O bsessive-compulsive disorder (OCD) is a relatively common, and often disabling, psychiatric disorder.
2,
3 It is characterized by obsessions (unwanted, recurrent intrusive thoughts that cause anxiety) and compulsions (repetitive behaviors that the patient feels driven to perform, often in response to an obsession) which generally coexist.
4 Imaging studies have consistently shown abnormalities in specific brain areas in patients with OCD. However, how the normal functioning of these brain areas is altered to produce the symptoms of OCD remains unknown. In this article, we assert that the completion of complex behaviors is normally accompanied by a reward signal, and that abnormalities in this process could account for some of the symptoms of OCD. We present evidence for this view and propose testable hypotheses.
This article is separated into five parts. The first part reviews the literature on the brain areas associated with OCD. A review of imaging studies on OCD was conducted by performing a MEDLINE search through 2006 on the term “obsessive-compulsive disorder” and one of the following terms: “imaging,” “CT,” “computed tomography,” “MRI,” “magnetic resonance imaging,” “PET,” “positron emission tomography.” The resulting abstracts were screened by one of the authors (EDH), and relevant studies were reviewed. In addition, we evaluated selected reviews.
5 –
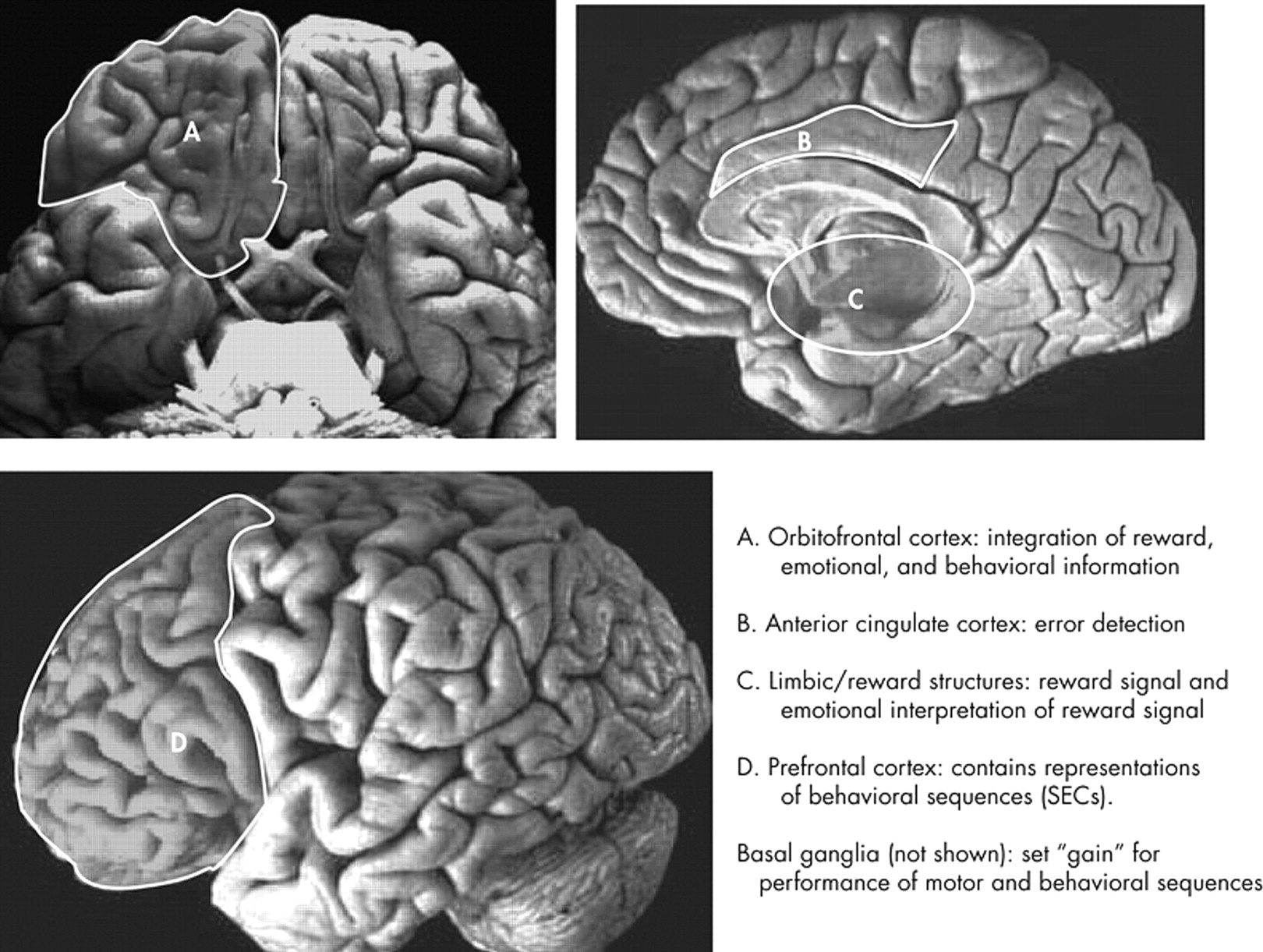
8 The second part of this article reviews recent findings from the field of cognitive neuroscience on the functions of these brain regions. We focus on the role of the orbitofrontal cortex (OFC) and reward structures in reinforcement, the basal ganglia in setting the threshold for activation of motor activity and complex behaviors, and the anterior cingulate cortex (ACC) for error detection (see
Table 1 ). The third part discusses the role of the prefrontal cortex (PFC) in the execution and reinforcement of complex behaviors. The fourth part presents some previous models of OCD. Finally, the fifth part proposes a new model that integrates the findings presented in the first three parts of the paper.
