Neuroanatomical changes in schizophrenia are well established
(1), but their genetic and neurodevelopmental origins are not well understood. Genetically at-risk children present an excellent research opportunity to study premorbid vulnerability markers and developmental brain changes, especially during the crucial period preceding disease onset. Genetically at-risk children and children who later develop schizophrenia have significant neurological, psychological, and social developmental differences from normal subjects
(2). Specifically, attention deficits, minor neuromotor deficits, and neuroanatomical differences such as a smaller amygdala and smaller hippocampi have been observed in young at-risk relatives of patients with schizophrenia. In addition, at-risk relatives exhibit deficits in language development such as poor speech comprehension and verbal memory
(3–
6). Disturbances in thought and language are core features of schizophrenia
(7–
9), and a faulty development of the language system has been proposed as a causal mechanism for schizophrenia
(8).
The superior temporal gyrus, typically larger on the left in right-handed individuals, includes Heschl’s gyrus and the planum temporale and is believed to be a major anatomical substrate for speech, language, and communication. Penfield and Perot
(10) observed psychosis-like symptoms with electrical stimulation of the superior temporal gyrus. Neurological injuries to the superior temporal gyrus such as epilepsy, stroke, or tumors cause disturbances of thought, hallucinations, and delusions similar to those seen in schizophrenia
(11). The superior temporal gyrus has wide connections to temporolimbic areas, including the hippocampus, amygdala, entorhinal cortex, thalamus, and neocortical association areas in the prefrontal and parietal cortices
(12). Although not well understood, the wide and complex connections seem to integrate these brain regions, which are essential for information processing and thought.
The superior temporal gyrus, with a clear role in thought and language processes, is appropriately a major area of interest in schizophrenia research. The widely replicated superior temporal gyrus volume reduction in schizophrenia in relation to symptoms appears to be relatively disease specific, seems to be present at illness onset, and may progress during the course of the illness
(13–
21). However, it is unclear whether a smaller superior temporal gyrus reflects a premorbid susceptibility trait or a result of persistent psychosis. Studies of nonpsychotic at-risk offspring of parents with schizophrenia allow us to investigate this question. In this preliminary study, we used magnetic resonance imagining (MRI) to examine the superior temporal gyrus in a group of at-risk offspring. Our hypothesis was that the superior temporal gyrus would be smaller in at-risk subjects than in comparison subjects.
Method
Twenty-nine at-risk offspring (15 males and 14 females, mean age=14.9 years, SD=3.4) who had one parent suffering from schizophrenia or schizoaffective disorder and 27 healthy comparison subjects (14 males and 13 females, mean age=16.9 years, SD=5.7) with no family psychiatric history were recruited from the same community. We did not match parental socioeconomic status because the at-risk group, with a parent suffering from schizophrenia, would be expected to have lower socioeconomic status. Matching on this variable would very likely remove illness-related variance. The University of Pittsburgh Institutional Review Board approved the study. All subjects and their parents or guardians provided written consent and assent, when appropriate, after full description of the study. Subjects 14 and younger were evaluated with the Schedule for Affective Disorders and Schizophrenia for School-Age Children
(22). Parents and subjects 15 or older were evaluated with the Structured Clinical Interview for DSM-IV
(23). Any subject from either group with a lifetime history of psychosis, neurological disorders, chronic medical or psychiatric illnesses, current substance use disorder, psychotropic treatment, or any contraindication for MRI study was excluded.
For MRI acquisition and processing, 124 T
1-weighted 1.5-mm coronal slices, without interslice gap, were obtained with a 1.5-T GE scanner (GE Medical Systems, Milwaukee); three-dimensional spoiled gradient recall acquisition coronal, matrix=256×256×192, field of view=24 cm, TR=25 msec, and TE=5 msec. Scans were reviewed to exclude structural abnormalities. Intracranial volume was measured with NIH IMAGE
(24). The superior temporal gyrus (gray plus white) was manually traced by one of the authors blind to the clinical data (R.R.) (intrarater reliability=0.98 for the left superior temporal gyrus and intrarater reliability=0.99 for the right superior temporal gyrus) using BRAINS2
(25) on a resampled three-dimensional Talairach image. The methodology of superior temporal gyrus measurement has been described previously
(16,
18).
