In the last decade a considerable spectrum of structural abnormalities has emerged from neuroimaging studies in affective disorders.
1 Important findings have come from volumetric investigations of subcortical structures. Previous lesion studies showing an association of depressive syndromes with basal ganglia alterations
2–7 were confirmed by MRI measurements showing smaller volumes of the caudate and the putaminal complex in patients with unipolar depressive disorders
8,9 and larger caudate nuclei in male bipolar patients.
10 These data, as well as functional imaging studies demonstrating hypoperfusion,
11,12 hypometabolism,
13–15 or higher dopamine D
2 receptor occupancy in basal ganglia regions,
16,17 indicated that basal ganglia may play a crucial role in the pathology of affective disorders. Along with recognizing the complex architecture of basal ganglia circuits,
18 it appears to be important to know which anatomic elements of the basal ganglia are predominantly affected in mood disorders.
Surprisingly, there is a lack of postmortem studies of brain volumes in affective disorders. Investigations of postmortem brains offer the possibility to determine volumes of brain structures to a high degree of resolution. Thereby, complex nuclei like the basal ganglia can be measured in detail. Since regional volumetric brain alterations are an indicator of macropathology in cerebral diseases, distinct clusters of structural deviations that might be detected by postmortem morphometry could contribute to our knowledge of the brain biology of mood disorders. In the present study, a planimetric method is used to measure volumes in postmortem brains from patients with mood disorders as compared with control subjects. This procedure addresses the following question: Is there a focal macropathology in subcortical structures, especially in the basal ganglia, in patients with mood disorders? We report here the first results of our postmortem morphometric study.
DISCUSSION
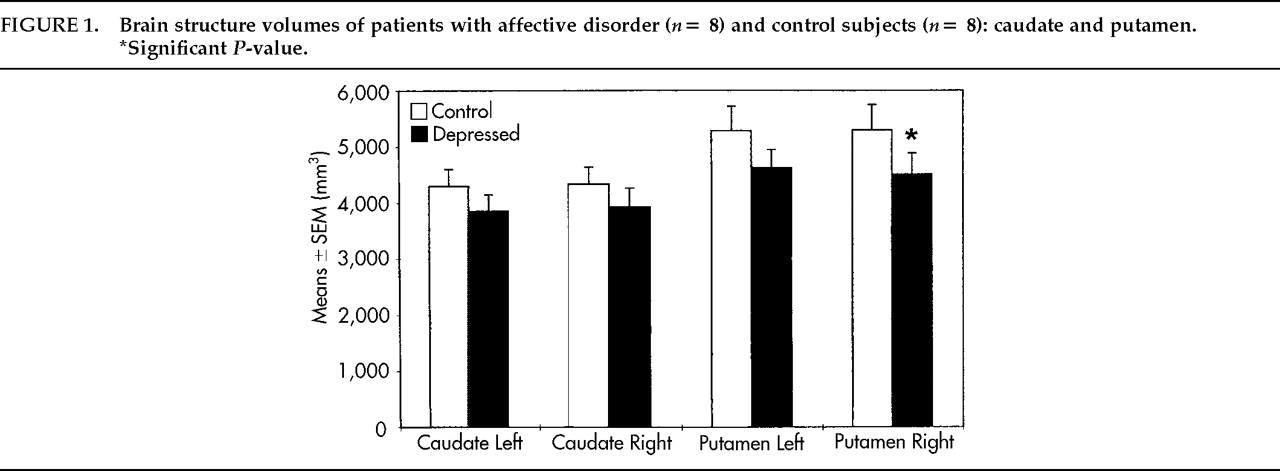
Results of the present postmortem study confirm and specify findings of basal ganglia abnormalities reported from CT or MRI studies. Neuroimaging studies have shown that structural abnormalities exist in basal ganglia of patients with mood disorders. In nonelderly patients with unipolar depression, controlled MRI studies found at least a trend for reduced volumes of the caudate and the putaminal complex.
8,9,21 However, this seems not to be the case for bipolar disorder.