The Brain Areas Involved in OCD
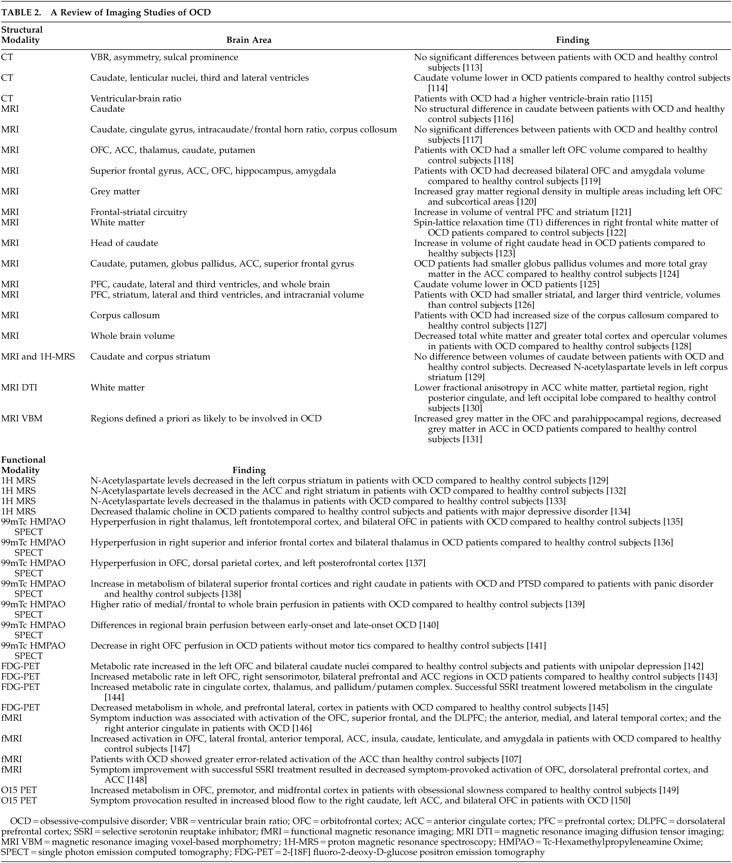
The majority of both structural and functional imaging studies have shown differences in the PFC, basal ganglia, ACC, and/or thalamus between patients with OCD and healthy comparison subjects (see
Table 2 ).
5 A recent meta-analysis reviewed functional imaging studies in OCD and found that the OFC (orbital gyrus) and head of the caudate were the only brain areas that significantly and consistently demonstrated increased tracer uptake in OCD patients relative to comparison subjects.
8 We will discuss the OFC, basal ganglia, ACC, and thalamus in this review, but will focus on the OFC and basal ganglia because these brain areas are most consistently associated with OCD in imaging studies.
8The imaging findings reviewed in Table 2 are corroborated by the finding that disrupting connections between the OFC, ACC, thalamus, and basal ganglia by means of a cingulotomy, anterior capsulotomy, or subcaudate tractotomy results in a symptomatic improvement in most OCD patients.
6,
9 –
14Some studies have examined the development of symptoms of OCD after brain injury.
15 Damage to the basal ganglia (especially the caudate), the OFC, and the ACC
16 –
22 are associated with the acquisition of OCD symptoms following brain injury.
15 Dysfunction of the basal ganglia secondary to a streptococcal infection
23 or encephalitis lethargica
24 has also been associated with the development of OCD symptoms. One report showed an association between lesions in the mesial frontal region (including the ACC) and collecting behavior resembling OCD.
25 Another demonstrated that repetitive motor activity in patients with dementia is uniquely associated with right ACC hypometabolism.
26 We have observed that repetitive motor activity is associated with right caudate and OFC atrophy in patients with frontotemporal dementia (Huey, presentation, UCSF 5th International Conference on Frontotemporal Dementia, 2006).
The Prefrontal Cortex (PFC) in the Execution and Reinforcement of Complex Behaviors
Inherent in our discussion so far is the assumption of a conservation of mechanisms of reward between nonhuman primates and humans and between events rewarding to nonhuman primates and humans (e.g., receiving food) and events that are specifically rewarding to humans (e.g., receiving money). However, comparing human and nonhuman reward raises the following question: what are the boundaries of the rewarding event for complex behaviors? Rewarding events for complex behaviors are often associated with several superordinate and subordinate rewarding events. For example, the rewarding experience of enjoying a dinner in a restaurant with a friend lasts a few hours, but it is a component of the larger rewarding friendship (which could last for a lifetime) and is composed of shorter rewarded events (e.g., enjoying a story your friend tells during the dinner). How are the boundaries set?
Our laboratory has proposed that the PFC stores memories of behavioral sequences termed “structured event complexes” (SECs) that have beginnings and ends, but exist in nested hierarchies. For example, eating in a restaurant would be such an SEC, and it would exist as several different variants (e.g., eating at a fast-food restaurant, eating at a fancy French restaurant, etc.) which would come under the superordinate category of eating in a restaurant. We have proposed that these SECs are abstractly encoded in the PFC similar to the way in which memories of complex motor programs are encoded in more posterior cortexes. We hypothesize that the perceived boundaries of these SECs signal transitions for the purposes of reward, and that completing SECs can be inherently rewarding. In support of this hypothesis, prefrontal cortical neurons in macaques exhibit phasic peaks of spike activity at the beginning and endpoint of sequential tasks.
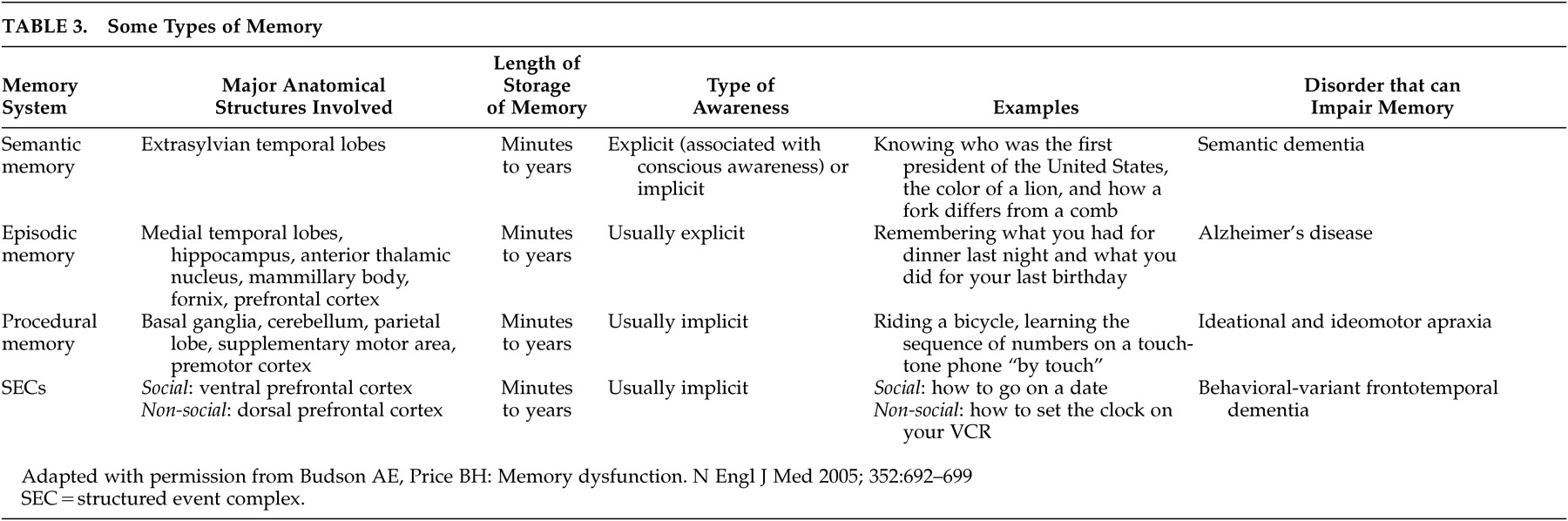
72In this theory, representations in the PFC differ from other types of memories that people are more familiar with, for example, semantic memory processes (
Table 3 ). Semantic (knowing the capital of France) memory is usually explicit (associated with conscious awareness), but it can be implicitly primed. SECs, in contrast, are usually implicitly recalled and executed often over long periods of time in the absence of directly relevant stimuli. This mechanism is most similar to that of procedural memory in the premotor cortex and supplementary motor area; one is not consciously aware of the ability to swim or ride a bicycle, yet one can execute these motor memories with minimal conscious control. We hypothesize that behavioral programs in the human PFC evolved from simpler motor programs in more posterior cortex.
