Deficits in smooth pursuit eye movements are among the most reproducible physiological abnormalities associated with schizophrenia
(1). A recent study from our laboratory
(2) revealed hyperactivation of the hippocampus during the task in subjects with schizophrenia compared with subjects without schizophrenia. This finding is consistent with neuropathological and neurophysiological evidence for inhibitory dysfunction in the hippocampus in schizophrenia
(3).
Nicotine temporarily normalizes smooth pursuit eye movement defects in schizophrenia
(4–
8). Olincy et al.
(4) reported a decrease in intrusive anticipatory (leading) saccades with nicotine, consistent with improved inhibitory function. Several studies of hippocampal dysfunction in schizophrenia implicate nicotinic cholinergic systems
(9–
11). Activation of nicotinic receptors on hippocampal inhibitory neurons
(12) may be an underlying mechanism of the response to nicotine.
The current study used functional magnetic resonance imaging (fMRI) to assess brain hemodynamic response associated with improved smooth pursuit eye movements following nicotine administration in subjects with schizophrenia. Specifically, we hypothesized that nicotinic cholinergic stimulation would be associated with decreased hippocampal activity, reflecting normalization of previously observed hyperactivity during pursuit eye movements in schizophrenia.
Method
Nine subjects with DSM-IV schizophrenia (seven with paranoid type, one with undifferentiated type) or schizoaffective disorder, depressed type (one subject) participated. The study group consisted of seven men and two women; their mean age was 34.6 years (SD=10.0). Five subjects were smokers. One subject was neuroleptic naive, two were taking first-generation neuroleptics, and seven were taking second-generation neuroleptics.
Subjects who smoked arrived 2 hours before scanning to ensure abstinence. The mean duration of self-reported abstinence before scanning was 9.2 hours (SD=6.7). Minnesota Withdrawal Scale
(13) scores were not significantly different between abstinent smokers and nonsmokers (t=–0.87, df=7, p<0.42). All subjects were volunteers and provided written, informed consent.
fMRIs were obtained at 1.5 T while subjects performed a constant velocity visual smooth pursuit task
(2). Subjects performed two runs of the pursuit task, each consisting of a 10-second equilibration period followed by four cycles of 25-second pursuit task/25-second rest, while gradient-echo planar imaging data were collected (TR=2500, TE=50, 642 matrix, 240-mm
2 field of view, 20 axial slices, 6-mm thick, 1-mm gap).
Subjects were then removed from the scanner and given either nicotine or placebo. Following drug administration, subjects were scanned while performing two additional runs of the task. Subjects returned the following week to repeat the experiment with either placebo or nicotine, whichever they had not received previously.
Subjects received nicotine or placebo in a randomized, counterbalanced design. Smokers were given 6 mg and nonsmokers were given 4 mg of nicotine administered as polacrilex gum. The placebo consisted of similar tasting gum. Subjects chewed vigorously for 10 minutes.
Data were analyzed with SPM 99
(2,
3). Following realignment, normalization to stereotactic space, and smoothing (4-mm full width at half maximum), data were modeled with a simple boxcar convolved with a hemodynamic response function and assessed according to the general linear model. A random effects analysis was implemented by entering estimates of each individual’s response into a second-level paired t test, contrasting for higher values after nicotine than after placebo or lower values after nicotine than after placebo.
Regions previously shown to be involved in pursuit eye movements and areas showing differential activity during the task in schizophrenia
(2,
14) were examined by evaluating the mean blood-oxygen-level-dependent (BOLD) response over all voxels in each region
(15). Regions included 10-mm-diameter spheres placed in the frontal, supplementary, and parietal eye fields, anterior and posterior cingulate gyrus, precuneus, MT/V5, and hippocampus. Data for each region were thresholded at p<0.05, with Bonferroni correction for the number of regions evaluated.
A Fourier analysis of time-series data identified task-correlated voxel intensity fluctuations near the optic nerve
(16). The square of the magnitude of the fast Fourier transform was calculated by using spatially normalized data from a 10×10×2-voxel region of interest including the eye and optic nerve. According to the task paradigm and volume acquisition time, the expected intensity fluctuations caused by eye movement were between 0.16 and 0.20 Hz. We calculated the area under the curve of this portion of the spectrum and compared the difference between before placebo and after placebo with the difference between before nicotine and after nicotine (a session-by-drug interaction) using a paired t test.