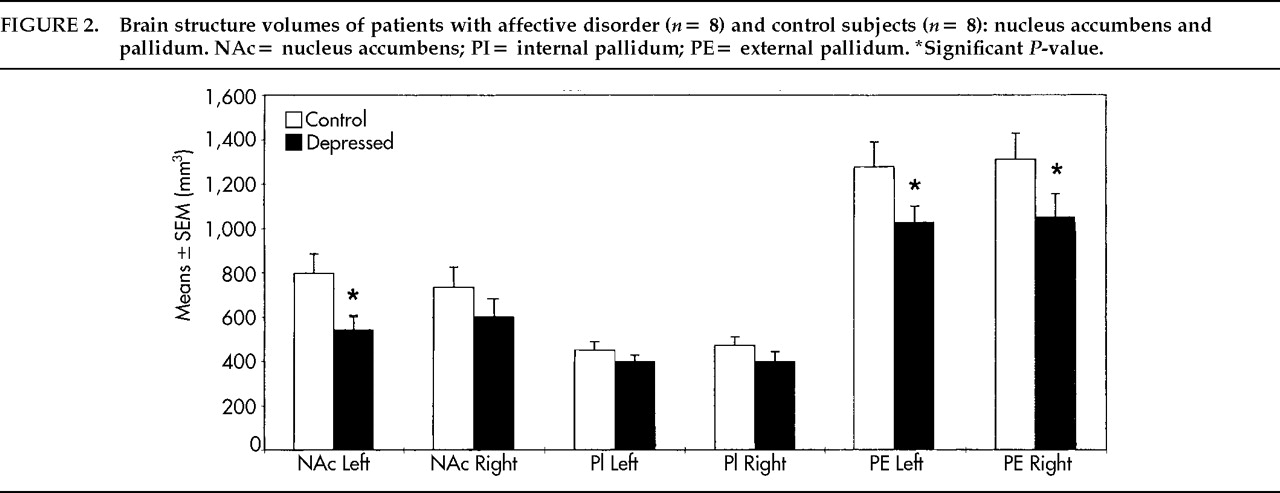
10,21–23In contrast to neuroimaging technology, postmortem morphometry provides the option of delineating and separating even small brain nuclei, such as the nucleus accumbens or the internal and external part of the pallidum, that have different connections and functions in basal ganglia circuits. These differences might explain in part the inconsistent results of MRI studies in which larger complexes of the basal ganglia were measured, such as the lentiform nucleus consisting of the pallidum and the putamen. More subtle structural abnormalities still cannot be detected by MRI. In the basal ganglia we found a focally accentuated volume reduction of the left nucleus accumbens, the external pallidum bilaterally, and the right putamen. Volumes of the left nucleus accumbens were significantly smaller in unipolar depressive patients and tended to be reduced in bipolar or schizoaffective patients.
This finding may have pathophysiological relevance, since the nucleus accumbens is part of the ventral striatum and thereby belongs to the limbic nuclei.
18,24,25 There is a reciprocal projection between the nucleus accumbens and the ventral pallidum,
26,27 which share common neuromodulators such as γ-aminobutyric acid and opioids.
28,29 Furthermore, the nucleus accumbens is the terminal of the mesolimbic dopaminergic projection from the ventral tegmental area.
30,31 These two pathways are known to be relevant to rewarding effects
32–34 and locomotion.
35–38 Moreover, there exist afferents to the nucleus accumbens from cortical and subcortical limbic structures such as the entorhinal cortex, prelimbic and infralimbic cortices, the anterior cingulate, the hippocampus, the mesencephalon including the dorsal raphe nucleus, the basal amygdala, and the lateral hypothalamus.
39–41 For this reason the nucleus accumbens is believed to be a pivotal structure at the interface of the limbic system and the basal ganglia and might be associated with psychic functions like mood and motivation.
42 The volume reduction of the left nucleus accumbens suggests pathology in the limbic loop of the basal ganglia at a site where information from neocortical inputs is modulated by limbic afferents before entering the nucleus accumbens. From the accumbens, information is transferred to the pallidum as one of the outputs of the basal ganglia to the thalamus, the cortex, and the brainstem.
18,43 The question remains open whether distinct compartments of the accumbens (that is, the core or the shell),
44 and thereby specific afferents to this nucleus, are altered in mood disorders.
Long-term application of neuroleptics reportedly may cause neuronal hypertrophy in the rat striatum
45 and volume increases of the striatum in humans.
46,47 These results are consistent with findings of a hypertrophic striatum in patients with Parkinson's disease, apparently a result of dopaminergic underactivity.
48 Thus, neuroleptic medication taken by 6 of 8 patients in our study might have increased volumes of the caudate and the putamen, which possibly were reduced in an unmedicated state.
Moreover, it is conceivable that only a distinct anatomic compartment of the striatum, such as the striosomes or the matrix, will show structural abnormality. This distinction might prove relevant, since connectivity with the neostriatum is topographically organized in these compartments; for example, most projections to the pallidum originate from the matrix,
49 and inputs from limbic system–affiliated structures are largely directed to striosomes.
