One way to distinguish between the two views of hypnosis is to show that it selectively alters neural mechanisms that should underlie a hypnotically induced experience (compare with reference
7). According to this logic, people cannot voluntarily alter the workings of the neural mechanisms that underlie a specific experience—for example, those involved in perception—unless they have the experience. The initial studies that adopted this approach relied primarily on event-related potentials and did in fact find that hypnotic sensory alteration is associated with changes in those potentials
(8–
11). However, this technique does not allow one to localize activity in the brain precisely, and one could argue that it is possible that the effort of “adopting a role” affects event-related potentials. For example, Jasiukaitis et al.
(11) asked subjects to view obstructions on the left or right side of the visual field and showed that some components of event-related potentials to signals on the obstructed side were diminished and that these effects were different from simple inattention. However, one could argue that the subjects simply did not look as carefully on these sides, defocused their eyes, or even averted their gaze. Although the absence of latency differences makes this explanation unlikely, a paradigm that requires consistent attention to the stimulus and provides better brain localization of effects is necessary to clarify the relationship between hypnotic experience and brain function.
The more recent imaging techniques provide excellent brain localization, but comparatively little use of such techniques has been made in studying hypnosis
(12–
16), and the results have been inconsistent. There is some evidence that hypnotic analgesia, which involves somatosensory imagery, is associated with increased blood flow in the somatosensory cortex as well as the orbitofrontal cortex
(17). However, a more recent positron emission tomography (PET) study has indicated that hypnotic reduction of the perceived unpleasantness of a stimulus is associated with significant changes in pain-evoked activity within the anterior cingulate but not in the somatosensory cortex
(18). In addition, Szechtman et al.
(19) measured regional cerebral blood flow (rCBF) by means of PET while highly hypnotizable subjects were hypnotized and asked to produce vivid auditory hallucinations. Subjects who could produce the hallucinations (eight of the 14 tested) had increased rCBF in the right anterior cingulate gyrus, and the externality and clarity of the hallucinations were highly correlated with blood flow in this region. By contrast, those who could not hallucinate had activation in the auditory association cortex (Brodmann’s area 22) but not in the anterior cingulate. Moreover, during the actual hearing of sounds, the hallucinators showed greater activation in the temporal lobe auditory regions than did those who could not hallucinate, which may suggest that hallucinatory abilities can also be mobilized to amplify ordinary perception. However, because all of the subjects in this important study were formally hypnotized, it is more informative about the neurophysiology of hallucination than about the effects of being hypnotized per se.
Method
Eight subjects (six women and two men) participated as paid volunteers recruited through advertisements posted at Harvard University. The research was approved by the Harvard University Faculty of Arts and Sciences Committee on the Use of Human Subjects and by the Massachusetts General Hospital Human Research Committee. Subjects had a mean age of 24 years (range=20–35). Scores on the Hypnotic Induction Profile
(4) and the Stanford Hypnotic Susceptibility Scale: Form C
(26) were obtained from a total of 125 subjects. Possible scores on the Hypnotic Induction Profile ranged from 0 to 10, and scores on the Standard Hypnotic Susceptibility Scale ranged from 0 to 12. The subjects included in the study scored 9–10 on the Hypnotic Induction Profile (mean score=9.1) and 10–11 on the Standard Hypnotic Susceptibility Scale (mean score=10.5). Subjects were not only prescreened with the hypnotizability scales but also evaluated and selected on the basis of their performance of the task. Subjects were shown both the color and gray-scale stimuli during the screening session, and after hypnotic induction, they were asked to drain the color from the color stimulus, add color to the gray-scale one, and report whether they could do so. All subjects included in the study reported being able to alter the stimuli during the screening session. Eight other subjects selected for being extremely low in hypnotizability were also scanned but were unable to perform the visual hallucination task, and their blood flow results were inconsistent. After complete description of the study to the subjects, written informed consent was obtained, and subjects were positioned in the PET scanner.
PET Procedures
Briefly, subjects were aligned relative to the canthomeatal line and fitted with a thermoplastic face mask, a set of nasal cannulae, and a vacuum mask. Transmission measurements were taken with an orbiting rod source before scanning. Subjects were imaged for a total of 1 minute per condition by means of [
15O]CO
2. Approximately 10 minutes separated each condition. The PET machine was a General Electric Scanditronix PC4096, 15-slice, whole-body tomograph (Uppsala, Sweden). The PET scanning procedures used at Massachusetts General Hospital have been described in detail previously
(27,
28).
