Several lines of evidence suggest that the prefrontal cortex, including Brodmann’s areas 8 to 10 and 44 to 47
(1,
2), is important to the pathology and treatment of schizophrenia. Neuropathological findings include low dendritic spine density on pyramidal neurons
(3) and low levels of synaptophysin (a presynaptic terminal protein)
(4–
6) in the prefrontal cortex of schizophrenia patients. In addition, patients with schizophrenia perform poorly on cognitive tasks, especially those involving short-term memory and attention, which are subserved by neural mechanisms intrinsic to prefrontal cortex circuitry
(7,
8).
Cholinergic afferents from the nucleus basalis of Meynert project to all layers of the cerebral cortex
(9), account for 70%–80% of cortical cholinergic input
(10–
12), constitute a major regulatory pathway of prefrontal cortical functioning
(9,
13,
14), and are important to higher cognitive processes
(12,
15). Specifically, cortical cholinergic inputs are hypothesized to mediate an individual’s ability to detect, select, and process sensory or associational information essential to cognition
(12). Lesions in the nucleus basalis of Meynert induce cortical cholinergic denervation
(16); decrease baseline levels of acetylcholine in the frontal cortex
(17); increase frontal cortical muscarinic cholinergic receptor levels, sensitivity, and mRNA
(18–
21); and impair learning and memory
(22,
23). Accordingly, dysfunction of cholinergic input from the nucleus basalis of Meynert to the prefrontal cortex may primarily or secondarily contribute to aberrant prefrontal cortical activation and the emergence of cognitive symptoms associated with schizophrenia and other neuropsychiatric disorders.
Because postsynaptic excitatory muscarinic cholinergic M
1 and M
4 receptors are the major cholinoceptive targets in the prefrontal cortex
(24), and hence essential to the circuitry between the nucleus basalis of Meynert and prefrontal cortex, we hypothesize that M
1 and M
4 receptors are primarily or secondarily altered in schizophrenia
(25). Moreover, on the basis of the putative high-affinity, antimuscarinic actions of typical and atypical antipsychotic drugs, such as thioridazine, clozapine, and olanzapine
(26–
29), and the commonly administered anticholinergic benztropine mesylate (used to treat extrapyramidal side effects [
30,
31]), we propose that M
1 and M
4 receptors are involved in the pharmacotherapeutics of schizophrenia. Therefore, using quantitative autoradiography, we measured the specific binding of the M
1/M
4 selective receptor antagonist [
3H]pirenzepine
(25,
32) in prefrontal cortex (Brodmann’s areas 8, 9, 10, and 46) from schizophrenia patients not treated or treated with benztropine and normal comparison subjects. Moreover, we performed preliminary studies of specific [
3H]pirenzepine binding in frontal cortex from rats given or not given antipsychotic drugs or benztropine.
Method
Human Tissue Studies
Postmortem human brain tissue was obtained during autopsy at the Victorian Institute of Forensic Medicine. The Human Ethics Committee of the Mental Health Research Institute of Victoria provided ethical approval for the collection of tissue. Specifically, prefrontal cortex (Brodmann’s areas 8, 9, 10, and 46) was collected from the left brain hemispheres of 17 patients with a diagnosis of schizophrenia and 20 subjects with no clinical history of psychiatric illness, neuroleptic exposure, or histopathological evidence of neurological disease (comparison subjects). Using a diagnostic instrument for brain studies
(33), a senior psychiatrist and psychologist extensively reviewed the case histories of subjects with schizophrenia and confirmed the diagnosis according to DSM-III-R criteria. For each subject with schizophrenia, the duration of illness (the time from first hospital admission to death), final recorded antipsychotic drug dose (converted to chlorpromazine equivalents
[34]), and use of benztropine was determined (
Table 1).
The age, postmortem interval, and sex of the subjects studied were determined (
Table 1 and
Table 2). The postmortem interval was taken as the time between witnessed death and autopsy. In cases where the death was not witnessed, tissue was collected only from subjects who had been seen alive within 5 hours of being found dead. In this instance, the postmortem interval was taken as the period starting half way between the last sighting of the subject while still alive and the time when he or she was found dead and ending at autopsy. In all cases, the cadavers were stored at 4°C within 5 hours of being discovered. After collection, the tissue was stored at –70°C until required (freezer time) (
Table 1 and
Table 2). To control for the effects of agonal state on the tissue collected, the pH of the brain tissue was determined as described previously
(35).