50 Postmortem studies using acetylcholine-esterase staining might be useful to address the question of whether structural abnormalities in the anatomical and functional parcellation of the striatum exist in affective disorders.
Several neuroanatomical models of mood regulation have been proposed in recent years.
1,51–57 From these models it could be hypothesized that the striatum and the ventral pallidum might be key structures of the basal ganglia implicated in the pathology of depressive disorders. Results of our study confirm this assumption, particularly emphasizing the role of ventral parts—that is, the limbic parts of the striatum—in this pathology. The structural abnormality of limbic basal ganglia in patients with affective disorders is consistent with the notion that these subcortical structures might be involved not only in the control of movement, but also in the motivation for action
58 and psychomotor behavior.
59 Both lack of drive and psychomotor retardation represent foci of psychopathology in mood disorders.
A question that has to be addressed is whether volume alterations could be caused by medication. Patients included in this study received variable amounts of antidepressants, neuroleptics, sedatives, and lithium. Although antidepressants may be toxic in hepatocytes,
60 there is no evidence, at least from clinical data, that antidepressants cause shrinkage of brain tissue.
61 From animal work, antidepressants are shown to 1) counteract cell death caused by neurotoxins,
62 2) attenuate stress-induced morphological changes in the rat hippocampus,
63 and 3) elicit regenerative changes in the cerebral cortex after toxic lesions.
64 In our study, mean daily doses of tricyclic and tetracyclic antidepressants given in the last 4 weeks of life did not correlate with any of the structure volumes. Thus, it appears unlikely that antidepressant medication might be responsible for the volume alterations found in this study.
Furthermore, mean daily doses of neuroleptics did not correlate with any of the significantly changed volumes. As noted above, however, chronic application of neuroleptics can elicit volume increase of parts of the striatum. Thus, the finding of smaller volumes of the nucleus accumbens does not seem to be a result of neuroleptic medication. On the other hand, although neuroleptic medication in most cases in this study group was at modest levels, it could not be ruled out that hypertrophy induced by neuroleptics might have covered primarily existing volume deficits of the putamen and the caudate in patients. Besides differences in measurement strategies, the divergent amounts of neuroleptic medication might in part explain the discrepant results between our study and MRI studies that described smaller volumes of caudate and putamen in unipolar depressive patients or a greater caudate nucleus volume in bipolar disorder.
8–10In addition, comparisons of patients who received either sedatives or lithium with those who got no such medications yielded no differences with respect to volumes of the investigated structures. Moreover, illness duration did not correlate with any of the structure volumes. It is therefore unlikely that smaller volumes of the left accumbens, the right putamen, and the lateral pallidum bilaterally are a result of medication.
A crucial point in the evaluation of postmortem volumetry is to consider perimortal and postmortal processes that may influence brain structure. Protracted agonal states can induce acute alterations of brain tissue due to hypoxia. In our study, 2 patients (cases 6 and 7) and 2 control subjects (cases 5 and 7) suffered 3 or 4 days of agony. By neuropathological examination, none of these brains showed macroscopically visible edema. Acute nerve cell alterations or phagocytosis were not present in the measured structures. Moreover, possible alterations due to agonal state are comparable in patients and control subjects.
Postmortal processes such as postmortem delay and shrinkage of tissue due to embedding in paraffin were not different in the patients and the control subjects. Shrinkage factors were included in the volume calculations. Mode of tissue fixation did not influence our morphometric data, since alteration of brain weight by fixation with 8% formaldehyde is remarkable only in the first 4 weeks of fixation.
65–67 The structural abnormalities found in the patient group are not an effect of general reduced brain size, since brain weight of the investigated patients was even slightly higher than that of control subjects and brain weight was calculated as a covariate.
Although the volume reduction of the left nucleus accumbens was most prominent in unipolar depressed patients, it remains unclear whether our findings are specific to certain subtypes of mood disorders, since the sample of patients was too small. Moreover, the limited number of cases here reported emphasizes the pilot character of the study.
In sum, results of this preliminary report support the hypothesis that predominantly limbic parts of the basal ganglia—that is, the nucleus accumbens and the pallidum—are a focus in the pathomorphology of affective disorders. Because of limitations in neuroimaging technology, such subtle structural deficits of basal ganglia have not yet been shown in affective disorders. The histological basis underlying the described volume alterations, their etiology, and their relationship to depressive symptomatology remain to be clarified. Larger samples are needed in order to detect possible structural differences of basal ganglia between unipolar depression and bipolar or schizoaffective disorder.