Recently, attention has been drawn to the possibility that interoceptive systems may be harnessed to treat major depressive disorder (
18). These systems relay information related to the physiologic state of the body to the central nervous system, and include sensory systems relevant to non-painful and painful senses. Non-painful interoceptive signals include those related to air hunger, bladder, blood serum (pH, osmolality, glucose), cardiovascular, fatigue, genital sensation, itch, gastrointestinal (esophageal, gastric, intestinal, colorectal), hunger, muscle tension, respiratory, sensual touch, shudder, temperature, thirst, tickle, and vasomotor flush physiologic processes (
18). One such interoceptive system is the thermosensitive system, with afferent fibers traveling within spinoparabrachial and spinothalamic pathways (
19). In recent open (
20) and randomized, double-blind, sham-controlled trials (
21), we have shown that infrared whole-body heating (WBH) has potential for the treatment of major depressive disorder. WBH is a novel intervention that appears to hold promise as an alternative and/or adjunct to pharmacological therapies based on data that it has a rapid and sustained antidepressant effect, confers none of the central nervous system (CNS)-mediated side effects that bedevil other somatic therapies for depression, and may operate via a body-to-brain pathway that provides biomarkers that reliably predict who is likely to respond (
19–
21).
Harnessing Evolved Sensory Signaling Pathways to Circumvent Limitations Inherent in Whole-Brain Manipulations
With few exceptions, currently extant somatic treatments for depression (e.g., antidepressant medications, electroconvulsive therapy) share in common mechanisms of action that simultaneously affect multiple areas of brain functioning, only a portion of which are likely to confer antidepressant benefit (
22). As noted earlier, it is increasingly recognized that the current pharmacological approach of indiscriminately targeting receptors and/or reuptake sites for neurotransmitter systems that impact widely distributed, and functionally divergent, brain regions may have reached the limit of its effectiveness (
5). One very obvious reason for our inability to materially enhance efficacy with this approach is the fact that it fails to recognize the highly complex functional topology of the CNS and the corollary truth that the same neurotransmitter and/or receptor can have very different, and not infrequently opposing, physiologic and behavioral effects, depending on the anatomical pathway in which they operate (
23). What is needed, therefore, is a methodology for enhanced anatomical (and, hence, functional) specificity in our interventions. In the present article, we explore the possibility of achieving this specificity by utilizing anatomical and functional specificity built into peripheral sensory signaling pathways by evolutionary processes to selectively affect functioning in specific brain regions linked in animal and human studies to depression and well-being (
22). For example, spinal afferents innervate multiple brain regions implicated in the etiology and pathophysiology of major depressive disorder (
24,
25).
Supporting this approach, prior studies by our group suggest that altering functional activity in such peripheral sensory pathways—including the innate immune inflammatory response—produces antidepressant effects without incurring the multiple disadvantages inherent in directly manipulating the CNS with either drugs or devices. For example, utilizing a randomized, placebo-controlled design in 60 patients with treatment-resistant depression, we have recently found that peripheral blockade of the proinflammatory cytokine tumor necrosis factor (TNF) with a biologic agent (infliximab) too large to cross the blood-brain barrier produces antidepressant effects that are similar in magnitude to those seen with standard antidepressants in individuals with elevated concentrations of peripheral inflammatory biomarkers (i.e., C-reactive protein [CRP] and TNF) while having no effect, or a detrimental effect compared with that of placebo, in patients with progressively lower levels of these same biomarkers (
26). Importantly, treatment with infliximab was associated with an incidence of side effects no greater than those of placebo, consistent with the notion that modulating peripheral pathways that signal the CNS via specific, evolutionarily conserved, mechanisms induces antidepressant effects that are apparent primarily in those individuals with evidence of abnormal functioning in these same pathways, while avoiding the myriad adverse events incurred by the widespread alteration of neurotransmitter systems typically induced by antidepressants. It is to this possibility that we now turn, focusing on the use of WBH to sensitize (enhance activity) in one such peripheral signaling system.