Serial sections (four 20-μm sections per subject) of Brodmann’s areas 8, 9, 10, and 46 were coronally cut at –20°C by using a cryomicrotome, thaw-mounted onto chrome-alum/gelatin-coated glass slides, and stored at –70°C until required. Selection of tissue blocks was standardized by using a set of standard landmarks
(36). Brodmann’s area 8 was taken as occupying the region posterior to Brodmann’s area 9 and anterior to Brodmann’s area 6, including the superior and middle frontal gyri immediately anterior to the superior precentral sulcus. Brodmann’s area 8 is wide on the superior surface, extending from the callosomarginal sulcus on the medial surface and narrowing laterally to the middle frontal gyrus. Brodmann’s area 9 was taken as being posterior to Brodmann’s area 10 and including the superior and middle frontal gyri. The medial surface of Brodmann’s area 9 was defined by the callosomarginal gyrus, while laterally it stopped ventrally in the region of the inferior frontal sulcus. Brodmann’s area 10 included the anterior quarter of the superior and middle frontal gyri on the convexity of the hemisphere, not extending medially as far as the callosomarginal gyrus. Inferomedially, Brodmann’s area 10 was demarcated by the superior rostral sulcus. Finally, Brodmann’s area 46 included the middle third of the middle and the anterior part of the inferior frontal gyri at the transition to the orbital surface.
Rat Tissue Studies
For the antipsychotic drug studies, 6-week-old male Sprague-Dawley rats (100–150 g, N=3 per group) were treated orally for 1 month (short-term) or 3 months (long-term) with vehicle (H2O; comparison rats), haloperidol, clozapine, chlorpromazine, or thioridazine. The haloperidol was initially dissolved in a minimal volume of 10% acetic acid (<0.001% final) and diluted with distilled water. Clozapine, chlorpromazine, and thioridazine were similarly dissolved in 0.1M HCl (<0.02 nM final), distilled water, and 10% ethyl alcohol (<0.001% final), respectively, and diluted with distilled water.
For the benztropine studies, 6-week-old male Sprague-Dawley rats (100–150 g, N=3 per group) were treated for 1 month with vehicle or benztropine. The benztropine was dissolved in distilled water and administered orally at a dose of 0.1 mg·kg
–1·day
–1 (37).
To ensure that each rat received an appropriate amount of drug, the amount of each drug in drinking water was modified according to the measured daily water intake and weekly body weight of the subjects. At the completion of drug administration, the rats received drug-free drinking water for 72 hours before being killed by decapitation. The brains were rapidly removed, frozen in isopentane on dry ice, and stored at –70°C. Serial sections of rat frontal cortex (six 20-μm sections per subject) were coronally cut (so as to correspond to plates 5–7 from Paxinos and Watson
[38]) at –20°C by using a cryomicrotome, thaw-mounted onto chrome-alum/gelatin-coated glass slides, and stored at –70°C until required.
In Situ [3H]Pirenzepine Binding With Autoradiography
The in vitro [
3H]pirenzepine binding studies with autoradiography were performed as previously described
(25). Briefly, mounted tissue sections were incubated with 15 nM [
3H]pirenzepine (specific activity: 72 Ci/mmol) (New England Nuclear Life Science Products, Boston) in the absence (total binding; human studies: two sections, rat studies: three sections) or presence (nonspecific binding; human studies: two sections, rat studies: three sections) of 1 μM quinuclidinyl xanthene-9-carboxylate hemioxilate tetraoxilate (Research Biochemicals International, Natick, Mass.). The sections were washed, air dried, and apposed to Amersham [
3H]-Hyperfilm (Amersham Pharmacia Biotech UK Ltd., Little Chalfont, Buckinghamshire, U.K.) with Amersham
3H micro-scales (Amersham Pharmacia Biotech UK Ltd., Little Chalfont, Buckinghamshire, U.K.) for up to 5 weeks. The images were analyzed by using a microcomputer imaging device and image analysis system (Imaging Research, Inc., St. Catherine’s, Ont., Canada). The results are expressed as femtomoles per milligram of estimated tissue equivalents. Specific radioligand binding was calculated as total binding minus nonspecific binding.
Statistical Analysis
One-way analyses of variance with least significant difference post hoc tests were used to compare the mean radioligand binding, age, postmortem interval, freezer time, and pH of the schizophrenia patients treated with benztropine, the untreated patients, and the comparison subjects, as well as levels of receptor binding in the frontal cortex of the rats given antipsychotic drugs and comparison rats for each length of administration. The relationships between receptor binding in human prefrontal cortex and subject age, postmortem interval, freezer time, pH, and, where relevant, duration of illness and final recorded antipsychotic drug dose were assessed by using Pearson product moment correlation coefficients, derived by using an assumed straight-line fit. A one-way analysis of covariance (ANCOVA) with least significant difference post hoc tests were used to determine whether freezer time was a confounding variable influencing the apparent differences between radioligand binding in the different groups. An unpaired t test was used to compare the mean levels of receptor binding in the frontal cortices of the rats given benztropine and the comparison rats.