Results
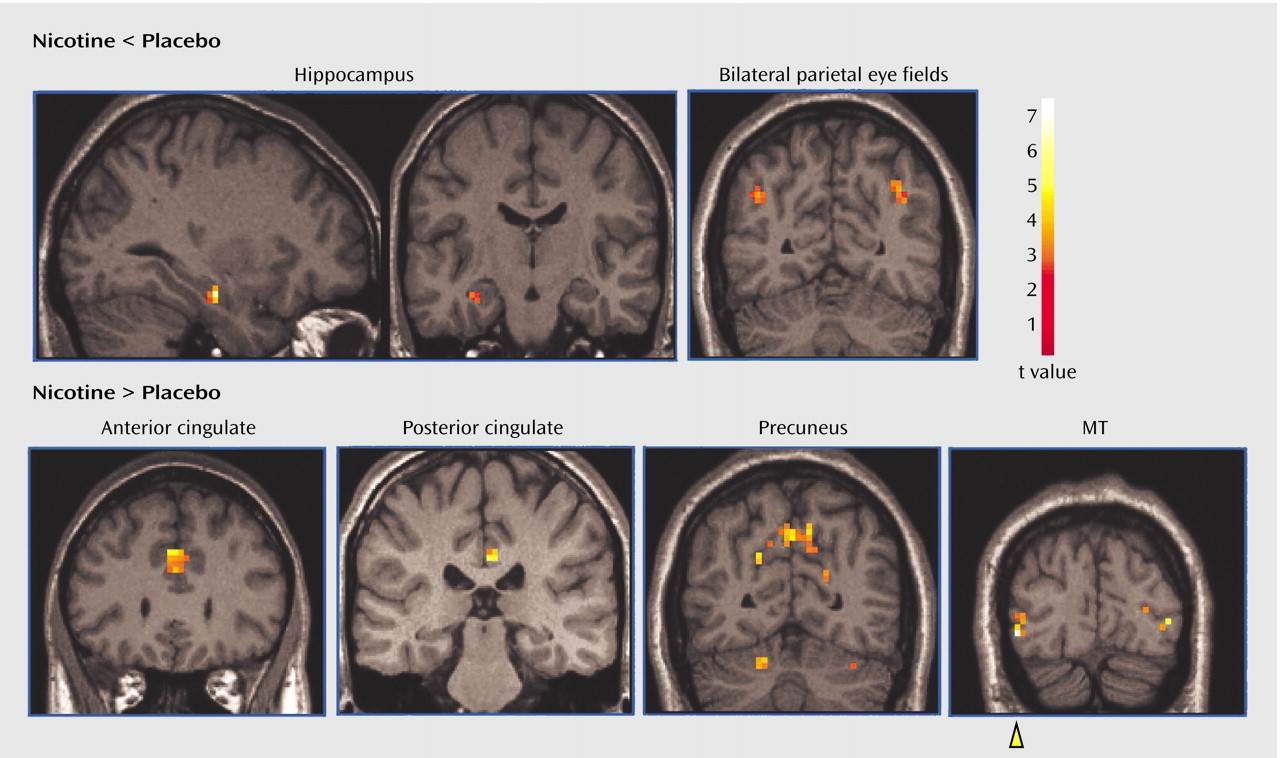
Compared with placebo, nicotine administration during the smooth pursuit eye movement task was associated with less activity in the right hippocampus (t=5.20, df=8, p<0.002) and bilateral parietal eye fields (right: t=3.06, df=8, p<0.03; left: t=2.89, df=8, p<0.04) (
Figure 1). Individually, all subjects demonstrated decreased activity (measured as percent BOLD signal change relative to the global mean) in the hippocampus (mean difference=–0.178, SD=0.103). Decreased activity was observed in the parietal eye fields in seven subjects (mean difference=–0.350, SD=0.344).
Nicotine administration was associated with increased activity in both the anterior (t=3.08, df=8, p<0.04) and posterior (t=3.61, df=8, p<0.02) cingulate gyrus, the precuneus (t=3.59, df=8, p<0.02), and area MT/V5 (t=4.23, df=8, p<0.009) (
Figure 1). BOLD responses were higher during the nicotine condition compared with placebo in all subjects in the anterior cingulate (mean difference=0.261, SD=0.254), posterior cingulate (mean difference=0.332, SD=0.276), and right area MT/V5 (mean difference=0.693, SD=0.491). Eight subjects demonstrated increased activity in the precuneus (mean difference=0.607, SD=0.507) during nicotine administration.
Nicotine administration was associated with a small improvement in pursuit performance (area under the curve change from 342 to 351), while placebo administration was associated with a decline in performance (area under the curve change from 338 to 306). A significant session-by-drug effect (the difference between before and after nicotine compared with the difference between before and after placebo) was observed (t=2.1, df=8, p<0.03).
Discussion
The most significant difference observed during smooth pursuit following administration of nicotine was decreased activity of the hippocampus. This change is consistent with the hypothesis that activation of nicotinic cholinergic receptors in the hippocampus normalizes previously observed hyperactivity in this region in schizophrenia
(2). The nicotine-associated decrease in activity in the parietal eye fields may also be consistent with improved inhibitory function. This region is involved in the reflexive exploration of the visual field, i.e., generating automatic saccades to objects that appear in the periphery
(18,
19). Reduced parietal eye field activation may be associated with decreased generation of unwanted saccades during the smooth pursuit task.
Posterior motion processing areas (V5 or MT) showed the most significant increase in task-associated activity in the nicotine condition. This may reflect nicotinic receptor binding in these regions or subcortical visual relays such as the lateral geniculate nucleus, which is known to have high nicotinic receptor concentrations
(20).
Nicotine-associated increases in activity were also observed in the precuneus and posterior cingulate gyrus, regions likely involved in attentive tracking and monitoring eye movements, respectively
(21,
22). The rostral area of the anterior cingulate shown to be more active in the nicotine condition may play a role in monitoring conflicts in information processing
(23). The cingulate gyrus contains a high density of nicotinic receptors and is richly interconnected with the hippocampus
(19).
Taken together, these results suggest that nicotine may improve smooth pursuit eye movement task performance in schizophrenia both by enhancing activity in brain regions involved in attending to moving stimuli and by improving inhibitory function, resulting in decreased activity in brain regions associated with the generation of intrusive saccadic eye movements.