73The types of memory outlined in
Table 3 work together in an integrated manner. For example, imagine you meet someone at a party and he or she gives you his or her telephone number. You will likely encode an episodic memory that this event occurred. You will keep the telephone number active in working memory until you can either write it down or encode the number in long-term memory through active rehearsal. If you dial the number enough, you may forget the actual digits and instead rely on the procedural memory of dialing the number on a touch-tone phone. Assuming that getting the other person’s number is a successful social outcome, you will encode a memory in the PFC of the behavioral sequence that led to this outcome, to be able to best repeat it in a similar situation.
73 –
75This theory asserts that related SECs are neuroanatomically localized together in specific areas of the PFC, an assertion that has obtained empiric support. The frequency with which healthy subjects had experienced an event determined how anterior or posterior fMRI activation was observed when the subjects determined if the events were correctly ordered,
76 neurons in the lateral PFC of monkeys selectively exhibit activity for specific categories of behaviors
77 and when the monkeys remember and perform particular action sequences.
78 Patients with PFC lesions (and thus disruption of their SECs) should show deficits in ordering events into a coherent sequence. Patients with PFC damage have particular difficulty sequencing events,
79 can generate a normal number of actions, but have difficulty ordering those actions into a coherent script,
80,
81 and appear to lose infrequently used SECs before frequently used (and thus overlearned) SECs.
80,
82 Patients with dementias affecting the frontal lobes typically demonstrate deficits in social behaviors with relative preservation of episodic memory, while patients with dementia initially affecting the medial temporal lobes (e.g., Alzheimer’s disease) typically demonstrate initial deficits in episodic memory with relative preservation of social behavior.
83The human brain can flexibly respond to events with an almost infinite variety of behaviors. How can such a large number of potential behaviors be encoded as memories? We hypothesize that humans (and other animals) can flexibly coactivate and combine SECs to form a large number of behaviors. This process could be analogous to language; a finite number of words and linguistic rules allow humans to form an almost infinite variety of expressions. In support of this, healthy adults are able to flexibly order the components of a plan while young children and patients with PFC damage tend to rigidly execute plans.
84 Also, rather than performing an infinite variety of behaviors, healthy comparison subjects generally perform a relatively small number of high frequency behaviors in their daily lives.
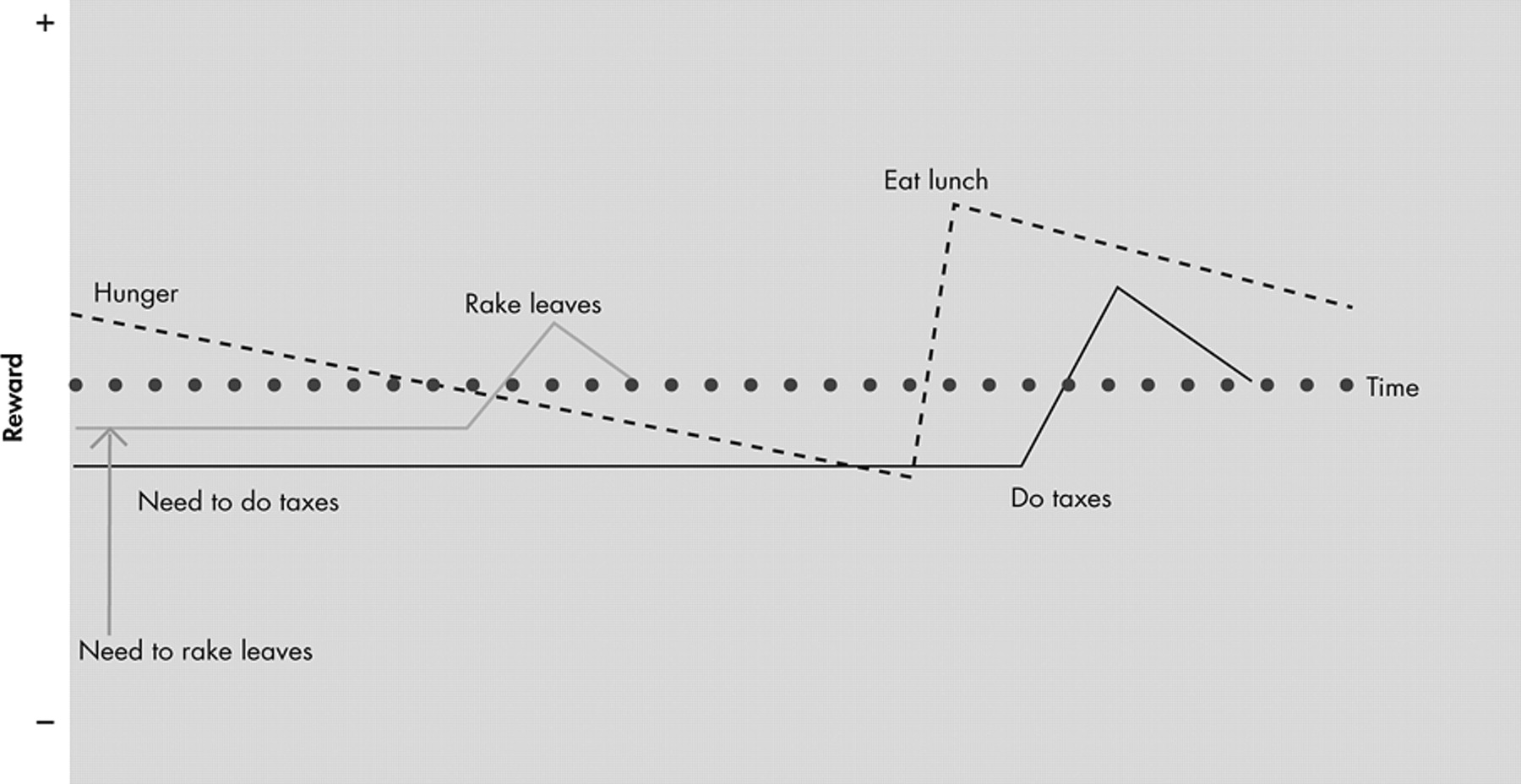
85So far, we have explained the reinforcing properties of performing SECs. However, the interaction between behavior and reward is bidirectional and dynamic. If performing a certain SEC is rewarding, being prevented from performing that sequence would be punishing. Completion of a punishing SEC would result in reinforcement when the punishment is removed after completion of the behavior. An example of this is doing one’s taxes. Few people enjoy doing their taxes, but they do enjoy the feeling of relief when they have completed this onerous, but necessary, task.
