from the annual 2.0 data release (
https://data-archive.nimh.nih.gov/abcd). The ABCD Study recruited 11,874 healthy children, ages 9–10, to be followed into early adulthood. Participants across 21 study sites were recruited through public and private elementary schools, with sampling approaches intended to yield a final sample that approximated national sociodemographic characteristics. Institutional review boards at participating universities approved all study procedures. Participants provided written assent, and their legal guardians written consent, for participation.
Measures
DBDs.
DBDs were classified using the Child Behavior Checklist (CBCL) (
22) and the Schedule for Affective Disorders and Schizophrenia for School-Age Children–Present and Lifetime version for DSM-5 (K-SADS-PL) (
23,
24). Parents completed the self-administered computerized version of each of these measures.
Callous-unemotional traits.
We classified callous-unemotional traits using a four-item measure derived and validated in previous studies using data from the ABCD Study (
25,
26) that included one item from the CBCL–Parent Report (“lack of guilt after misbehaving”) and three (reverse-scored) items from the Strengths and Difficulties Questionnaire–Parent Report (
27) (e.g., “is helpful if someone is hurt or upset”). This measure showed strong psychometric properties, including discriminant (e.g., crossover suppression effect between callous-unemotional traits and anxiety; see also Table S2 in the
online supplement) and convergent validity, measurement invariance, independent replication, and overlap with other commonly used measures of callous-unemotional traits (
25). Finally, we also derived maximum a posteriori scale scores that provided person-specific callous-unemotional trait factor scores (see the
online supplement).
Group classification.
To attain a robust sample size to explore DBD groups, we inclusively classified the presence of DBDs on the basis of youths scoring at or above the borderline clinical range (i.e., T-scores ≥67) on either the CBCL DSM-oriented conduct problems subscale or oppositional defiant problems subscale or receiving a K-SADS-PL conduct disorder or oppositional defiant disorder diagnosis (N
=995). Youths with DBDs were further categorized on the basis of extremes of callous-unemotional traits. The DBD+CU group had high callous-unemotional traits based on a conjunction of summed scores ≥4 on the summed callous-unemotional traits measure and callous-unemotional maximum a posteriori scores in the 90th percentile (DBD+CU traits, N
=198). To maximize phenotypic differences between groups, youths in the DBD-only group had summed scores of zero on the callous-unemotional traits measure (N=276). Typically developing youths were those with T-scores equaling 50 across all CBCL scales and summed scores of zero for callous-unemotional traits (N=693) (
Table 1). Additional analyses using alternative grouping approaches (i.e., both more and less stringent) and covariate sets are presented in Tables S3–S8 and S16 in the
online supplement.
Potential confounders.
To account for the potentially confounding effects of attentional, cognitive, or emotional difficulties that are often comorbid with DBDs, all analyses included CBCL DSM-oriented attention problems and internalizing subscale scores as covariates. We also covaried for sex, age, race/ethnicity, and parental education.
Imaging Measures
Monetary incentive delay task.
A version of the monetary incentive delay task was used to measure brain activation during anticipation and receipt of monetary rewards (
28), which had three trial types: win money ($0.20 or $5), lose money (−$0.20 or −$5), or no incentive ($0). Participants saw a cue (pink circle, yellow square, or blue triangle) at the beginning of trials that indicated the valence (win, loss, or no incentive) and the amount of money at stake ($0, $0.20, or $5). This cue presentation (2,000 ms) was followed by a jittered anticipatory delay (1,500–4,000 ms). A black target shape (same shape as the previously presented cue) was then shown, and participants could gain money or avoid losing money by pressing a response button while the target shape was on the screen. The time the target was on the screen was dynamically manipulated to maintain a 60% success rate, and participants could not hit the shape every time. After a short response window, feedback was provided (2,000 ms). During the anticipation phase, participants received 40 reward and loss trials and 20 neutral trials. During the outcome phase, the adaptive algorithm resulted in 24 positive feedback trials (for both reward and loss) and 16 negative feedback trials (for both reward and loss) on average (
28). Participants completed two runs of the task (approximately 5.5 minutes each).
Image preprocessing and calculation of region-of-interest data.
