Bipolar disorder is a psychiatric condition with a lifetime prevalence of approximately 1.2% (bipolar I and bipolar II subtypes combined) of the population of industrialized nations (
1, p. 161). The course of this disorder is frequently poor, with a risk of chronicity of more than 20%
(2) and a higher than average risk of suicide (
1, p. 228). The familial aggregation of bipolar disorder is well established
(3–
5); both adoption
(6,
7) and twin
(8) studies show that a substantial proportion of the variance in the etiology of bipolar disorder may be attributed to genetic factors.
In the last two decades, rapid advancement in molecular genetic techniques and more accurate phenotype assessment have increased the chances of finding a relationship between psychiatric disorders (i.e., phenotypes) and DNA polymorphisms. Consequently, investigators in a series of genetic studies have tried to establish linkage between bipolar disorder and markers on all chromosomes.
One candidate gene on the X chromosome that is of particular interest for bipolar disorder is the gene for monoamine oxidase A (MAOA), since this MAO isozyme plays an important role in the degradation of the neurotransmitters norepinephrine, epinephrine, dopamine, and serotonin. In fact, several studies have demonstrated abnormalities in serotonergic neurotransmission in mood disorders. The concentration of the major serotonin metabolite, 5-hydroxyindoleacetic acid, was found to be low in the CSF of a subgroup of unipolar depressed patients who had attempted suicide by violent means
(9). Moreover, lower than normal binding of [
3H]imipramine or [
3H]paroxetine to brain serotonin uptake sites has been observed in depressive patients
(10,
11) and patients with bipolar disorder
(12).
Several studies have focused on the MAOA gene (
MAOA) that has been mapped to Xp11.12–Xp11.4 on the short arm of the X chromosome
(13,
14). In 1993, Brunner and colleagues
(15) reported evidence of disturbed monoamine metabolism and an X-linked borderline mental retardation with a behavioral phenotype that resembled a manic syndrome. The authors established linkage between the phenotype and a nonsense mutation in
MAOA (16). Studies of the association between bipolar disorder and
MAOA have mainly focused on three markers in this gene: 1) a dinucleotide repeat polymorphism referred to as MAOA-CA
(17), 2) a dinucleotide repeat directly adjacent to the imperfectly duplicated 23-base-pair variable number of tandem repeats (VNTR) motif
(18), and 3) a restriction fragment length polymorphism (RFLP) resulting from single-base-pair substitution in the third base of a triplet codon
(19).
The findings of the available association studies involving these markers are contradictory. In a study sample in the United Kingdom, Lim and colleagues
(20) found an association between bipolar disorder and alleles of the three
MAOA markers among the overall sample and among female patients. Within the MAOA-CA repeat polymorphism, the major difference in the allelic distribution was found for the a2 allele (less frequent in bipolar subjects than in healthy subjects) and the a5 allele (more frequent in bipolar subjects). In another study conducted in the United Kingdom, Rubinsztein and colleagues
(21) replicated the different allelic distribution on the MAOA-CA but not the RFLP locus when male and female subjects were pooled. A significant association between bipolar disorder and MAOA-CA was also observed in a Japanese sample
(22). However, the alleles overrepresented in those bipolar subjects were different from those in the study by Lim and colleagues
(20).
These positive findings were not replicated by Craddock and colleagues
(23) in another British sample, N�n and colleagues
(24) in a German sample, and Parsian and Todd
(25) in a sample of non-Hispanic whites in the United States.
In order to resolve discrepancies between the British association studies of
MAOA, Rubinsztein and colleagues
(21) pooled the subjects of the three U.K. studies in a meta-analysis and found a significant difference between allele frequencies of the patient and comparison groups at the MAOA-CA locus and the RFLP locus for the overall sample and for the female, but not male, subjects.
As differences across studies may be attributable to either undetected population stratification
(24,
25) or real genetic differences between study populations
(26), replication studies with different populations are crucial. Consequently, we attempted to replicate the association between bipolar disorder and
MAOA in French and Swiss populations, using a larger sample than those included in previous studies.