Expectation of outcome can affect the reward value of an event. Schultz
86 has demonstrated the importance of “prediction error” in reward and learning. Prediction error refers to the difference (positive or negative) between the expected and received reward. Certain dopamine neurons in the pars compacta of the substantia nigra and the medially adjoining ventral tegmental area (groups A8, A9, and A10) and the OFC of macaques respond most to a stimulus that is paired with an unpredicted reward.
86,
87 Thus, the same stimulus could be rewarding or punishing depending on expectation. For example, you could receive punishment by learning that one-half of your lottery winnings will go to taxes after learning that you have won the lottery, even though it is a large net financial gain. The dynamic nature of reward over time and the role of expectation make the concept of a “baseline” of reward state for an animal difficult to define. We believe that the reward state of an animal at any given time is, in part, a summation of the reward values associated with the many different SECs active and at different stages of completion at that moment (see
Figure 3 ).
Previous Models of OCD
So far, we have presented research from Rolls
47 showing that the OFC is central for reward mechanisms. The ACC plays a central role in error detection for complex behaviors. Frank, O’Reilly, and colleagues
52 –
54 have proposed a model with the basal ganglia setting the “gain” for activation of representations in the PFC similar to the way in which the basal ganglia sets the “gain” for activation of motor programs in the supplemental motor area and premotor cortex. Our laboratory has asserted that SECs exist in the PFC similarly to motor programs contained in the supplementary motor area and premotor cortex, and that performance of those SECs is rewarded. Schultz and colleagues
86,
87 have demonstrated that areas involved in reward, including the OFC, are activated by a difference between the expected and observed outcomes of events. In this part, we discuss some current models of OCD (see
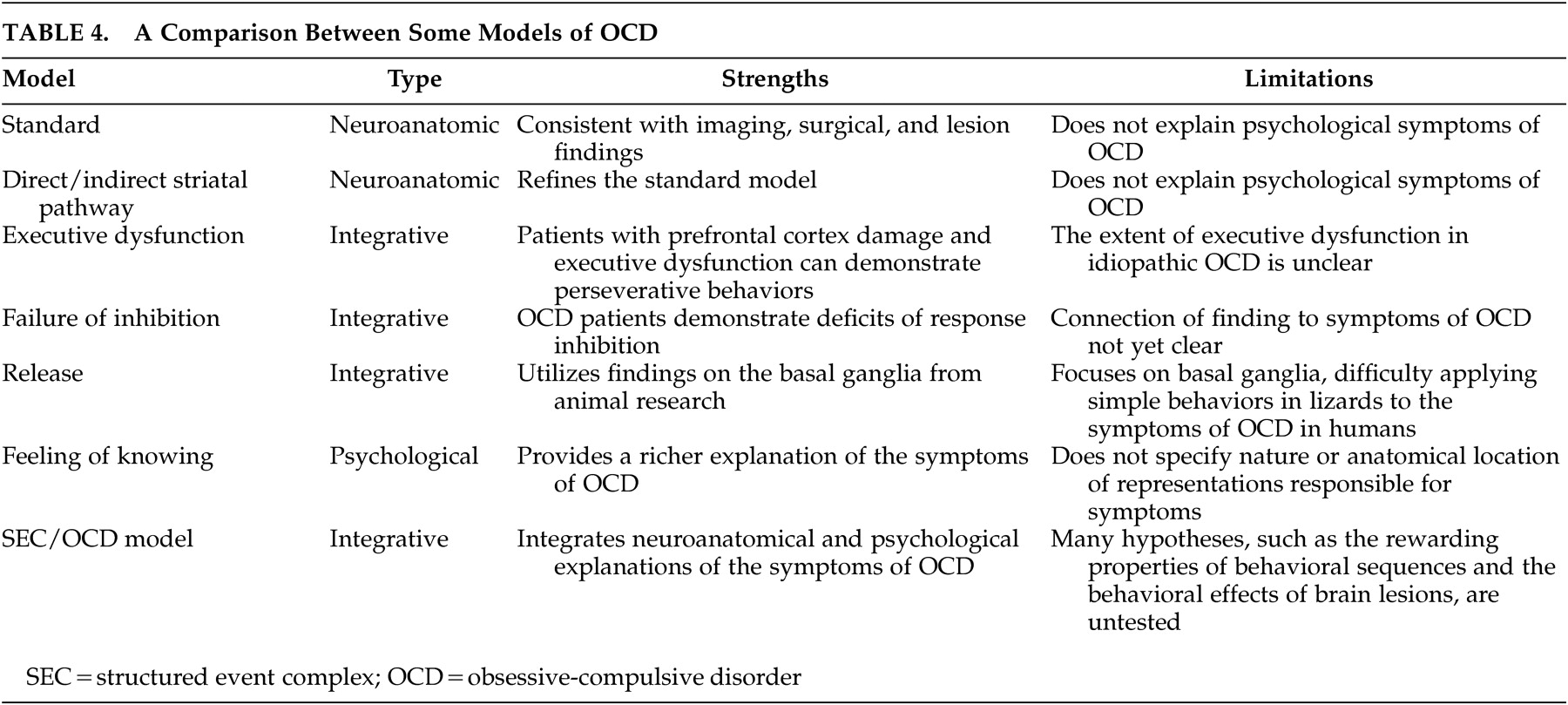
Table 4 for a comparison). In the next section we propose a new model that suggests that the symptoms of OCD arise from abnormalities in the reward mechanisms of complex behaviors.
The Standard Model
The most accepted neuroanatomic model of OCD is based on the finding that there are separate cortico-basal ganglia-thalamic-cortical loops,
70,
71 (although recent evidence suggests that these loops are not as separate as previously thought).
