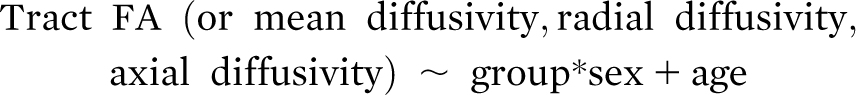Anxiety disorders emerge in childhood and early adolescence and are among the most common psychiatric illnesses in this age group (
1,
2). Youths with anxiety disorders, such as generalized, separation, and social anxiety disorder, suffer from significant impairment and face increased risk for additional stress-related psychopathology (
3). A more in-depth understanding of the pathophysiological processes underlying childhood anxiety disorders will provide rationales for novel treatment targets in children with anxiety disorders. Our work and that of others has identified alterations in white matter (WM) pathways in children and adults with anxiety disorders (
4–
9), which is of interest because WM plays an important role in mediating optimal neuronal communication. It is important to recognize that WM is modifiable, as preclinical and clinical studies demonstrate that WM and myelin dynamically change with development and in response to environmental factors, including behavioral and pharmacological interventions (
10–
14).
Prefrontal-limbic dysfunction is thought to underlie the pathophysiological processes associated with anxiety disorders (
15). In a previous study (
4), we found anxiety disorder–related alterations in the WM tract that connects prefrontal cortical regulatory regions to limbic regions (i.e., the uncinate fasciculus [UF]) in youths with anxiety disorders. Of particular interest is that these UF alterations appeared to occur selectively in boys and not girls. In further support of this finding, our nonhuman primate studies in anxious young monkeys showed a similar sexually dimorphic effect (
16). The absence of apparent UF alterations in girls does not obviate the possibility of prefrontal cortical–limbic involvement in pathological anxiety. The UF fractional anisotropy (FA) effects could be present in girls but less robust than in boys and, therefore, not detectable in the sample size analyzed in our previous study. Also, it is possible that boys and girls differ in the prefrontal cortical–limbic mechanisms that underlie the expression of pathological anxiety. In addition to the sexually dimorphic anxiety disorder–related effects in UF WM, lower FA was observed in other brain regions in both boys and girls with anxiety disorders, including regions such as the corpus callosum, inferior fronto-occipital fasciculus, and internal capsule (
4). Only one study has examined WM microstructural integrity in adolescents with anxiety disorders (
6). While sexually dimorphic effects were not reported, that study in adolescents with generalized anxiety disorder also found FA reductions in the UF as well as in the inferior fronto-occipital fasciculus, inferior longitudinal fasciculus, and corona radiata.
Given the small number of youths in whom the relation between WM integrity and anxiety disorders has been examined, we performed a mega-analysis across our studies to maximize the sample size and enhance confidence in the findings. A particular focus was to further explore the extent to which anxiety disorder–related WM alterations are sexually dimorphic. Understanding sex-related differences in the pathophysiology of anxiety disorders may be helpful in explaining the increased risk faced by girls, as with puberty, girls are twice as likely as boys to develop anxiety disorders (
17). The present analysis includes data from the two largest cross-sectional diffusion tensor imaging studies of pediatric anxiety disorders (
4,
18) in addition to a newly collected sample, for a total of 163 children with anxiety disorders and 132 healthy volunteers (total N=295). Medication-free children with current diagnoses of generalized, separation, and/or social anxiety disorders were studied. We utilized tractography and voxel-wise approaches to assess anxiety disorder–related alterations in WM microstructure.
Discussion
In this mega-analytic study comparing preadolescent children with anxiety disorders to healthy volunteers, we combined data from two published samples from our laboratories (
4,
18) with data from a new sample of 97 children. Our aim was to increase confidence in the previous findings by maximizing the sample size for exploring the relationships among WM microstructural alterations, anxiety disorders, and sex in preadolescent children. We found 1) reduced UF FA in boys with anxiety disorders relative to healthy volunteer boys, consistent with our previous work (
4); 2) beyond the UF, more broadly distributed WM reductions in boys with anxiety disorders; and 3) no evidence for anxiety disorder–related WM changes in preadolescent girls.