Method
Patient and Comparison Groups
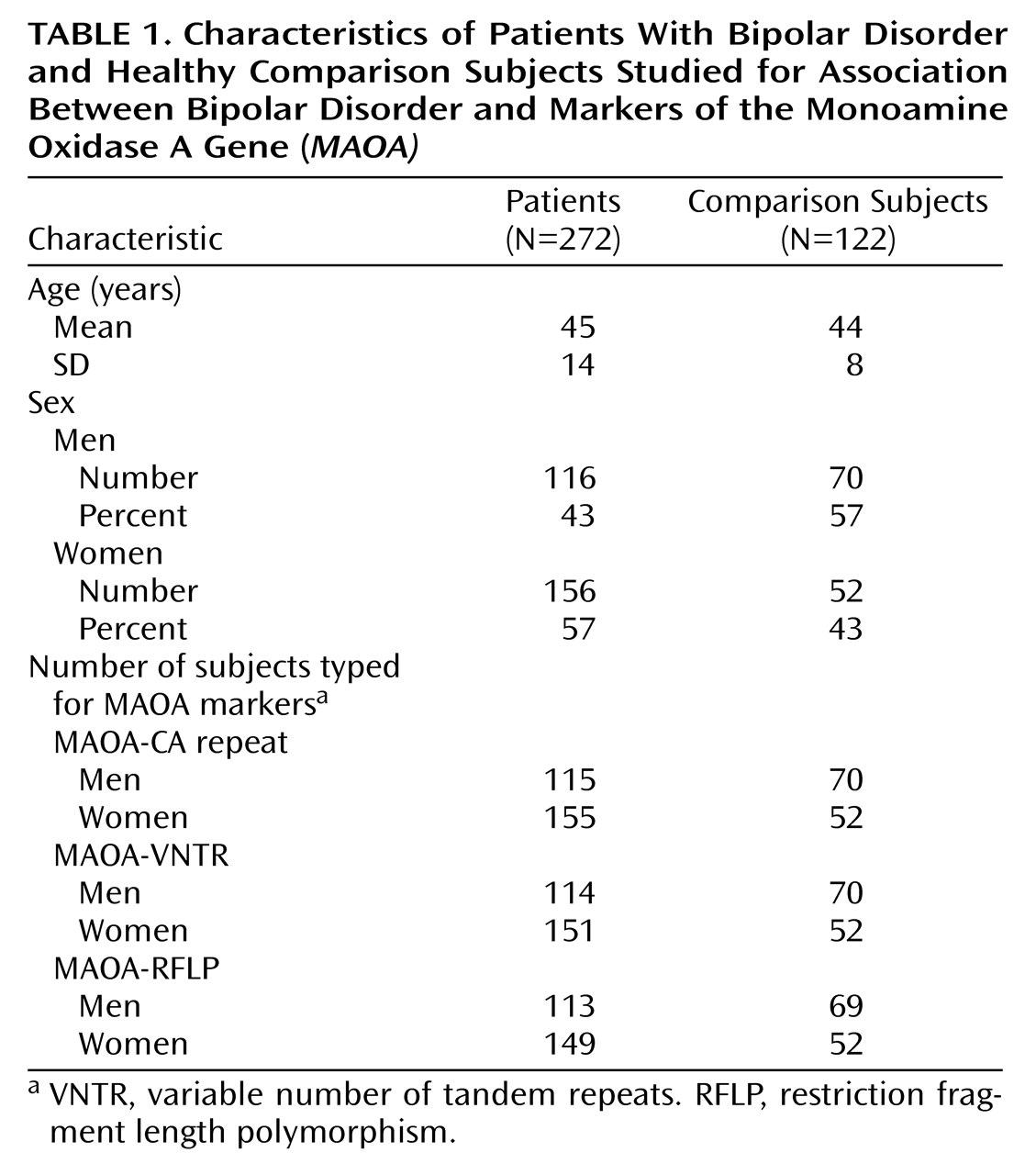
The study sample consisted of 272 bipolar inpatients and outpatients (bipolar I, N=207; bipolar II, N=65) and 122 healthy subjects. The four patient recruitment sites were the Piti笓alp賲i籥 and the Bic鳲e hospitals in Paris and the adult psychiatric departments of the Universities of Lausanne and Geneva in Switzerland. The maternal and paternal grandparents of all subjects were required to be Caucasians originating from western European countries. The subjects in the comparison group were persons without a history of major psychiatric disorders who were recruited from blood donors at the Piti笓alp賲i籥 hospital and from orthopedic patients at the two Swiss recruitment sites. In addition, the blood donors in Paris were required to be 1) older than 35 years and 2) without a family history of mood disorders or suicide. The patient and comparison groups did not differ by mean age but did differ by gender: women predominated in the patient group, while men predominated in the comparison group. A description of the study sample is provided in
Table 1.