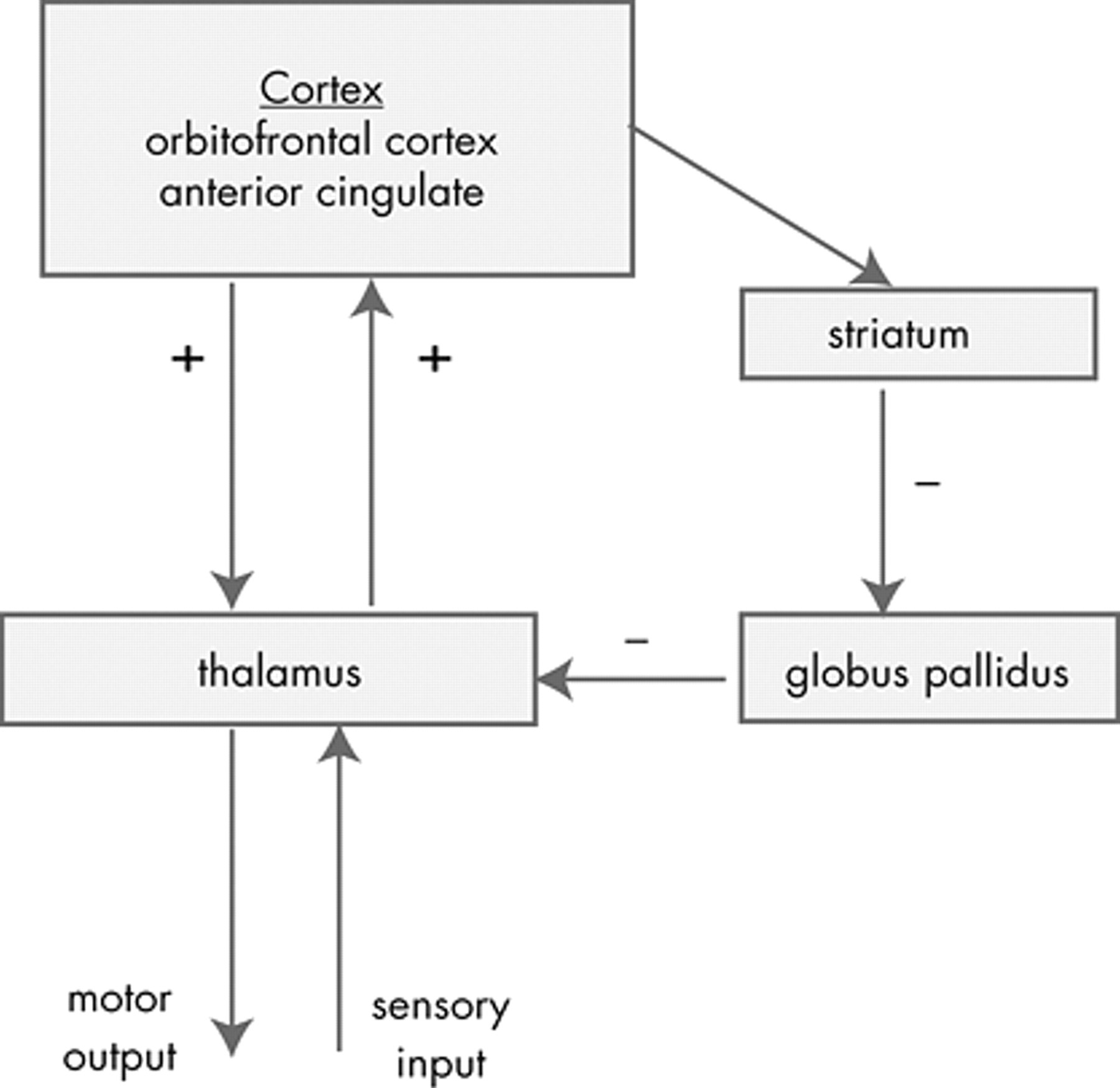
88 The standard anatomic model of OCD proposes that the symptoms of OCD are caused by dysfunction of elements of a PFC-basal ganglia-thalamic-PFC loop
89 –
93 (
Figure 4 ). The imaging findings presented above support that these structures are involved in OCD. In addition, surgical interruption
6,
9 –
14 or deep brain stimulation
94 of the anterior internal capsule can reduce the symptoms of OCD. Overactivation of this loop is suggested by the hypermetabolism of these structures observed in the imaging studies presented in the first part of this article.
The advantage of this model is that it is consistent with the evidence collected to date on OCD. This model forms the neuroanatomic basis of most subsequent models. The limitation of the standard model is that while it specifies the brain structures involved, it does not provide a psychological explanation for the specific symptoms of OCD.
Direct/Indirect Striatal Pathways
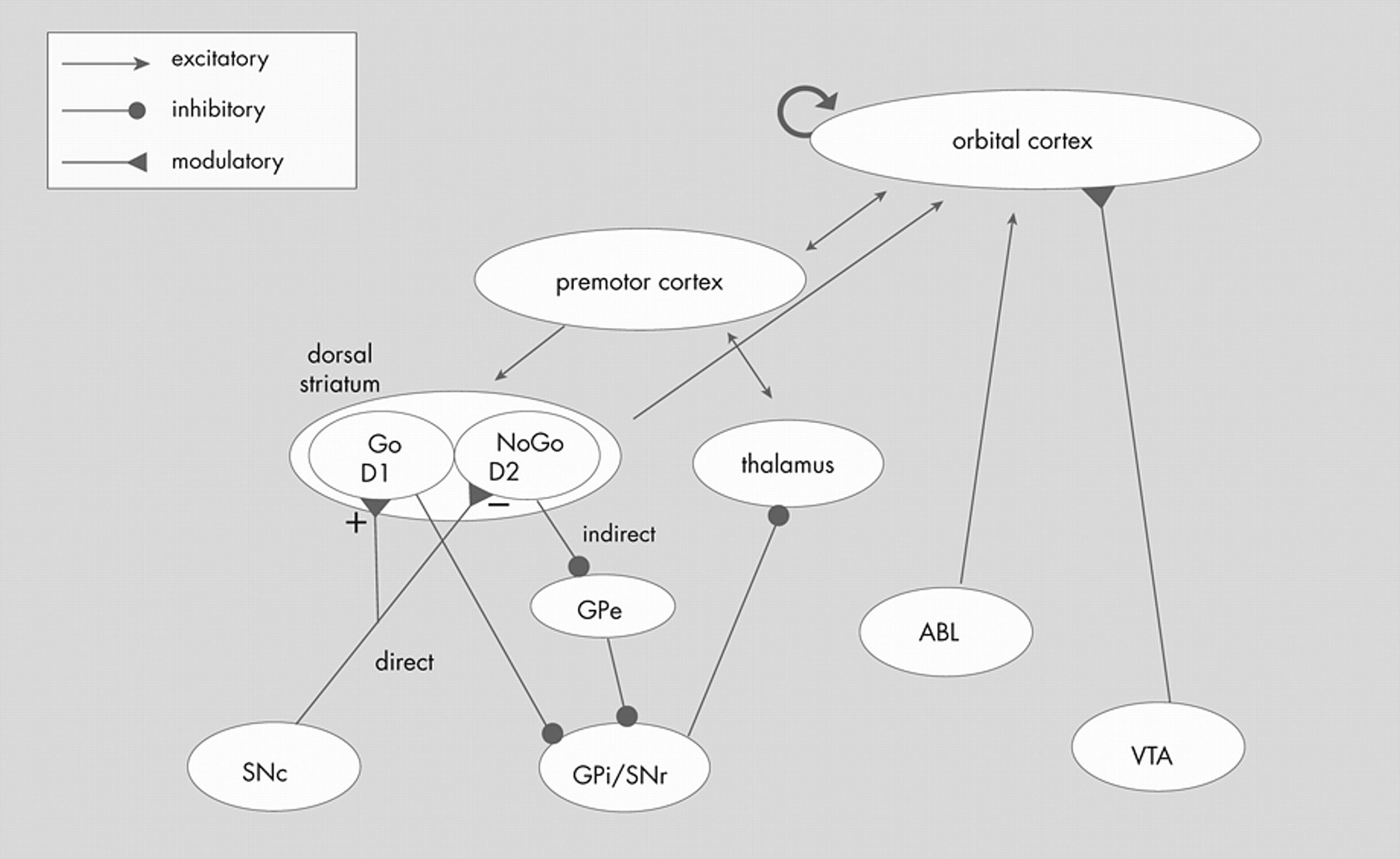
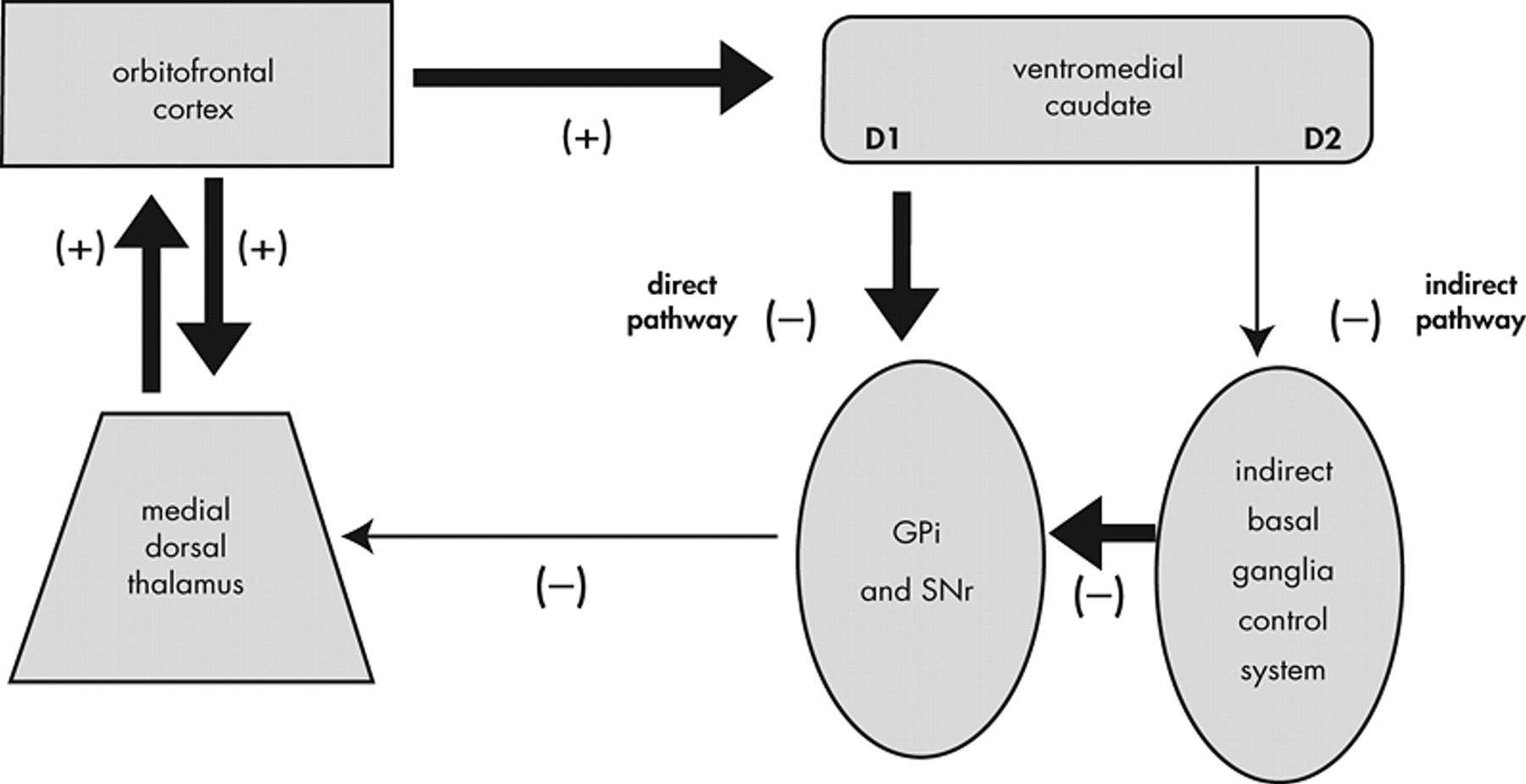
The standard anatomic model has been refined by specifying that overactivation of the direct pathway in the basal ganglia relative to the indirect pathway results in an orbitofrontal-subcortical hyperactivity (
Figure 5 ). According to this model, “[p]atients with OCD, however, may have a low threshold for system ‘capture’ by socioterritorial stimuli, possibly caused by excess ‘tone’ in the direct relative to the indirect orbitofrontal-subcortical pathway, allowing for concerns about danger, violence, hygiene, order, and sex to rivet attention to themselves.”
95This model adds explanatory power to the standard model by proposing a specific mechanism within the striatum that results in overactivation of the neuroanatomical loop of the standard model. In support of this model, patients with excessive nigrostriatal dopaminergic input (such as patients with Huntington’s disease) have excessive motor output.
95 This model is supported by the imaging findings presented in the first part of this article and because damage to specific basal ganglia structures (e.g., caudate) is associated with the development of symptoms of OCD. The limitation of this model, similar to the standard neuroanatomical model, is that it does not specify or explain the psychological mechanisms of OCD. For example, how do concerns “rivet attention to themselves?” It also focuses on dysfunction in the basal ganglia, but does not specify the role of the OFC or explain how patients with OFC lesions can develop aspects of the OCD syndrome.