The ABCD Data Analysis and Informatics Center performed centralized processing and analysis of MRI data, leveraging validated methods used in other large-scale studies (
28,
29) (see also the Methods section in the
online supplement). Briefly, parcellated cortical and subcortical regions used in study analyses were derived from cortical surface reconstruction and subcortical segmentation performed using FreeSurfer, version 5.3.0 (
30). Estimates of canonical task-related activation were computed at the individual subject level using a general linear model implemented in AFNI’s 3dDeconvolve and were released as contrast beta weights (
31). We focused on two contrasts: anticipation of large reward versus no incentive and reward receipt versus missed reward. For these contrasts, average general linear model beta coefficients were computed during each of two runs and then averaged across runs (
28,
29) (see also the Methods section in the
online supplement).
Analytic Strategy
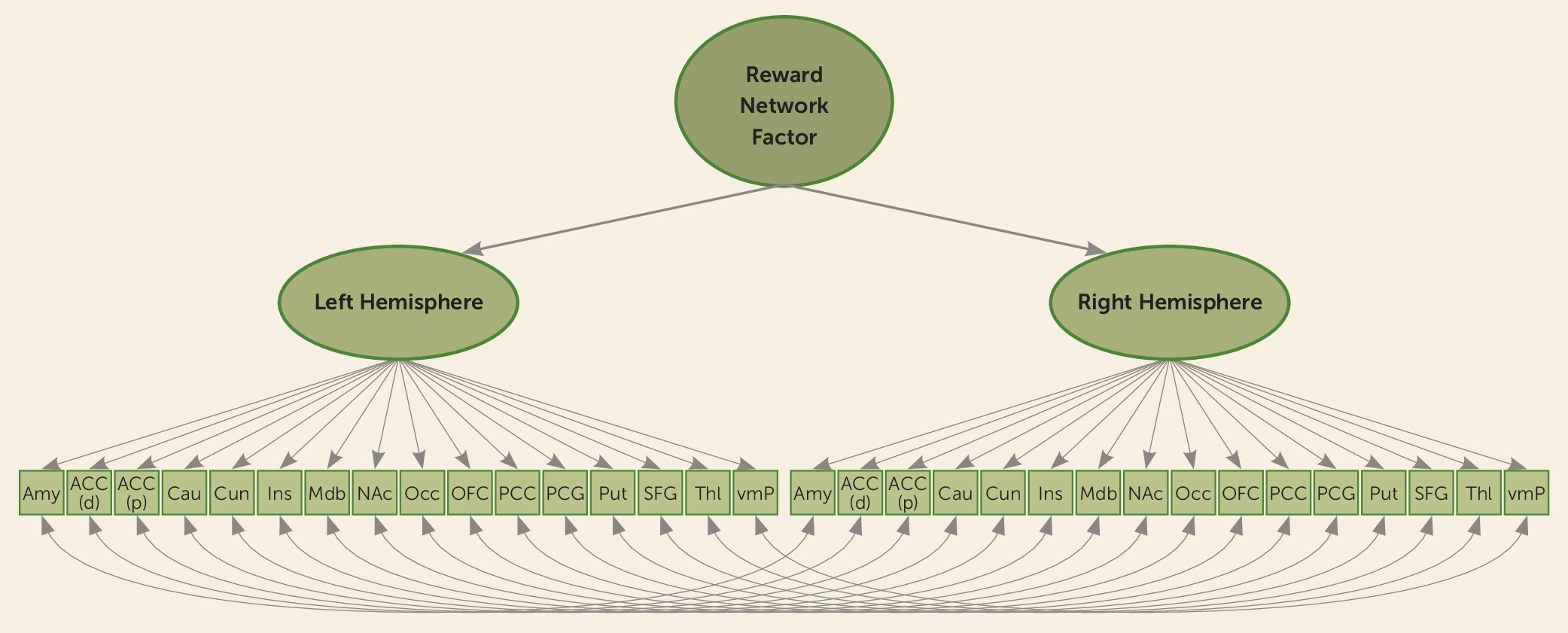
We used multinomial logistic regression to examine differences in reward-related brain activation in regions of interest between the following groups: youths with DBDs compared with typically developing youths, youths with DBD only compared with typically developing youths, youths with DBD+CU compared with typically developing youths, and youths with DBD only compared with youths with DBD+CU. We also examined whether sex moderated these associations. Next, we used latent variable modeling to determine whether the spatial pattern of coactivation across all 16 reward-relevant regions (i.e., relative contribution of each region to a broader reward network) differed between groups during reward anticipation and receipt. We specified a second-order factor model in which each (separably lateralized) region of interest loaded onto a right or left reward factor. These first-order right and left lateralized factors were then used as indicators of a higher-order factor, representing an overarching latent reward network factor (
Figure 2). The residuals of homologous region-of-interest pairs were specified to correlate (e.g., the left OFC with the right OFC).