All subjects were assessed by means of the semistructured Diagnostic Interview for Genetic Studies developed by the National Institute of Mental Health
(27) and translated into French
(28). The English version has demonstrated high interrater and test-retest reliability for the major DSM-IV axis I disorders
(27). Similar psychometric properties have also been found for the French version, tested in Lausanne
(29). Information on the family history of the comparison group in Paris was obtained by using the Family Interview for Genetic Studies
(30).
After a complete description of the study was provided, written informed consent was obtained from the subjects.
Laboratory Methods
Genomic DNA was extracted from white blood cells after centrifugation of 15 ml of edetic acid blood samples.
For polymerase chain reaction (PCR), 100 ng of genomic DNA was amplified by means of a thermocycler. PCR was performed in a 50-ml volume containing 20 pmol of different primers, 1.5 or 1.0 mM MgCl2, 200 nM deoxyribonucleotide triphosphate (dNTP), and 2 U of Taq polymerase.
For determination of MAOA-CA, 8 ml of PCR products was deposited on a 1.5-mm-thick 12% acrylamide gel. Electrophoresis was run for 7 hours at 250 V. The gels were stained with ethidium bromide and visualized under ultraviolet light. After having tested several alternatives, we chose this method (use of denaturing 5% acrylamide gel and labeled PCR products, hybridization of membrane with MAOA probe after gel transfer).
For MAOA-VNTR, 5 ml of PCR products was denatured by heating it at 95˚C for 3 minutes and running it on denaturing 5% acrylamide gel. Electrophoresis was done at 55 W for 3 hours; the labeled M13 sequence was the size marker of different PCR products. After fixation in a 10% ethanol and acetic acid bath, the gel was dried and exposed overnight.
For determination of MAOA-RFLP, 8 ml of PCR products was digested overnight with 5 U of endonuclease Fnu4HI in appropriate buffer. The digested samples were run on a 2% agarose gel, stained by ethidium bromide, and visualized under ultraviolet light.
Statistical Analysis
As some alleles of the MAOA-CA (12 alleles) are rare, all chi-square probability distributions of the alleles of this marker were estimated by using the program CLUMP, version 1.1 (10,000 simulations)
(31). This method is based on Monte Carlo simulation and estimates the significance level of the observed difference in allelic distribution between case and comparison groups when the traditional chi-square test is not applicable in large contingency tables with expected cell counts smaller than 5.
In accordance with the methods of Lim and colleagues (20) and Craddock and colleagues
(23), alleles at the MAOA-VNTR marker were first divided into five subgroups (V1 to V5). As we had no V1 or V2 and very few V5 alleles, the VNTR alleles were dichotomized into short (V4 and V5) and long (V3) alleles. Analyses of the distributions of the collapsed MAOA-VNTR alleles and the MAOA-RFLP alleles (only two) were performed by using chi-square statistics or Fisher’s exact tests for two-by-two tables when necessary.
Several comparisons were made to ensure the appropriateness of data pooling across bipolar subtypes and recruitment sites. As we did not find any significant differences in the allelic distribution of any of the three markers between bipolar I and bipolar II disorder, the two bipolar subtypes were combined in further analyses. Similarly, there was no evidence of genetic heterogeneity between the recruitment sites in Switzerland and France for either the patients or healthy subjects, therefore suggesting that combining data across sites was also appropriate. Finally, neither the patients nor healthy subjects demonstrated sex differences in allelic frequencies at the three marker loci. For this reason, the results of the comparison of allelic distributions in the patients and healthy subjects are presented for each gender and the overall sample. As the analysis comparing allelic distributions in the patients and healthy subjects included the independent testing of three markers by gender (six comparisons), the significance level was adjusted by using Bonferroni correction (significant p<0.05/6=0.0083).