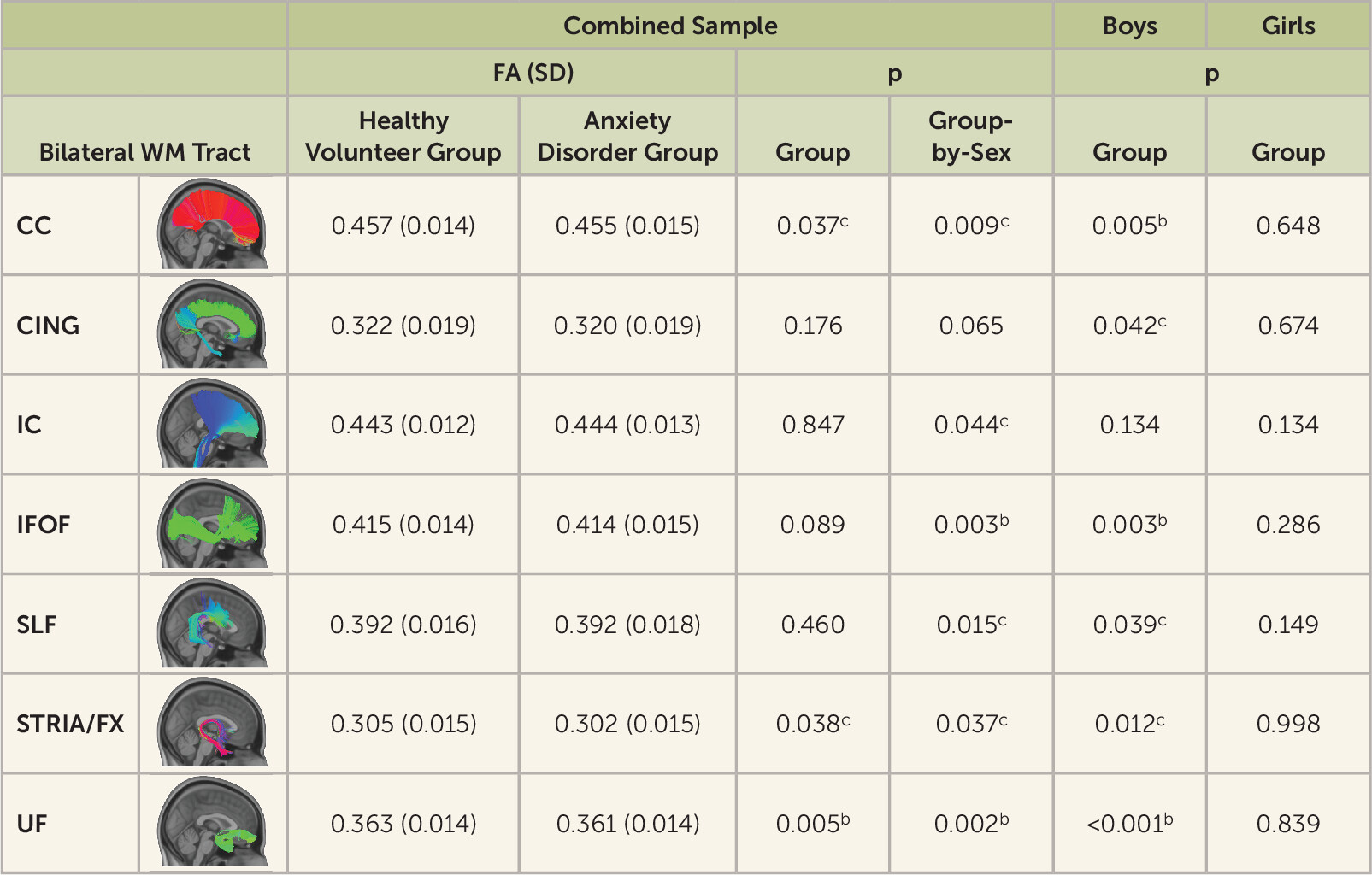
Findings from the tract-based and voxel-wise analyses demonstrated significantly decreased UF FA in boys with anxiety disorders and not girls with anxiety disorders. Anxiety disorder–related FA reductions were also observed in other WM tracts in boys, including association connections (UF, inferior fronto-occipital fasciculus, superior longitudinal fasciculus, external capsule, sagittal striatum, inferior longitudinal fasciculus), commissural connections (corpus callosum), projection connections (corona radiata, internal capsule), and brainstem connections (corticospinal tract, cerebral peduncles, medial lemniscus). Among youths with anxiety disorders, radial diffusivity analyses also revealed findings in boys and not girls in many of the regions in which the FA alterations were detected in boys (see Figure S4 in the
online supplement). Studies have demonstrated that DTI metrics can reflect different aspects of WM microstructure (
44). FA values broadly reflect microstructural integrity (e.g., myelination, cellular density, and/or axonal coherence) (
45,
46), whereas radial diffusivity, which is incorporated into FA, has been more consistently associated with changes in myelination (
47,
48). Taken together, the FA and radial diffusivity findings suggest that the WM changes observed in boys with anxiety disorders could be due to alterations in myelination. These broadly distributed WM findings in boys with anxiety disorders generally overlap with and connect components of the neural circuitry that underlie anxiety (
49,
50). For example, the UF is the primary WM tract linking prefrontal cortical and limbic regions, which are involved in the regulation and expression of anxiety, and the inferior fronto-occipital fasciculus facilitates prefrontal interactions with occipital and parietal regions thought to subserve functions relevant to emotion recognition and affective processing (
51,
52). Considering the essential role of myelination in supporting efficient neuronal communication and neural network synchrony, it is possible that the reductions in WM microstructural integrity in these pathways, observed in boys with anxiety disorders, contribute to aberrant communication between and among key neural networks implicated in the pathophysiology of anxiety disorders.
DTI studies of individuals with anxiety disorders have generally reported findings linking anxiety to reduced FA across various WM regions throughout the brain. The majority of this work has been performed in adult populations (
7–
9,
34,
53–
59). Importantly, very few studies have characterized WM alterations in youths with anxiety disorders (