The reward network factor model allowed us to examine group differences in activation at a network level, supplementing the region-of-interest analyses in several ways. First, we could evaluate how each region contributed to the hypothesized reward network and assess whether the network could be modeled similarly across groups. We used measurement invariance testing (via the DIFFTEST procedure in Mplus, version 7 [
32]) to evaluate the reward network factor model across groups (i.e., configural invariance) and whether each region contributed to the higher-order network equivalently across groups (i.e., metric and scalar invariance) (see the Methods section in the
online supplement). Second, our modeling approach allowed us to test for group differences in mean levels of activation at the network level (i.e., not just individual regions), which was made more advantageous given that latent variables are modeled free of measurement error, resulting in increased statistical power.
All analyses were conducted using Mplus, version 7 (
32), with models specified using maximum likelihood estimation with robust standard errors and a Monte Carlo numerical integration algorithm. Complex sampling and recruitment procedures for the ABCD Study were accounted for using cluster correction (i.e., for sibling pairs) and stratification sampling (i.e. study site) procedures (
32). The median absolute deviation was used to identify study outliers, with values winsorized at ±2.5 times the median absolute deviation. Outlier cases did not exceed 5% for any region.
Results
Descriptive statistics revealed that the study groups were equivalent on demographic variables, except for lower proportional representation of female and Hispanic children in the DBD groups (
Table 1). In addition, there were no differences across groups on monetary incentive delay task behavioral performance (e.g., mean reaction time and total earnings) (see Table S10 in the
online supplement).
Reward Anticipation in Individual Regions of Interest
Youths in the overall DBD group demonstrated decreased activity in the left dACC compared with typically developing youths (
Table 2). In the DBD subgroups, youths in the DBD-only and DBD+CU groups exhibited reduced dACC activation compared with typically developing youths, although they did not differ from each other. Compared with typically developing youths and youths with DBD+CU, the DBD-only group exhibited reduced activation in the NAcc, caudate, and putamen. In addition, the DBD-only group showed decreased activation in the left midbrain compared with typically developing youths. Finally, the DBD-only group showed reduced activity in the left and right cuneus, left and right occipital cortex, left and right precentral gyrus, left thalamus, right insula, right superior frontal gyrus, and right posterior cingulate cortex (PCC) compared with the DBD+CU group. Sex did not moderate any associations.
Reward Receipt in Individual Regions of Interest
Compared with typically developing youths, those in the overall DBD group demonstrated greater activation in the left and right perigenual ACC, amygdala, NAcc, and occipital cortex, as well as the left thalamus, left vmPFC, right dACC, right midbrain, and right orbitofrontal cortex (
Table 2). In the DBD subgroups, both the DBD-only and DBD+CU groups exhibited increased activation in the left and right NAcc and left and right OFC compared with typically developing youths, but they did not differ from each other. Additionally, the DBD-only and DBD+CU groups showed greater activation in the dACC compared with typically developing youths. The DBD+CU group also exhibited greater activation in the dACC compared with youths in the DBD-only group. Finally, youths in the DBD+CU group demonstrated increased activity in the right amygdala and left thalamus compared with typically developing youths, along with increased activation in the left and right dACC, right PCC, and right precentral gyrus compared with the DBD-only group. Sex did not moderate any associations.
Network-Level Responses Using Latent Reward Network Factor Models
Reward anticipation network.
Model fit was in the acceptable range for each group (see Table S11 in the online supplement), suggesting that a reward network factor model was present across groups. Multigroup invariance testing also revealed no significant differences in model fit (see Table S12 in the online supplement), suggesting that region-of-interest activation contributed to the higher-order reward network factor equivalently across groups. Comparing mean levels of reward network factor activation across groups revealed reduced mean-level reward network factor anticipatory activation in the DBD-only group compared with the DBD+CU and typically developing groups. There were no differences in mean-level reward network factor anticipatory activation between the DBD+CU group and the typically developing group (see Table S13 in the online supplement).