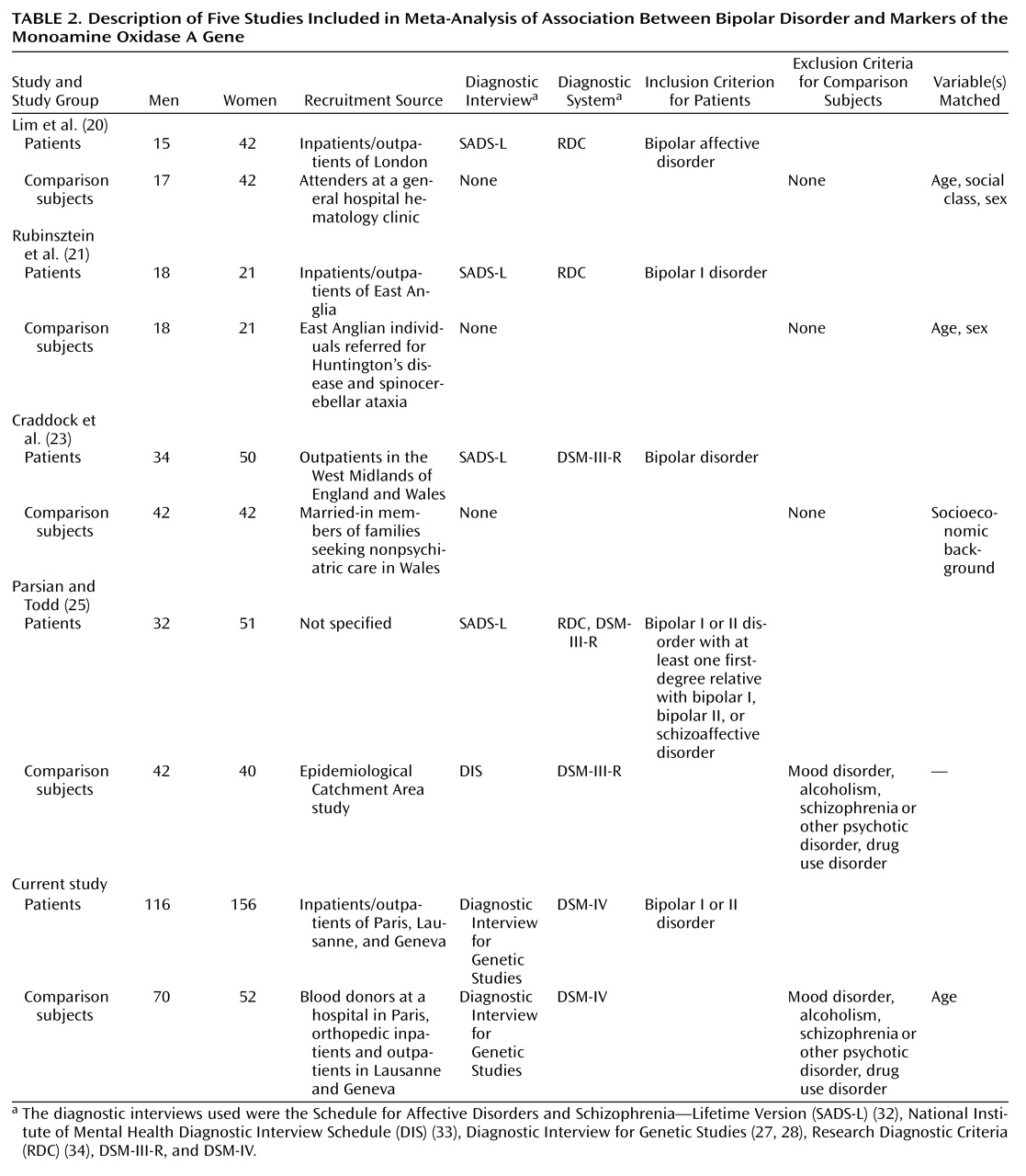
In order to resolve discrepancies across studies, we also tested whether the data of the three British studies
(20,
21,
23), the American study
(25), and our study could be combined. All these studies included only Caucasians in the patient and comparison groups. The methodological features of these studies are provided in
Table 2. The testing of genetic heterogeneity included pairwise comparisons of the distributions of the MAOA-CA alleles in the five studies by means of the program CLUMP, version 1.1 (10,000 simulations)
(31). As 40 pairwise comparisons were needed to compare the patient and comparison groups in the five studies by gender, the significance level was adjusted by using Bonferroni correction (significant p<0.05/40=0.00125). Alternatively, we tested for genetic heterogeneity according to the method used by Rubinsztein and colleagues
(21), whereby they subdivided the MAOA-CA alleles into four categories: a2, a5, a6, and “rare alleles.” With respect to the VNTR (collapsed into long and short alleles) and RFLP alleles, genetic heterogeneity was tested by using chi-square statistics for contingency tables. These tests for genetic heterogeneity were performed separately for the patient and comparison groups by gender (12 comparisons; significant p<0.05/12=0.0042).