Discussion
We found low receptor-bound [
3H]pirenzepine levels in prefrontal cortex from patients who had schizophrenia. This low [
3H]pirenzepine binding is consistent with the low binding of [
3H]pirenzepine in the caudate-putamen
(39) and hippocampal formation
(25) that was found in earlier studies.
Antipsychotic drug treatment is nearly ubiquitous in patients with schizophrenia, and antiparkinsonian drugs such as benztropine are commonly administered for the control of extrapyramidal side effects. Subsequently, neuroleptic agents are potential confounders in many neurobiological studies of schizophrenia
(40).
Although we are unable to unequivocally exclude a causal relationship between antipsychotic drug treatment and low receptor-bound [
3H]pirenzepine levels in the prefrontal cortex of schizophrenia patients, preliminary studies suggest that treatment with antipsychotic drugs did not confound the measurement of receptors in schizophrenia. In our rat studies, treatment with typical or atypical antipsychotic drugs tended to increase or have no effect on the levels of receptor binding. As the drugs presently considered are putative muscarinic antagonists
(41,
42), they would not be expected to directly down-regulate muscarinic receptor levels. However, the possibility that receptor levels are reduced through more complex mechanisms in human brains than in rat brains cannot be excluded.
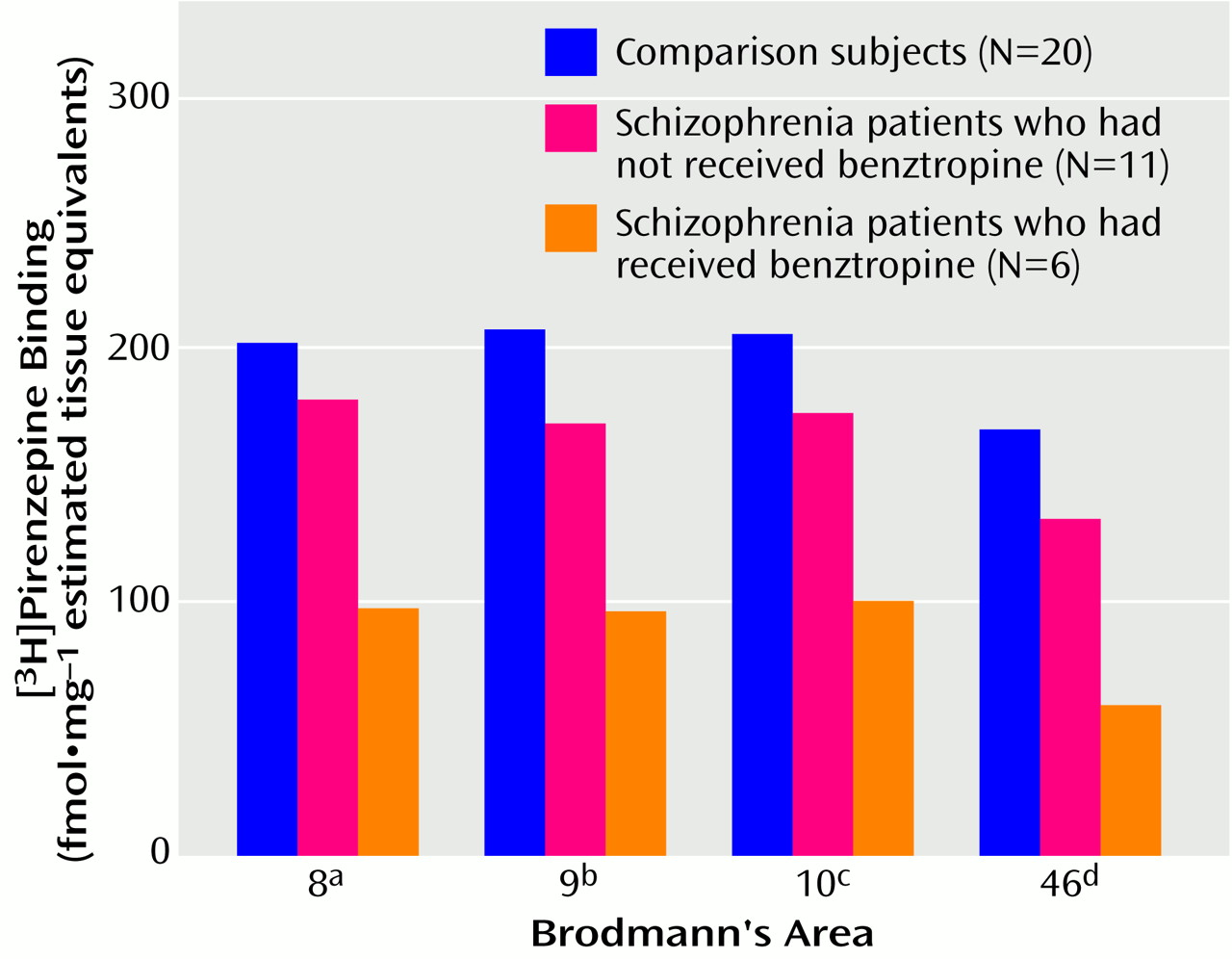
In regard to the benztropine studies, while the rat data indicate no effect of drug administration on receptor-bound [
3H]pirenzepine levels, a consideration of patient drug treatment revealed a more complex picture. The difference in radioligand binding from comparison values was greater for the schizophrenia patients who had been treated with benztropine than for those who had not received benztropine. However, the radioligand binding of the non-benztropine-treated patients was significantly lower in Brodmann’s areas 9 and 46, and nonsignificantly lower in Brodmann’s areas 8 and 10, than in the comparison subjects. We previously reported
(39) that the lower-than-normal [
3H]pirenzepine binding in caudate-putamen from schizophrenia patients was greater for patients treated with benztropine than for non-benztropine-treated patients. Moreover, as in the present findings, the density of [
3H]pirenzepine binding to caudate-putamen for the non-benztropine-treated schizophrenia group was also lower than that for the comparison group. With the limitations of our drug studies, there are a number of possible explanations for the present findings
(39). One possibility is that although treatment of patients with benztropine may cause a down-regulation of muscarinic receptor binding, benztropine is not the sole cause of the reduced receptor binding. It is important to note that in the event that altered radioligand binding reflects altered receptor levels, a muscarinic receptor antagonist such as benztropine would not be expected to directly down-regulate muscarinic receptor levels. However, a drug-induced decrease in receptor levels through other mechanisms cannot be excluded. A second possibility is supported by the rat studies, which indicate that benztropine treatment does not induce a decrease in muscarinic receptor binding. Perhaps the observed relationship between lower radioligand binding levels in the human prefrontal cortex and treatment with benztropine reflects a propensity to develop extrapyramidal side effects and a concomitant need for treatment after alteration of muscarinic receptor activity