Antidepressant Effects of WBH in Humans and Antidepressant-Like Effects in an Animal Model
Based on both the theoretical considerations discussed earlier and basic/clinical studies conducted by our group (as discussed later), we have conducted two studies examining the acute antidepressant effects of a single session of WBH (
20,
21). In the first open trial, we found, in 16 medically healthy patients with major depressive disorder, that WBH induced a rapid, robust, and sustained reduction in depressive symptom scores (assessed with the German version of the Centers for Epidemiologic Studies Depression Scale [CES-D]) (
27), based on CES-D mean±SD scores dropping from 29.9±10.6 pretreatment to 19.2±12.3 five days posttreatment, t=4.53, df=15, p<.001, effect size=1.13. Thirteen of these subjects received no other pharmacological or psychotherapeutic intervention during this period, whereas three subjects were on chronic treatment with a selective serotonin reuptake inhibitor (SSRI) antidepressant (with no change in dosage during the study period). When looked at separately, WBH appeared to have no effect in these three subjects and, in fact, worsened depressive symptoms in one of them. In animal models, acute administration of SSRIs increases extracellular serotonin concentrations in the dorsal raphe nucleus, resulting in inhibition of serotonergic neuronal excitability and firing rates (
28). Meanwhile, studies in animals have shown that WBH activates brainstem serotonergic neurons, as measured by increased immediate-early gene expression (
29). If therapeutic effects of WBH depend in part on activation of brainstem serotonergic neurons, SSRIs may interfere with this effect through inhibition of serotonergic neuronal excitability. With the three subjects receiving SSRIs removed from analysis, the effect size of WBH increased, t=5.15, df=12, p<.001, effect size=1.4. Interestingly, the antidepressant response observed five days posttreatment was actually improved six weeks later in nine patients for whom these longer term follow-up data were available (CES-D mean score, 13.9). Although these patients received additional treatment during this period, these findings, although preliminary, suggest—at the least—that WBH may induce a rapid antidepressant response that can be solidified by longer term modalities. Although this was the first study, to our knowledge, to examine WBH specifically for major depressive disorder, our findings were consistent with reports that WBH improves mood in patients with cancer and improves quality of life scores in patients with type II diabetes (
30,
31).
We subsequently conducted a randomized, double-blind, sham-controlled study of the effects of WBH in 29 medically healthy individuals with major depressive disorder (
21). When compared with the sham group, the active WBH group showed improved Hamilton Depression Rating Scale scores across the six-week postintervention study period (WBH versus sham; week 1: −6.53, 95% CI=−9.90 to −3.16, p<.001; week 2: −6.35, 95% CI=−9.95 to −2.74, p=.001; week 4: −4.50, 95% CI=−8.17 to −0.84, p=.02; and week 6: −4.27, 95% CI=−7.94 to −0.61, p=.02). In a subsequent study, Naumann and colleagues (
32) found in a randomized, two-arm placebo-controlled study of medically healthy depressed individuals that an intervention consisting of two hyperthermic baths (40 °C) per week for four weeks improved depressive symptoms measured using the Hamilton Depression Rating Scale, relative to placebo, at the two-week time point. Although additional randomized, double-blind, placebo-controlled studies are needed, the studies discussed earlier suggest that infrared WBH or other thermal warming approaches hold promise as novel therapeutic approaches for the treatment of major depressive disorder.