Models Based on Other Brain Areas
Other theorists have focused more on the orbitofrontal cortex (OFC) in modeling OCD. Chamberlain et al.
7 discuss the failure of inhibition in patients with OFC lesions and propose that a similar failure to inhibit contributes to symptoms of OCD. Some have implicated the anterior cingulate cortex (ACC) by proposing that faulty error detection may be central to the pathogenesis of OCD.
91,
96 Others have suggested that dysfunction of reward mechanisms may contribute to the symptoms of OCD.
6“Release” Models
Baxter
92 demonstrated the role of the basal ganglia in “releasing” territorial display programs in lizards and proposed that the basal ganglia in patients with OCD may inappropriately “release” territorial behaviors. Stein and Lochner
97 have conceptualized OCD as a “dysfunction in the control of procedural strategies with inappropriate release of symptoms ranging from simple motoric stereotypies to more complex behavioral programs.” In the same paper, they note that many of the structures involved in OCD are also involved in learning and reward. They also observe that dopaminergic agonists can increase the symptoms of OCD and putative OCD spectrum disorders, including Tourette’s syndrome. Where in the brain and how these “behavioral programs” are represented, or why they are inappropriately released in OCD, is not specified. Graybeil and Rauch
56 hypothesized that anxiety suffered by OCD patients may indicate a “lack of loop closure between expected outcomes and the chunks of behavior that should generate them.”
“Feeling of Knowing” Models
Szechtman and Woody
98 have proposed a model of OCD based on the hypothesis that the symptoms of OCD arise from an inability to generate a normal “feeling of knowing” that would otherwise signal task completion. This deficit results in an overactivation of neural systems designed to respond to danger in the environment (which they term the “security motivation system”). They base their work on earlier cognitive theories of OCD including those of Janet,
1 Pitman,
99 and Reed.
100 Their theory is supported by interviews that revealed that the majority of OCD patients describe their symptoms as being “unable to stop” the behavior rather than being forced to continue.
100,
101 Also, patients with OCD often engage in few but extended episodes of compulsive behavior during the day rather than excessively frequent episodes but normal duration, which is consistent with an inability to stop the compulsive behavior.
102The “Structured Event Complex” Model of OCD
The Structured Event Complex (SEC)/OCD model builds upon those proposed by Stein and Lochner
97 and Szechtman and Woody.
98 However, in the SEC/OCD model we specify how abnormal interactions of representations of complex behaviors in the PFC, reward information in the OFC, error detection in the ACC, and reward and limbic structures can result in the symptoms of OCD. To our knowledge, this is the first model of OCD to fully integrate the neuroanatomy and psychological experience of OCD.
We propose that the initiation of an SEC is accompanied by a motivational signal, likely determined through interaction between reward structures (including the OFC) and limbic structures, experienced as motivational anxiety. This anxiety likely exists to motivate animals to complete necessary SECs. Completion of the SEC is accompanied by a reward signal, experienced as relief from anxiety. People with OCD have a deficiency in this process. They may receive only a fraction of the full relief from anxiety that most healthy people receive upon completing the SEC. Even after completion, the patient is left with the unpleasant sensation that the SEC is not done. In this way, our model resembles the “feeling of knowing” model proposed by Szechtman and Woody.
98 A difference is that we specify the nature and location of the task that is perceived as not completed (SECs contained in the PFC), and the relative contribution of other brain systems involved (see
Figure 6 ).
In this review, we have focused on the separable roles of the brain structures found to be involved in OCD. However, all of these brain areas communicate extensively, and are frequently coactivated in imaging studies of OCD. Our laboratory has previously proposed that the subjective experience of a particular mental state is biologically represented by synchronous activity of a system of brain areas, each of which contributes a component to the experience.
51,
103 In our model of OCD, the experience of OCD symptoms comes from binding together SECs in the prefrontal cortex (PFC), the reward signal in reward structures and the OFC, the threshold for activation of SECs in the basal ganglia, emotional relevance by limbic structures, and the error signal by the ACC. For example, an active contamination obsession in a patient with OCD could involve representations of the following:
1. an error signal of an incomplete task generated in the ACC;
2. punishment represented in the OFC and reward structures;
3. the emotional experience of anxiety arising from limbic structures;
4. a lowering of the threshold for a compensatory SEC in the basal ganglia;
5. activation of the compensatory SEC contained in the PFC.
The specific degree and nature of these coactivations could correspond to the particular cognitive and emotional state of the patient with OCD. In this way, we argue against the commonly held view that the PFC opposes emotional input from the limbic system.
104In the SEC/OCD model, the OCD patient’s cognitive interpretation of the feeling of leaving an SEC incomplete forms the basis of an obsession. The feeling is unconscious, but the patient explicitly attempts to assign it a cause and reduce it through conscious action. The explicit interpretation of the feeling is the “obsession” and the conscious attempt to reduce it is the “compulsion.” This interpretation is influenced by individual and societal attributes, but is often based on common themes (e.g., contamination or pathologic doubt). The societal influence on this interpretation can be observed in the change that has occurred in obsessional themes over time (e.g., no one had germ contamination obsessions prior to the proposal of the germ theory of infection). Patients with OCD may receive a fraction of the full reward signal with each performance of the SEC (or its elements). Thus each performance of the SEC (or its elements) can be partially successful at alleviating the anxiety reinforcing a maladaptive learning mechanism. For example, most of us, if our hands are dirty, feel a motivational anxiety to perform the SEC of washing our hands. Upon completion of this SEC we receive relief from the motivational anxiety. Patients with OCD do not receive full relief from the anxiety upon completion of the SEC, but may receive partial relief. They may, understandably, cognitively appraise the implicit continued motivational anxiety to mean that their hands are still dirty, despite having washed them. Thus there is dissociation in patients with OCD between the conscious awareness that one’s hands are clean, and the “feeling” that they are not. Consequently, the patient may repeat the hand-washing SEC, receiving partial relief with each repetition, until the motivational anxiety is resolved.
Findings presented above suggest that the basal ganglia can set the threshold for activation of motor programs in the premotor cortex and supplemental motor area by facilitating some motor programs and inhibiting others through reinforcement learning.
53,
54,
105 In the SEC/OCD model, the basal ganglia perform a similar role for the activation of SECs in the PFC. We propose that the basal ganglia set the threshold for activation of SECs, and if this threshold is lowered, SECs can be overactivated, resulting in excessive motor activity (e.g., tics) and/or the excessive activation of SECs. Thus damage to the basal ganglia that results in a reduction of its net inhibitory output to the OFC (such as occurs with caudate damage) can result in symptoms of OCD, as observed in cases of autoimmune basal ganglia damage.