4,
6,
18). A single study has been performed in adolescents (
6), and two in preadolescents (
4,
18), both conducted in our laboratory. Studies examining WM alterations in relation to anxiety disorder–related measures in normative populations also generally find negative associations between WM metrics and anxiety across preadolescents, adolescents, and adults (
33,
38,
60–
68). We note that a minority of studies also report positive WM-anxiety correlations, including two in participants with anxiety disorders and three in normative samples (
34,
35,
59,
69,
70).
Few studies in the WM-anxiety literature have examined the interactions between sex, anxiety, and WM microstructure. Our previous studies in preadolescents with anxiety disorders revealed a sexually dimorphic WM-anxiety relationship, such that boys with anxiety disorders, and not girls, demonstrated reduced UF FA (
4,
18). However, with regard to DTI studies of adolescents and adults with anxiety disorders, it is unclear whether any of these studies explicitly tested anxiety disorder-by-sex interactions in relation to WM metrics. Some studies in normative populations have tested the moderating effect of sex on WM-anxiety associations (
38,
62,
64,
66,
67,
69,
70). The two largest studies in adults reported significant negative correlations between corticolimbic pathway FA and trait anxiety only in females, and not males (
66,
67). These findings contrast with the male-specific anxiety disorder–related FA decreases in preadolescents detailed in the present study.
While the findings in this study demonstrate effects exclusively in boys and, given reductions in UF FA, provide a potential mechanism by which prefrontal-limbic interactions could be altered in relation to anxiety disorders, this does not preclude the possibility that girls with anxiety disorders also have prefrontal-limbic alterations. The sexually dimorphic nature of the finding here in preadolescent children is striking. While boys with anxiety disorders exhibit significant and widespread reductions in microstructural integrity relative to healthy volunteer boys, we find no evidence of anxiety disorder–related WM effects in girls in this cross-sectional analysis. We note that in a previous report from our laboratory (
18), we examined a large sample of girls longitudinally and found a within-participant negative association between anxiety symptom severity and whole-brain FA. The lack of a between-participant anxiety-WM effect in girls in the context of a significant longitudinal finding may suggest that anxiety disorder–related WM alterations in girls are more subtle than those in boys.