Thermosensitive Systems and Major Depressive Disorder
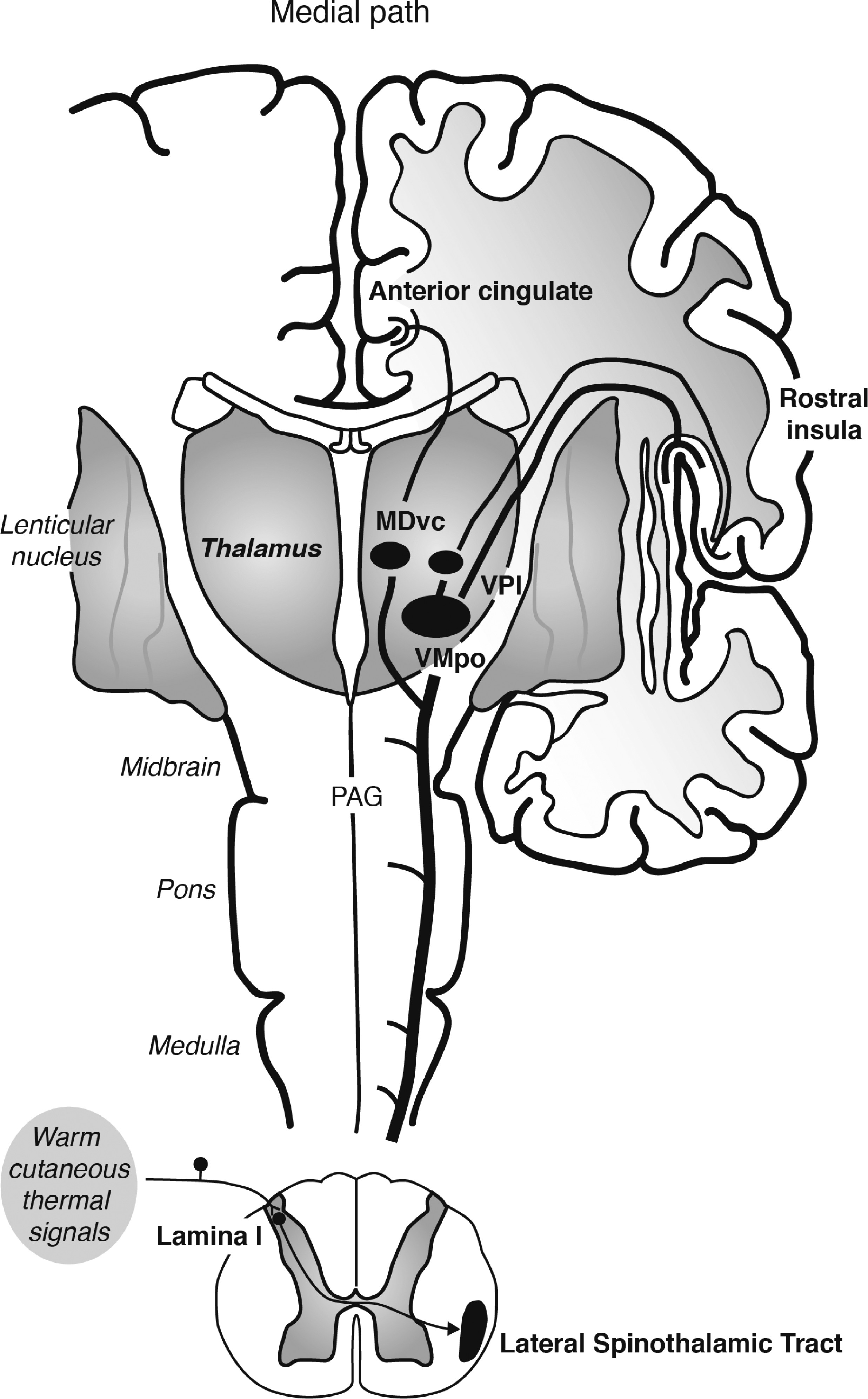
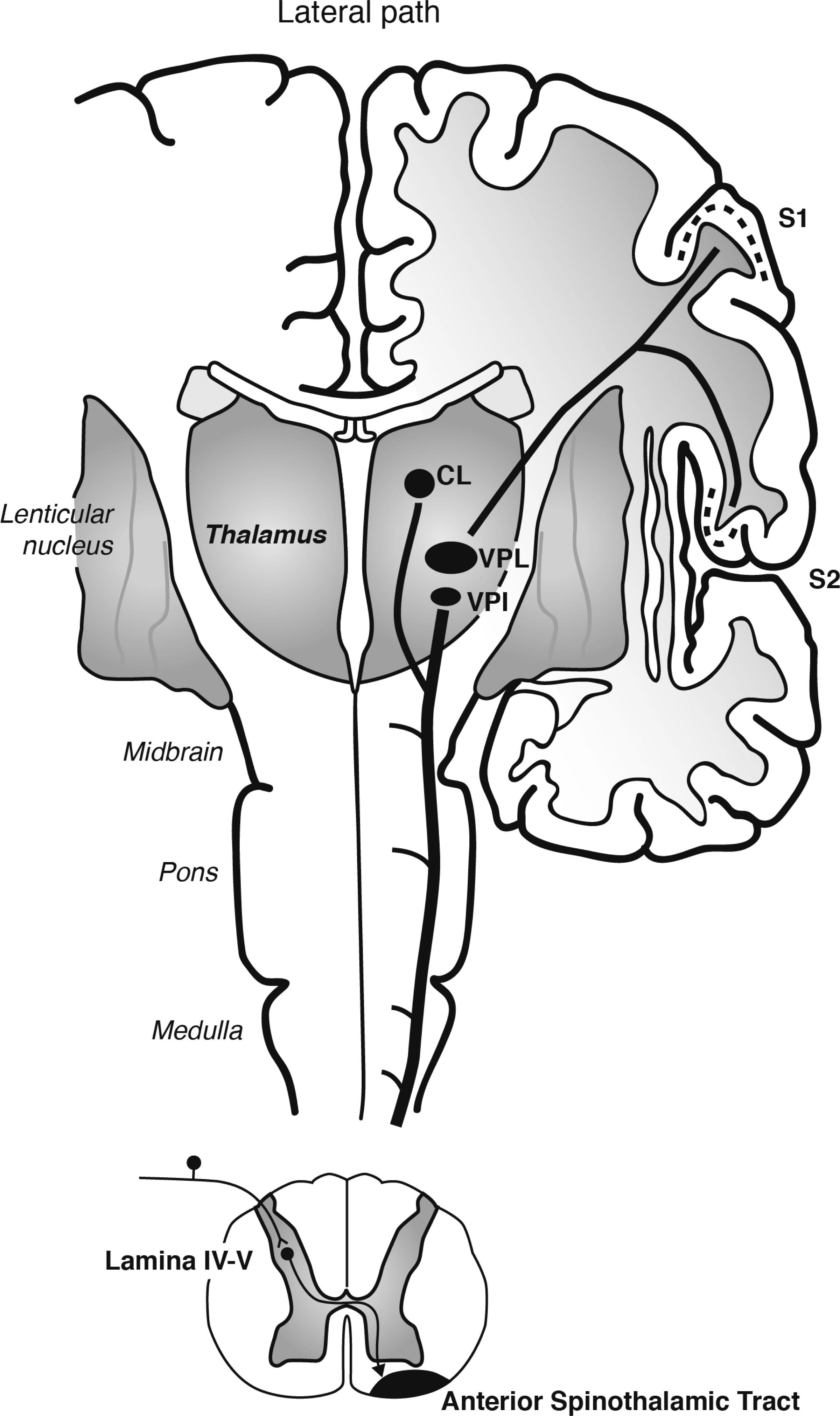
Warm cutaneous thermal signals activate thermosensitive afferents that travel in spinal nerves and synapse in lamina I of the spinal cord (
Figure 1). There, signals traveling in the lateral spinothalamic tract are relayed via the spinoparabrachial and spinothalamic pathways to the CNS. Spinoparabrachial pathways have been implicated in thermoregulatory cooling responses to warm temperature (
33) and may be involved in the activation of brainstem serotonergic neurons (
22). Spinothalamic pathways relay signals to somatosensory and limbic forebrain circuits (
19,
34–
36). Spinothalamic pathways originating in lamina I of the spinal cord include a lateral branch, which projects to the ventral posterior nucleus of the thalamus (including the thermosensitive ventromedial posterior nucleus [VMpo]); the ventral posterior nucleus of the thalamus in turn projects to the mid/posterior dorsal insula, which relays signals to the anterior insula (
34,
35). This system has been proposed to play a central role in interoception (
34–
36). In addition, spinothalamic pathways originating in lamina I of the spinal cord include a medial branch, which projects to the ventral caudal subdivision of the mediodorsal nucleus (MDvc) (
36). The MDvc in turn projects to the anterior cingulate cortex (
36). Together, the spinothalamic pathways relaying signals to the insula and anterior cingulate cortex are thought to represent the affective aspects of thermosensation (
34–
36). In addition, spinothalamic afferents originating in deeper layers of the spinal cord, lamina IV-V, travel in the anterior spinothalamic tract to also innervate the ventral posterior nucleus of the thalamus (in this case, the ventral posterolateral nucleus [VPL] and VPI); the ventral posterior nucleus of the thalamus in turn projects to the S1 and S2 regions of the somatosensory cortex (
Figure 2). The spinothalamic pathways relaying signals to the somatosensory cortex are thought to represent the discriminative aspect of thermosensation (
36).