The Japanese data
(22) were excluded from the meta-analysis because 1) they revealed a different pattern of allelic association with bipolar affective disorder at the MAOA locus and 2) the allele distributions were not reported by gender. The German study
(24) could not be included in the meta-analysis because of the application of the haplotype relative risk method, i.e., the comparison groups were family based.
Results
Association Study
The most frequent genotypes of the MAOA-CA (i.e., 2-5, 2-6, 3-6, 5-6, and 6-6), the MAOA-VNTR (divided into long and short alleles), and the MAOA-RFLP (two alleles) markers were in Hardy-Weinberg equilibrium in both the patients (χ2=1.13, df=4, p=0.88; c2=0.09, df=2, p=0.96; c2=1.74, df=2, p=0.42, respectively) and comparison group (c2=0.45, df=4, p=0.98; c2=0.04, df=2, p=0.98; c2=0.04, df=2, p=0.98, respectively).
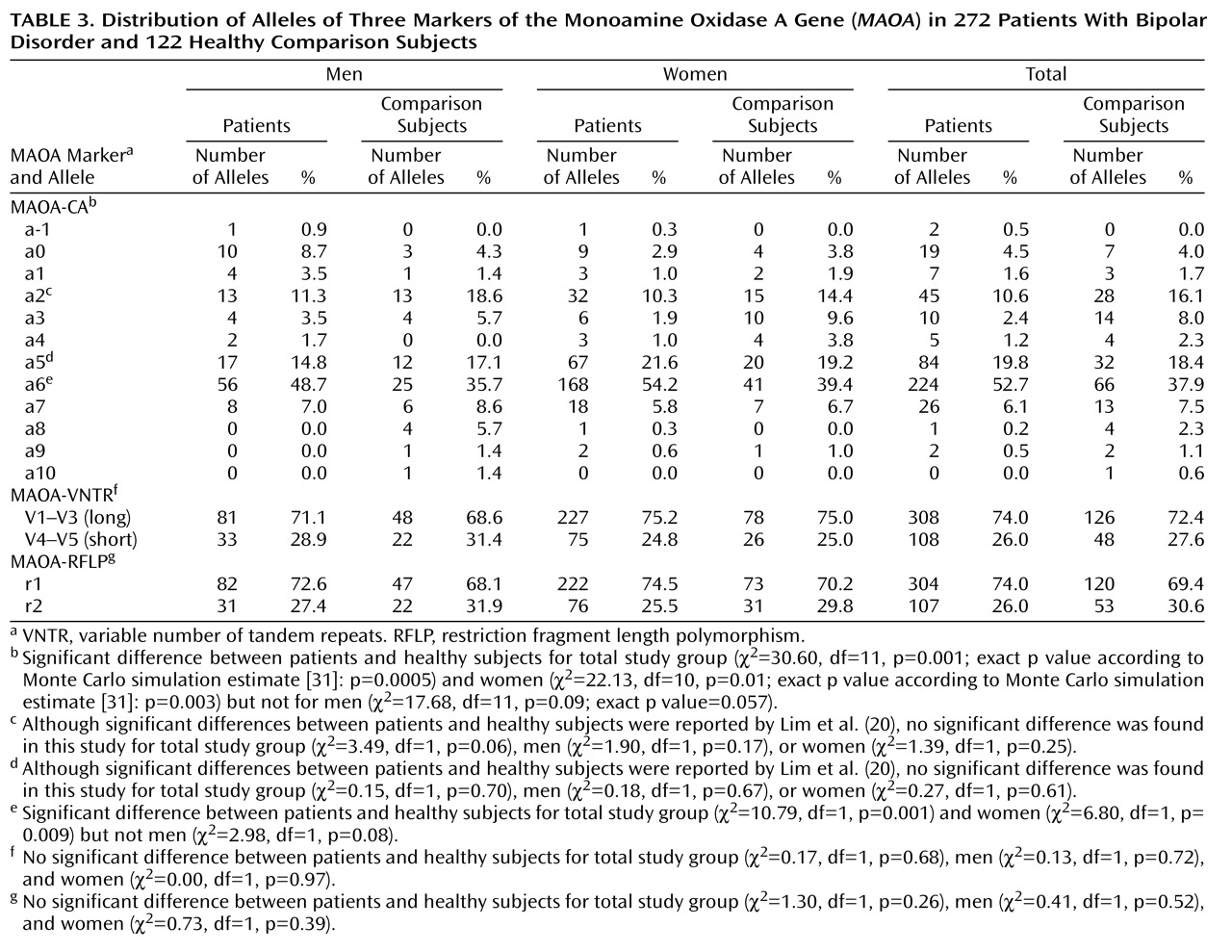
The results of bivariate analyses comparing the allelic distributions in the patient and comparison groups are presented in
Table 3. A significant difference in the allelic distribution of the MAOA-CA repeat was observed in the overall sample and in the women. These differences were mainly attributable to a higher frequency of the a6 allele in the bipolar subjects than in the healthy subjects; for the overall sample the odds ratio was 1.82 and the 99.17% confidence interval (CI) was 0.99–3.34, and for the female subjects the odds ratio was 1.82 and the 99.17% CI was 1.12–2.96.