While the potential mechanisms underlying the sexually dimorphic findings in the present study are unclear, sex differences in WM maturation and oligodendrocyte biology have been reported. Some studies suggest that patterns of WM growth differ between males and females, although these are generally reported to be small effects (
71,
72). One study demonstrated a more protracted trajectory of WM growth in males and also a greater magnitude of FA in males (
71). Additionally, previous research has indicated sex-specific pubertal effects on WM microstructure, as well as differential relationships between sex hormones (i.e., testosterone, estradiol) and WM (
73–
75). Therefore, based on these findings, it would not be surprising to observe differences in WM parameters between boys and girls with anxiety disorders. However, in our sample, we did not find significant effects of age, pubertal status, or sex hormones on the observed group differences. It is still possible that a period of delayed WM growth could have occurred in boys with anxiety disorders prior to the age at which we studied them, and this could potentially contribute to the observed differences in FA magnitude between boys with anxiety disorders and healthy volunteer boys. A few preclinical studies have examined sex differences in oligodendrocyte and oligodendrocyte progenitor cell (OPC) biology (
76–
78). Findings include the demonstration that female OPCs exhibit more robust cell proliferation and migration capacities in response to injury, in addition to having higher levels of intracellular ATP (
76,
77). There is also some evidence to suggest that progesterone may protect female oligodendrocytes from cellular stress to a greater degree than for male oligodendrocytes (
78). In addition, male OPCs have been shown to have heightened sensitivity to cytotoxic stress and increased likelihood of cell death (
77). Because of the data supporting greater vulnerability of male OPCs, it is possible that elevated levels of early-life adversity/stress—which are known to increase the risk for psychopathology and negatively affect WM microstructure—may differentially impact male and female OPCs.
Some limitations must be considered in the interpretation of the study results. Although this study was performed in a relatively large sample of children, the majority of the study participants were non-Hispanic White children. Studying more individuals from other racial and ethnic groups in future studies is critical to more fully understand the relations between WM and anxiety across diverse populations. This study focused on the preadolescent period (8–12 years old), and interpretation of the results is therefore limited to this age range. Also, the subgroup of anxious boys was relatively small. While we assessed estradiol and testosterone in relation to WM-anxiety associations, it may be important to include other hormonal markers, including dehydroepiandrosterone, in future studies. In relation to interpreting the data from this study, it is important to emphasize that DTI metrics are indirect measures that reflect water diffusion patterns within brain tissue and therefore may reflect not only WM properties but also other factors that influence water diffusion (e.g., other glial cells, microtubules) (
79). Additionally, while DTI metrics can reflect myelin content, they can also reflect other microstructural properties, including fiber density and cellular permeability (
45). Another important consideration is that tensor-based metrics are limited in their ability to resolve complex microstructural features, such as crossing and bending fibers, as well as partial volume effects. Finally, other evidence suggests that girls may have more subtle anxiety disorder–related WM relations, which, in a cross-sectional design, might be elucidated with other methods, such as neurite orientation dispersion and density imaging (NODDI) and quantitative relaxometry.
In summary, in the largest cross-sectional DTI study of childhood anxiety to date, the results demonstrate that childhood anxiety disorders are associated with broadly distributed alterations in WM microarchitecture across the brain, and, importantly, this cross-sectional relationship is evident only in boys. Anxiety disorders most commonly emerge during preadolescence and early adolescence, periods during which there is rapid WM development (
1,
10,
80). While not addressed in this study, it is possible that the WM microstructural alterations observed here are related to the underlying pathophysiology of childhood anxiety disorders. Because evidence demonstrates that WM can dynamically change in relation to various environmental, behavioral, and pharmacological factors (
12–
14,
81), WM microarchitecture could be a potentially modifiable target for treating childhood anxiety disorders.