Reward receipt network.
Model fit was in the acceptable range for each group (see Table S11 in the online supplement), providing evidence to support a reward network factor model of reward receipt across groups. Multigroup invariance testing revealed no significant differences in model fit (see Table S12 in the online supplement), indicating that region-of-interest activation contributed to the higher-order reward network factor equivalently across groups. Examination of mean-level differences in activation revealed an increased response in the left hemisphere of the reward network factor model for the DBD+CU group compared with the typically developing group. No additional differences in mean-level reward network factor activation were found between groups (see Table S13 in the online supplement).
Discussion
We examined neural response to reward anticipation and receipt during a critical developmental window in late childhood, leveraging the ABCD Study to derive phenotypically narrow groups in the largest DBD-focused fMRI study to date. Consistent with hypotheses and previous reports (
33), youths with DBDs exhibited decreased frontostriatal activation during reward anticipation. In particular, youths with DBDs (regardless of callous-unemotional traits) demonstrated reduced activation in the dACC compared with typically developing youths when anticipating rewards. The dACC is a key node of the salience network that is strongly connected with the prefrontal cortex, parietal cortex, and striatum and underpins an array of cognitive and motor control functions (
34). Specifically, the dACC integrates reward-related target detection with value encoding to facilitate attentional allocation and guide behavior (
34). A decrement in response-anticipatory dACC recruitment is consistent with youths with DBDs exhibiting limited mobilization of attentional resources, commensurate with their generally lower executive function irrespective of callous-unemotional traits (
11). Indeed, relatively reduced activation of the dACC during periods of evaluation and decision making has been linked with cognitive-control deficits and risky choice selection among youths (
35).
Youths in the DBD-only group showed a specific pattern of reduced anticipatory reward activation in the ventral striatum and NAcc compared with typically developing youths and youths with DBD+CU. Anticipatory ventral striatum reward activation has been linked to expected reward value and underlies motivation-dependent positive arousal in response to future reward (
4). The relatively reduced reward-related ventral striatum activity in the DBD-only group is consistent with findings from one previous study, which suggested that striatal hypoactivity during reward anticipation may represent a general risk marker of DBDs but not callous-unemotional traits (
33). Youths in the DBD-only group also showed reduced anticipatory reward activation in the dorsal striatum, encompassing the caudate and putamen. The dorsal striatum is strongly interconnected with motor effector regions and contributes to decision making through its role in integrating sensorimotor, cognitive, and motivational information (
5). The relatively reduced dorsal striatum activation we identified in the DBD-only group could, along with reduced dACC recruitment, represent further evidence of a network-wide decrement in effort mobilization. Moreover, this decreased activation could give rise to poor decision making in the context of changing contingencies, producing the characteristic features of DBDs, including continued aggression or rule-breaking even when such behavior is no longer rewarded. In support of this interpretation, previous studies have shown that youths with DBD without callous-unemotional traits exhibit impaired and riskier decision making not only compared with typically developing youths but also compared with youths with high callous-unemotional traits (
12).
We found that youths with DBDs (irrespective of callous-unemotional traits) were characterized by heightened neural activation in response to reward receipt across various cortical (e.g., the ACC, OFC, and vmPFC) and subcortical (e.g., the amygdala, midbrain, NAcc, and thalamus) regions. These findings align with research showing that youths with DBDs exhibit strong behavioral responses to reward receipt (
3) and with previous neuroimaging findings linking externalizing psychopathology to exaggerated frontostriatal activity in response to reward outcomes (
36). During reward receipt, activation in the OFC and vmPFC represents the value of the received reward, outputting this information to the ventral striatum, which codes for hedonic value and prediction errors (
5,
9). The greater activation evident across these regions suggests that youths with DBDs experience heightened pleasure from receiving rewards compared with typically developing youths, leading to a reward-orientated behavioral style. A psychological manifestation of this neurocircuit abnormality is trait impulsivity, which is known to characterize youths with DBDs (irrespective of callous-unemotional traits) and is robustly related to heightened ventral striatum activation during reward receipt (
36).