Task Procedures
Each subject performed four tasks after having undergone hypnotic induction and four tasks with no hypnotic induction. The order of hypnotic states was balanced so that one-half the subjects began with the hypnosis block and one-half began with the no-hypnosis block. Within each block (hypnosis and no hypnosis), subjects were presented with a rectangular color stimulus that itself was made up of a set of smaller rectangles, thus making the stimulus resemble a painting by the Dutch artist Piet Mondrian. The stimuli appeared on a 13-inch color monitor placed at a distance of approximately 50 cm from the subject’s eyes. As illustrated in
Figure 1, there was also a gray-scale version of the stimulus, which was produced to have comparable saturation and intensity differences as the color one but with no variations in hue.
All subjects received the following four conditions twice, once after the hypnotic induction procedure and once without hypnosis: 1) subjects were shown the color stimulus and asked to perceive it veridically (i.e., in color); 2) subjects were shown the color stimulus but asked to drain the color from it, visualizing the stimulus as if it were in shades of gray; 3) subjects were shown the gray-scale stimulus and asked to perceive it as it was; and 4) subjects were shown the gray-scale stimulus but asked to add color, visualizing it as if it were a color stimulus. Subjects were always asked to experience the stimulus in a certain way (i.e., veridically or altered) for the entire duration of the scan. The order of the blocks and conditions within a block was counterbalanced, except that the color stimulus was always presented first because we wanted subjects to know what the stimulus looked like when variations in hues were present.
In the hypnosis condition before scanning began, the subjects were asked to alter actively the stimulus, to drain or add color while focusing on the altered stimuli. They were asked to alter their perception of the stimuli as much as possible and to let the investigators know when they had successfully added or drained the color. The no-hypnosis condition was identical to the hypnosis condition except that when subjects were asked to perceive the gray-scale stimulus in color or vice versa, they were asked to try to “remember and visualize” the stimulus in its other form. Thus, the no-hypnosis condition served as a mental imagery control for the hypnosis condition. The wording used here was chosen to lead subjects to attend to the visible stimulus and to alter it, rather than to substitute a complete hallucination for the observed pattern. The instructions for this condition were worded to avoid leading the subjects, who were all highly hypnotizable, to fall into a hypnotic state during performance of the task.
PET Statistical Analysis
We analyzed the resulting data with the statistical parametric mapping technique
(29–
31) by means of the SPM 95 software package (Wellcome Department of Cognitive Neurology, London). At each voxel, the data acquired during PET scanning were normalized to a global mean of 50 ml/min/100 g and fit by the method of least squares to a linear model. Cognitive state (i.e., scan condition) was considered the main effect in the resulting analysis of variance (ANOVA) comparisons, and subjects were considered a block effect. Planned contrasts at each voxel were carried out. This technique fits a linear statistical model, voxel by voxel, to the data; hypotheses are tested as contrasts in which differences between conditions are evaluated by means of the t statistic. Data from all conditions were used to compute the contrast error term. Any z scores greater than 3.29 (p<0.001, uncorrected, two-tailed) were considered to meet statistical significance, except in the case of the lingual or fusiform gyri, where a strong hypothesis based on earlier results led us to expect greater activation during the viewing of color. In this case, we set the threshold at 3.09 (p<0.001, uncorrected, one-tailed). We considered these thresholds to be a compromise between the higher thresholds suggested by Gaussian fields theory, which supposes no previous hypotheses regarding the anatomic localization of activations, and simple statistical theories that fail to take into account the spatial correlations necessarily encountered with PET and other neuroimaging methods
(32).
The region-of-interest analyses were performed by means of software developed at Massachusetts General Hospital. The software permitted the tracing of a region of specified size and shape onto the statistical image outputs of SPM 95. Thus, values averaged over the voxels in the traced regions were extracted and later submitted to ANOVA comparisons between the conditions by means of the SuperANOVA program (Abacus Concepts, Berkeley, Calif.) for the Macintosh.
Results
We began by examining the results when subjects viewed color versus gray-scale stimuli and were told to see them veridically.