In contrast, the allelic distributions of the collapsed VNTR and RFLP markers did not differ between the patients and healthy subjects of either gender.
Meta-Analysis of Available Data
None of the tests of genetic heterogeneity revealed evidence for significant differences across studies for any of the three markers in any combination of sex and disease status. Therefore, we pooled the data of the five available studies for meta-analysis.
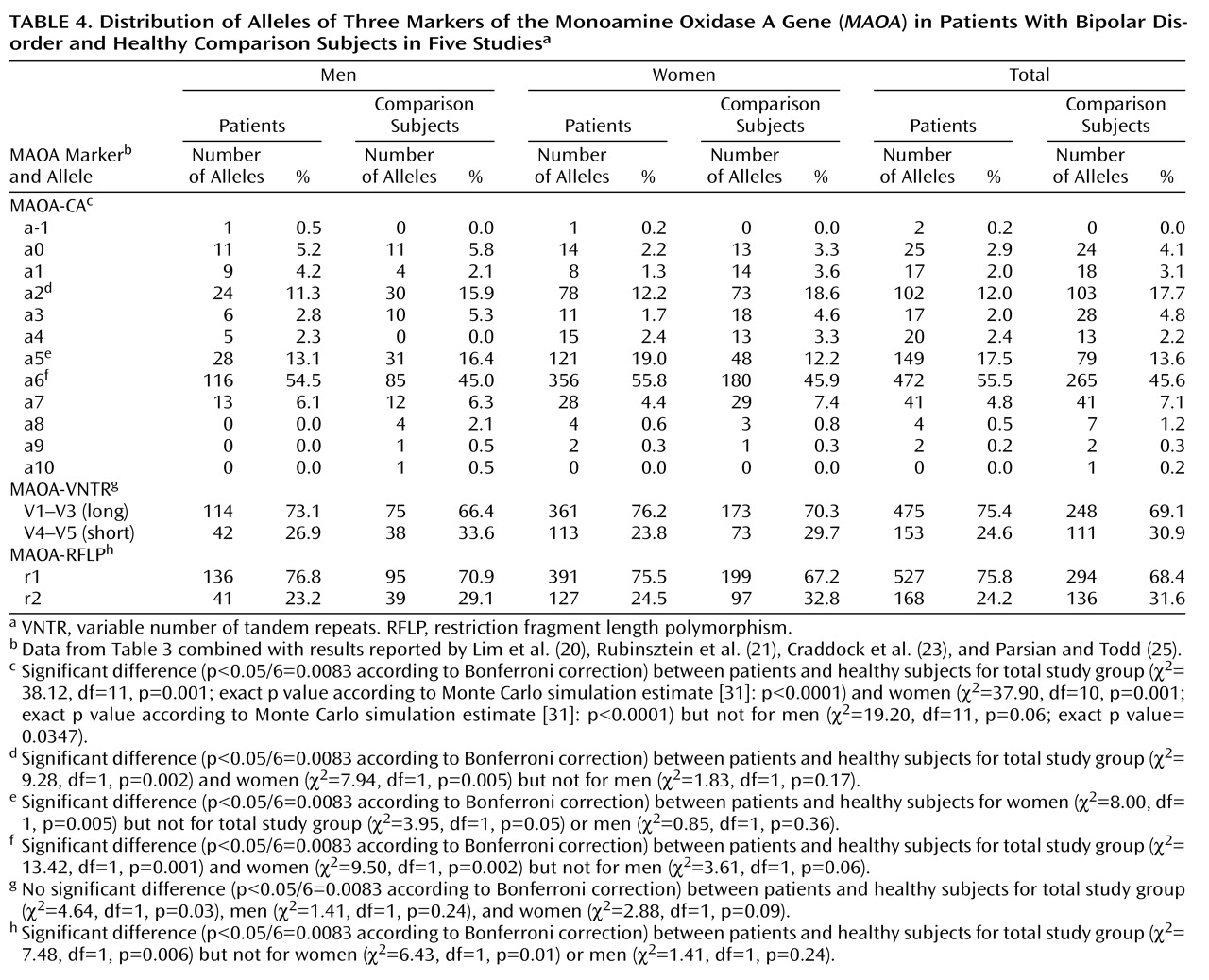
At the MAOA-CA locus, an association between the distribution of all alleles and bipolar disorder was found in the overall sample and in the women (
Table 4). The a6 allele (overall sample: odds ratio=1.49, 99.17% CI=1.12–1.98; women: odds ratio=1.49, 99.17% CI=1.06–2.01) and a5 allele (women: odds ratio=1.68, 99.17% CI=1.03–2.73) were more frequent among the bipolar patients than among the healthy subjects, whereas the a2 allele was less frequent among the patients (overall sample: odds ratio=0.63, 99.17% CI=0.42–0.94; women: odds ratio=0.61, 99.17% CI=0.38–0.97). However, among the men, alleles at the MAOA-CA locus were not significantly associated with bipolar disorder.
Pooled analysis also revealed an association between the r1 allele of the MAOA-RFLP marker and bipolar disorder in the overall sample (odds ratio=1.45, 99.17% CI=1.01–2.08), whereas no association was found at the MAOA-VNTR locus.
Discussion
Previous research has produced conflicting results regarding the association between three MAOA markers and bipolar disorder. In order to examine whether the associations could be further replicated in western European populations, we conducted what we believe to be the first replication study in French and Swiss populations. By collecting a sample approximately the size of the combined previous U.K. and U.S. studies, we hoped to minimize the risk of type II error.
Partial Replication in French-Swiss Data
In our sample of 272 bipolar patients and 122 screened healthy subjects, we replicated the finding of an association between bipolar disorder and the MAOA-CA marker in female subjects and the overall sample, as previously reported by Lim and colleagues
(20). However, in contrast to Lim and colleagues
(20), we found the a6 allele to be more strongly associated with bipolar disorder than the a5 allele. This discrepancy may be partially attributable to typing difficulties, as the distinction between the a5 and a6 alleles was difficult because their length differs by only two base pairs. Regarding the MAOA-VNTR and MAOA-RFLP markers, we did not replicate the association between these markers and bipolar disorder reported by Lim and colleagues
(20).