Differences in age, IQ, and intracranial volume were examined by two-tailed unpaired t tests. Multivariate analysis based on the general linear model was conducted with group status (at-risk subjects versus healthy comparison subjects), intracranial volume, and age as predictor variables and superior temporal gyrus measures as dependent variables. We examined age-related differences using partial correlations between age and superior temporal gyrus volumes controlling for intracranial volume. Parental socioeconomic status was not used as a covariate because the grouping was based on parental illness. All tests were two-tailed, and the significance level was set at p≤0.05.
Results
The two groups were not significantly different in age (t=–1.57, df=54, p=0.12), IQ (t=–1.28, df=54, p=0.21), handedness (25 right-handed, one left-handed, and three ambidextrous at-risk subjects and 23 right-handed, one left-handed, one ambidextrous, and two unknown-handed healthy comparison subjects), and gender distribution (χ2=0.03, df=1, p=0.85). The parental socioeconomic status was different between the groups (t=4.75, df=49, p<0.001), which would be expected because one parent of each at-risk subject was suffering from schizophrenia.
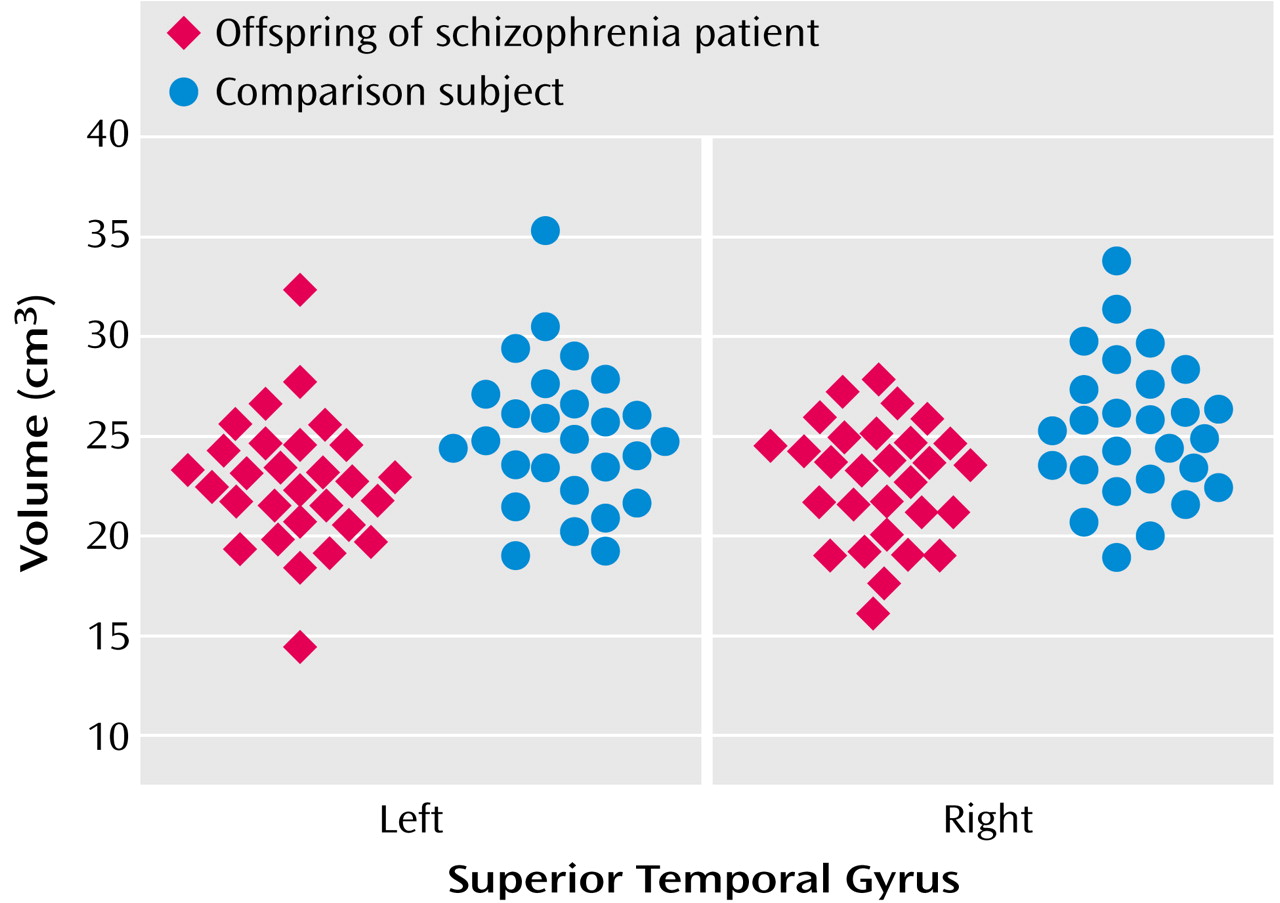
The intracranial volume of at-risk subjects (mean=1408.65 cc, SD=174.17) was not significantly different from that of healthy comparison subjects (1473.94 cc, SD=155.73) (t=1.48, df=54, p=0.15). Analyses of variance based on the general linear model showed significant differences in volume between the groups in the right superior temporal gyrus (for at-risk subjects, mean=22.8 cc, SD=2.98; for healthy comparison subjects, mean=25.3 cc, SD=3.5) (F=8.48, df=1, 56, p=0.005) and the left superior temporal gyrus (for at-risk subjects, mean=22.7 cc, SD=3.3; for healthy comparison subjects, mean=25.0 cc, SD=3.6) (F=6.49, df=1, 56, p=0.02) (
Figure 1). Similar analysis with IQ as an additional covariate did not change the results (left superior temporal gyrus: F=5.87, df=1, 55, p=0.02; right superior temporal gyrus: F=5.96, df=1, 55, p<0.02).
Left superior temporal gyrus volume showed a significant inverse correlation with age in healthy comparison subjects (partial r=–0.40, df=24, p=0.04) but not in at-risk subjects (partial r=–0.21, df=26, p=0.28).
Discussion