(39).
While we can only speculate on the import of the present findings, it seems reasonable to suggest that the muscarinic system may be involved in the pathology and/or pharmacotherapeutics of schizophrenia. First, in the event that altered muscarinic receptor binding in schizophrenia is influenced by antipsychotic or antiparkinsonian drug treatment, this may be relevant to understanding the mechanisms of current and future pharmacotherapeutic strategies. Second, a possible primary change of cortical muscarinic receptors in the pathology of schizophrenia may be subsequent to a pre- or posttranscriptional abnormality. Studies of M
1 and M
4 receptor transcripts may be useful in determining aberrant receptor gene expression. Finally, decreased radioligand binding may reflect a down-regulation in M
1 and/or M
4 receptor levels after an increase in cholinergic efflux following overactivity of the cholinergic basal forebrain system. If this is the case, on the basis of the putative dopaminergic modulation of transmission in the basal forebrain mediated by γ-aminobutyric acid (GABA), a hypercholinergic model may be relevant to the dopamine hypothesis of schizophrenia
(43–
45). It is interesting that dopamine receptor stimulation in the nucleus accumbens has been shown to reduce the activity of inhibitory GABA-ergic efferents to the basal forebrain and increase the excitability of cholinergic basal-forebrain corticopetal neurons
(46–
48). Thus, increased mesolimbic dopaminergic transmission, deemed central to schizophrenia
(49), may induce increased cortical cholinergic efflux and down-regulate postsynaptic M
1 and M
4 receptor levels and the binding presently measured. This model is consistent with the current understanding of the actions of antipsychotic drugs such as haloperidol. Haloperidol normalizes dopamine-mediated transmission in the nucleus accumbens, possibly through dopamine D
2 receptors, and hence reduces the excitability of cortical cholinergic input
(48,
50). Accordingly, the present rat studies suggest that long-term treatment with haloperidol induces up-regulation of levels of [
3H]pirenzepine-labeled receptors in the rat frontal cortex.
In conclusion, these studies indicate an abnormality of the muscarinic receptor system in the prefrontal cortex in schizophrenia. Our earlier findings of low muscarinic receptor binding in the caudate-putamen
(39,
51) and hippocampal formation
(25) in schizophrenia suggest that the abnormalities presently measured might not be limited to the prefrontal cortex. Although the interpretation of the data presented here remains tentative, these findings may be important in elucidating the cascade of events involved in the abnormal neurobiology of schizophrenia, and they may provide insight into current pharmacological interventions that could prove useful to the development of new strategies for the treatment of schizophrenia.