We found several differences in neural activation during reward receipt that were specific to youths in the DBD+CU group. First, these youths demonstrated increased dACC activity compared with both the DBD-only and typically developing groups and greater PCC activity compared with the DBD-only group. The dACC exhibits a strong response after actions that lead to a reward (
5,
34), while PCC activity is associated with positive self-appraisal following reward receipt consistent with its role in self-referential neurocircuitry (
21). Our findings thus offer a potential neural mechanism underpinning the behavioral features of youths with DBD+CU, who exhibit an increase in positive emotions in response to obtaining a reward (
37). Second, the DBD+CU group showed increased right amygdala activation during reward receipt compared with typically developing youths. While previous research suggests that youths with DBD+CU exhibit decreased amygdala activation in response to threat-based stimuli during emotion processing, the amygdala is also broadly implicated in appetitive incentive motivation and salience detection (
5,
6). Our results are consistent with the finding that greater reward value in trait impulsivity, a core feature of youths in the DBD+CU group, is associated with increased amygdala activation following monetary reward receipt (
38). Adults with psychopathic traits have also been shown to exhibit heightened amygdala activation during automatic attention allocation in response to unexpected events (
39) (i.e., unexpected reward receipt in our task). Together, these findings suggest that youths with DBD+CU may show greater reliance on the motivational saliency of external feedback, including the value of obtained rewards, because they are less adept at using internal signals to guide behavior.
Finally, we integrated region-of-interest findings to assess broader network-level activation by examining a higher-order latent reward network factor. For both reward anticipation and outcome, regions of interest showed good fit to the hypothesized reward network factor models. Invariance testing indicated that the reward network factor model fit equivalently across study groups for both reward anticipation and receipt. Our results are among the first to suggest that a higher-order reward network can be modeled. Nevertheless, we found preliminary evidence for divergence in mean-level network activation among youths with DBDs contingent on callous-unemotional traits and the reward phase. First, the DBD-only group not only showed reduced anticipatory reward-related activity within individual regions as described above but also across a broad reward network factor model. These findings suggest that youths in the DBD-only group may have specific difficulties integrating previous information about previously rewarded outcomes and subsequent reward-predicting cues (
11). In contrast, youths in the DBD+CU group showed increased left hemispheric network-level activation following reward receipt. This finding is consistent with those from studies that found youths with DBD+CU to display heightened reward responsiveness and a reward-dominant interpersonal style (
37,
40). Additionally, increased reward drive is associated with impaired processing of potential costs and difficulties initiating behavioral change once attention has been allocated toward motivationally salient stimuli, which could explain the highly exploitative and aggressive behavior of youths with DBD+CU.
There are several limitations to our study that should be considered. First, the cross-sectional design did not allow for causal inferences. Follow-up studies that utilize future waves of ABCD Study data are paramount to test whether reward-processing deficits represent prospective risk markers for persistent DBD or callous-unemotional traits. Second, while we observed increased amygdala activation during reward receipt among youths in the DBD+CU group, further studies are needed to examine whether this is specific to monetary reward (i.e., versus social reward). Third, while our regions of interest fit a higher-order reward network factor model, the model represents a simplified version of the reward network, which likely encompasses an expanded set of brain regions. Fourth, information about psychotropic medication use or clinical services uptake was not assessed. We also note that youths assigned to DBD groups were not diagnosed by a clinician, and 12% of youths included in the overall DBD group did not meet K-SADS-PL criteria for DBDs. Finally, although we found no evidence that sex moderated our findings, the age of our sample (9–10 years old) could predate many of the sex-based differences in brain-behavior associations that are thought to emerge during adolescence following pubertal development.
In summary, we found evidence that DBDs are characterized by decreased neural activation across reward and cognitive-control regions during reward anticipation, which was particularly evident in the DBD-only group. Moreover, youths with DBDs showed relatively increased activation of salience and reward regions during reward receipt, which was particularly evident in youths with DBD+CU. These neural profiles provide further insight into the behavioral features underpinning DBDs, including potentially divergent profiles contingent on callous-unemotional traits. In particular, the combination of relatively reduced reward-anticipation recruitment of the dACC with an oversensitivity to the value of obtained rewards may explain why youths with DBDs persist in harmful behaviors that they perceive to be rewarding. Importantly, this may underlie many of the hallmark features of DBDs, including impaired decision making, frustration, reactive aggression, and impulsivity. Future studies conducted within a prospective longitudinal design can establish whether differences in neural sensitivity to reward represent a predictive biomarker of youths at risk for DBDs, including those with callous-unemotional traits, which could inform personalized interventions.