To our knowledge, this is the first report of reduced superior temporal gyrus volume in young offspring genetically at risk for schizophrenia. Superior temporal gyrus volume reductions in these nonpsychotic at-risk subjects suggest that abnormal development rather than degeneration of the superior temporal gyrus is associated with schizophrenia and that such deviant development may be mediated by familial/genetic factors. The absence of the age-related reduction in the superior temporal gyrus in at-risk subjects may be related to a failure of either the normative pruning process that occurs during adolescence or the synaptic/axonal proliferation that occurs earlier in development
(26).
Schizophrenia susceptibility genes may influence the integrity of brain regions responsible for thought and language processes and may account for the premorbid language abnormalities in at-risk individuals. Variations in genetic threshold, brain maturational processes such as neuronal pruning and myelination, environmental stressors, and compensatory mechanisms may explain occurrence of or protection from schizophrenia spectrum pathology in genetically vulnerable individuals.
Other structures in the hierarchy of information processing that are interconnected to the superior temporal gyrus, such as the hippocampus, thalamus, or frontal lobe, may also be developmentally impaired in at-risk subjects. For example, the hippocampus has been found to be smaller in patients with schizophrenia as well as at-risk subjects, and hippocampal sclerosis is known to reduce blood flow in the superior temporal gyrus, in association with an impairment of language function
(27–
31).
The relatively small number of subjects in our study points to a need for an independent replication. The small size of the study group may be the cause for the lack of expected left-greater-than-right asymmetry. This preliminary cross-sectional group comparison will be followed up with longitudinal studies to assess the predictive value and age-related changes in the superior temporal gyrus. Future studies ideally should include subjects carefully matched for age and sex and should incorporate periodic longitudinal clinical and imaging evaluations. Use of other approaches like functional imaging, diffusion tensor imaging, and spectroscopy will help us to understand the physiology of the superior temporal gyrus and other brain structures involved in schizophrenia and help clarify the altered development of functional neuroanatomy and physiology in at-risk individuals.