106 The role of the basal ganglia in our model has some of the properties proposed in the “release” models of OCD.
92,
97Evidence for the SEC/OCD Model
The Structured Event Complex (SEC)/OCD model fits well with the research presented above about the roles of the OFC, the basal ganglia, and the ACC. The OFC is central for receiving and interpreting reward signals in a social and behavioral context, and can be especially activated when there is a large difference between the expected and received reward signal, as would be the case for patients with OCD in the SEC/OCD model. The basal ganglia set the threshold for activation of SECs. If this threshold were lowered, these SECs could be overactivated, resulting in excessive motor activity (e.g., tics) and/or the excessive activation of SECs. The ACC is involved in error detection (especially a discrepancy between expected and observed outcome), and thus in the SEC/OCD model we would expect it to be overactive in OCD where the OFC is receiving a neural message that the SEC was not successfully completed (as is observed in imaging studies). Patients with OCD demonstrate greater ACC activation than healthy subjects when the patients make errors that do not elicit OCD symptoms.
107Most of the evidence in favor of the “feeling of knowing” model supports the SEC/OCD model as well: patients with OCD usually report that they experience their symptoms as a feeling of being unable to stop an action with a lack of a sense of completion of an action,
100,
101 and engage in few but extended episodes of compulsive behavior during the day rather than episodes of excessive frequency but normal duration, suggestive of an inability to stop the compulsive behavior.
102The SEC/OCD model asserts that obsessions are primary in OCD and that compulsions are a secondary response to the obsession. In support of this, obsessions and compulsions most frequently co-occur in idiopathic OCD in adults,
4 obsessions and compulsions are usually thematically related,
108 and the majority of OCD patients report the obsession as the primary motivation for the compulsion.
100,
101 Specific compulsions are associated with particular brain areas,
109 consistent with the theory from our laboratory that specific SECs are regionally separable.
73 There is some evidence that the performance of SECs is inherently rewarding. Direct stimulation of reward pathways in rats resulted in repetitive stereotyped complex behaviors.
99,
110 Humans performing SECs
111 or responding to novel stimuli
112 show activation of reward structures on fMRI.
Experimental Predictions
The SEC/OCD model provides several testable hypotheses. Most previous anatomical models have proposed a primary source of the psychopathology of OCD (usually either the basal ganglia or OFC). A limitation of that approach is that it fails to explain how damage to several different structures can lead to acquired symptoms of OCD in lesion studies. In the SEC/OCD model, we hypothesize that damage to different brain structures, or damage to the communication between structures, will result in different and separable aspects of the OCD syndrome (
Table 5 ). Because the PFC contains memories of SECs, patients with acquired OCD from PFC damage should show impaired performance of SECs. This is in contrast to patients with idiopathic OCD and patients with OCD acquired from basal ganglia damage who should have relatively preserved performance of SECs. We also hypothesize that, because of the role of the OFC in perceiving and interpreting reward and anxiety signals, patients with acquired symptoms of OCD from OFC damage will have fewer obsessions and less anxiety compared to patients with idiopathic OCD who have comparable levels of compulsive behavior. This has been reported,
17 and we have clinically observed this in our laboratory in patients with frontotemporal dementia (Huey, presentation, UCSF 5th International Conference on Frontotemporal Dementia, 2006). However the literature on this topic is limited because studies have been performed retrospectively on patients identified and defined by having the entire OCD syndrome. We also hypothesize that patients with symptoms of OCD acquired from brain injury will show hypometabolism of injured structures and areas of the brain that are closely connected, in contrast to the hyperactivation of these brain areas observed in idiopathic OCD. Prospective studies on patients with brain damage should be performed to determine the relative contributions of different brain areas to the OCD syndrome (
Table 5 ). Newer imaging techniques such as diffusion tensor imaging may be useful for exploring abnormalities in white matter tracts between brain areas involved in idiopathic OCD.
The potential rewarding properties of the completion of structured event complexes (SECs) in healthy subjects have been minimally examined. Functional MRI studies of symptom provocation with careful clinical correlation in patients with idiopathic OCD could be performed with the hypothesis that the ACC will be initially activated (and correspond to the initial detection of the provoking stimulus), followed by OFC and limbic activation (corresponding to the anxiety provocation), then the basal ganglia and PFC (corresponding to activation of the compensatory compulsion). The SEC model of OCD is amenable to computer modeling, similar to that performed by Frank, O’Reilly, and colleagues
52,
54 to model basal ganglia-OFC interactions. Finally, the SEC/OCD model suggests that lesioning certain brain areas in animals can result in behaviors similar to aspects of the idiopathic OCD syndrome (
Table 5 ).
CONCLUSION
Imaging, surgical, and lesion studies suggest that the OFC, basal ganglia, and ACC are involved in the pathogenesis of OCD. Recent research on the normal function of these brain areas demonstrates the role of the OFC in reward, the basal ganglia in affecting the threshold for activation of motor and behavioral programs, and the ACC in error detection. We discussed theories that the PFC stores memories of behavioral sequences (called SECs) and that initiation of an SEC results in motivational anxiety that is relieved upon completion. We discussed previous models of OCD and proposed a new model of OCD (the SEC/OCD model), which hypothesizes that a deficit in the relief of anxiety that usually accompanies the completion of an SEC is responsible for the symptoms of OCD. Specifically, this anxiety forms the basis of an obsession, and a compulsion is an attempt to receive relief from the anxiety by repeating parts of, or an entire, SEC. We discussed empiric support for the SEC/OCD model and specific experimental predictions of the model. We believe that the SEC/OCD model explains the specific symptoms of OCD and integrates the neuroanatomy and psychology of this disorder better than previous models.
Addendum
Studies identified in a non-systematic review of papers published between the completion and publication of this article are mostly supportive of the SEC/OCD theory. Frontotemporal dementia affecting the OFC is associated with stereotypic behaviors as predicted in Table 5.
151 Deficits in reversal learning linked to the OFC were demonstrated in patients with OCD
152,
153 and their unaffected relatives,
153 supporting the assertion of the SEC/OCD theory that the inability to stop an action is associated with risk for OCD. Similar results were found for task-switching in patients with OCD.
154 Damage to the medial striatum (including the head of the caudate) in monkeys also resulted in impairments in reversal learning, as would be predicted from Table 5.
155 One study showed specific sites of grey and white matter volume decreases associated with specific symptom clusters of OCD, which is supportive of the SEC/OCD theory, which proposes that the PFC contains separable representations.
156 However, some of the associations occurred in brain areas other than the PFC, which is not supportive of the theory (although the association seen in these areas, such as caudate, could be related to their action on the PFC).