To study effects of WBH on brain serotonergic neurons and antidepressant-like behavioral responses in rodent models, we placed rats in incubators with temperature maintained at 37°C or room temperature for 85–105 minutes (
29,
37) and monitored body temperature using rectal thermometry (
29) or chronically implanted biotelemetric devices (
37). Consistent with older animal studies suggesting that WBH affects mood-relevant serotonergic activity in the CNS (
38), we have shown in rodent models that WBH acts synergistically with a subthreshold dose of the SSRI citalopram to produce an antidepressant-like behavioral effect (
37). We have also shown that WBH activates serotonergic neurons in the midbrain (dorsal raphe nucleus, interfascicular part [DRI], and dorsal raphe nucleus, ventrolateral part [DRVL]) (
29) that are implicated in antidepressant (
22) and antipanic-like (
39) behavioral effects, respectively.
Consistent with these preclinical findings, studies using functional magnetic resonance imaging have shown that warm (41°C) cutaneous stimuli applied to the hand of humans induces activations in the midorbitofrontal cortex, pregenual cingulate cortex, and the ventral striatum that are correlated with subjective pleasantness ratings (suggesting a central role of spinothalamic afferents originating in lamina I of the spinal cord, innervating the MDvc and subsequently the anterior cingulate cortex [
Figure 1]) (
40). In contrast, intensity ratings of the stimuli were associated with activation of the somatosensory cortex and ventral and anterior insular cortices (suggesting a central role of spinothalamic afferents innervating the ventral posterior nucleus of the thalamus and subsequently the somatosensory and insular cortices [
Figure 1 and
Figure 2]) (
40). It is important to note that the brain areas associated with affective responses to warm cutaneous temperature constitute neural systems that are dysregulated in patients with depression (
25,
41–
43) and are projection targets for DRI serotonergic neurons (
22,
44,
45). Indeed, DRI neurons project heavily to the orbitofrontal cortex and anterior cingulate cortex in primates (
46) and to the medial prefrontal cortex and ventral striatum in rodents (
47). Moreover, DRI neurons project to other nodes within neural circuits implicated in depression, including frontal and entorhinal cortices, hippocampus, and midline thalamus (
25,
47).
The relevance of these findings and considerations for the pathophysiology and treatment of mood disorders is further highlighted by the fact that major depressive disorder has been repeatedly associated with physiologic changes that would be expected to result from suboptimal activity in thermoregulatory circuits (e.g., increased core body temperature as a result of suboptimal thermoregulatory cooling ability [
48–
54], including a decreased ability to sweat [
52–
55]). Moreover, effective treatments tend to reverse these abnormalities (
49). In this regard, it is important to emphasize that a single session of WBH resulted in long-term lower core body temperature (potentially via increased thermoregulatory cooling) in our open trial in humans (
20), raising the intriguing possibility that the procedure may actually normalize functioning of the thermosensory pathways and thus impact core depressive pathophysiology, as opposed to merely utilizing this pathway for therapeutic purposes. Furthermore, our finding that increased core body temperature prior to WBH predicted symptom improvement suggests that treatment response may identify a subgroup of patients with depression with some degree of biological homogeneity and that, therefore, biomarkers might be developed to predict who would and would not respond.
The primary conceptual shift in this review is to highlight the potential etiological and therapeutic relevance of peripheral sensory signaling pathways as “gateways” into the CNS functional circuitry implicated in the development of depression. The demonstration that WBH is superior to placebo in the acute treatment of depression, while conferring a very low side effect burden (
20,
21), opens the door to entirely novel technologies and treatments that would be easy to administer and highly cost effective. In addition to reducing the time to therapeutic response when added as an “accelerator” strategy to standard treatments, the observation of a prolonged antidepressant effect from a single WBH session raises the possibility that this modality might hold promise as a “stand-alone” option for the chronic treatment of depression. Moreover, if our preliminary observation that increased body temperature at baseline predicts therapeutic response is confirmed, WBH may enhance psychiatry’s ability to offer personalized treatment strategies, which is an important—and persistently elusive—goal. Finally, future studies should examine the potential utility of WBH as a preventive strategy for the development or relapse of depression (
56).