One crucial problem of association studies is the adjustment of the significance level when several polymorphisms with multiple alleles are tested. Although we chose the Bonferroni correction, which is usually considered as conservative
(35), there is still a risk that an observed positive finding is due to chance given the large number of potential candidate genes
(36). However, the goal of the present study was not to detect a new association between bipolar disorder and one of the thousands of potential candidate genes for brain disorders, but to replicate already reported positive findings obtained in other study populations.
Meta-Analysis of Available Data
Considering the different results obtained in the five studies that have examined the association between MAOA markers and bipolar disorder in Caucasians, we tested whether the observed allelic distribution differed significantly across populations. Analyses did not provide evidence for genetic heterogeneity for any of the three markers. The absence of genetic heterogeneity is remarkable since different diagnostic classification systems, diagnostic interviews, and inclusion criteria (for example, bipolar I only versus combined bipolar I and bipolar II) were used for the patient and comparison groups across studies.
Analysis of the combined data of the five available studies confirmed the association between the MAOA-CA polymorphism and bipolar disorder in women and the overall sample. The MAOA-RFLP marker was less strongly associated with bipolar disorder. It is unlikely that the positive findings in both the British and French/Swiss studies are merely due to chance, but their limitation is the use of an association study approach, which cannot rule out confounding by population stratification. However, the similarity of results across studies reduces the risk that the observed association is entirely explained by this artifact.
An important problem of meta-analyses is the risk that publication bias induces a weighting of positive results, since studies with negative data are more likely to remain unpublished. In our analysis we were able to include two published studies with negative findings, which suggests that negative findings are not a major impediment for publication in this field. As we are not aware of unpublished studies or theses on the association between bipolar disorder and
MAOA, we cannot use them to further estimate the effect of publication bias, as suggested by Glass et al.
(37). However, if the proposed statistical approach of Hedges and Olkin
(38) was adapted to our pooled data, unpublished studies including 1,452 genes of patients and 1,452 genes of comparison subjects with identical allelic distributions at the MAOA-CA locus (presence or absence of the a6 allele) would be necessary to overturn the conclusion of our meta-analysis (if we assume significance at p<0.0083, as used for our allelic analyses). It is unlikely that such a large number of unpublished data exist in this field.
Biological Context
Our finding of an association between
MAOA and bipolar disorder is compatible with the demonstration of abnormalities in serotonergic neurotransmission
(9–
12) and abnormal levels of MAO activity
(39–
42) in mood disorders.
The MAOA-CA polymorphism was more strongly associated with bipolar disorder than the other two
MAOA markers (MAOA-VNTR, MAOA-RFLP) according to our meta-analysis. This suggests that either the microsatellite or a nearby genetic variant may be functionally responsible for the susceptibility to bipolar disorder. However, it is unlikely that the microsatellite itself is functionally responsible for this susceptibility since it has not been demonstrated that a dinucleotide repeat could modify the expression of the gene into which it is inserted. The positive relationship between RFLP and bipolar disorder established in the meta-analysis is of particular interest since currently it is the only marker directly associated with MAOA activity
(19). In fact, Lim and colleagues
(20) demonstrated that a smaller number of bipolar patients than healthy subjects have an
Fnu4HI site in
MAOA, which is associated with high activity, therefore suggesting that bipolar patients have lower MAOA activity than healthy subjects.
Conclusions
Our findings, based on a combined French and Swiss sample as well as a meta-analysis, provide additional evidence for the existence of an association between the MAOA-CA polymorphism and bipolar disorder in females. However, in order to definitely exclude undetected population stratification as the source of the observed association, it is necessary to conduct replication studies that use the family association method with an “internal comparison group” (the haplotype relative risk method or the transmission disequilibrium test) and complementary approaches, such as the affected sib pair strategy. In addition, further analyses should attempt to identify clinically meaningful subtypes of bipolar disorder according to the presence or absence of the a2 and a6 MAOA-CA alleles.
Up to this time, several markers of enzymes implicated in the metabolism of neurotransmitters have been shown to be associated with bipolar disorder. In order to increase our understanding of the genetic mechanisms involved in its pathogenesis, further association studies with simultaneous assessment of several markers will be necessary to elucidate the combined effect of two and more “high-risk” alleles.