Table 1 notes all areas activated with z>3.29 or 3.09 (for strongly predicted regions), and
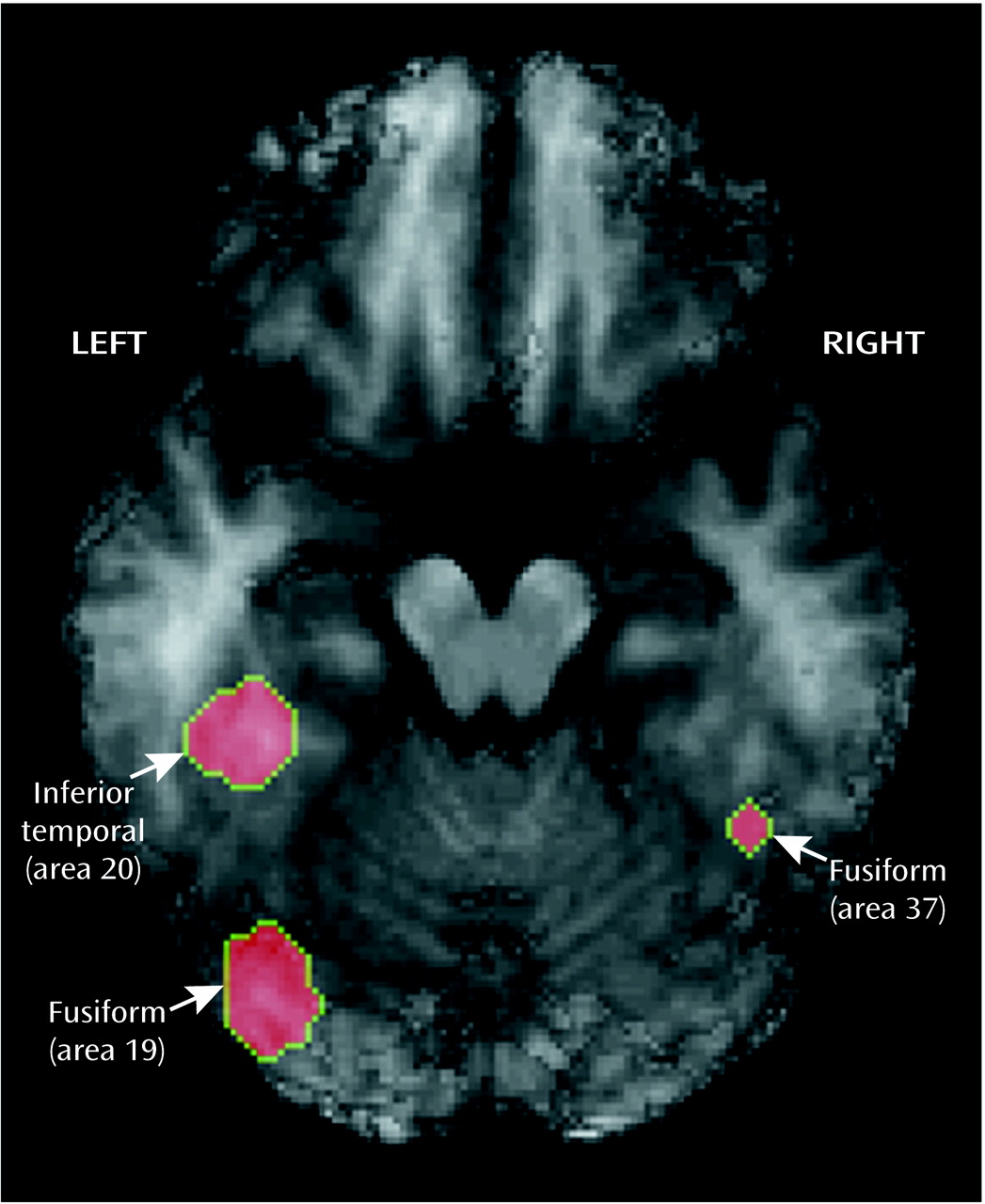
Figure 2 illustrates activation found in the left and right fusiform areas. As is evident, color is processed by a circuit, not a single area. However, it is notable that we found activation in the lingual or fusiform area, as reported by Lueck et al.
(23), Corbetta et al.
(24), Chao and Martin
(25), and Allison et al.
(33). We defined a region of interest with a 10-mm radius in the left hemisphere fusiform area at coordinates –34, –72, and –12 and another in the right hemisphere at coordinates 38, –50, and –12, both of which were activated during color perception. The locations of these two regions are clearly consistent with those from previous reports.
The results are illustrated in
Figure 3 and
Figure 4. We began by analyzing data from both regions of interest in an ANOVA. This analysis revealed that both the presentation of color (F=8.69, df=1, 7, p=0.02) and the perception of color independent of presentation (F=8.98, df=1, 7, p=0.02) resulted in greater blood flow than the presentation or perception of shades of gray. The hypnotic illusion of color induced blood flow change consistent with actually observing color. In addition, we found greater blood flow in the right hemisphere color region than in the left (F=5.30, df=1, 7, p=0.05). Moreover, the effect of what was presented was greater in the left hemisphere than in the right hemisphere (F=8.22, df=1, 7, p=0.02), and the effect of hypnosis on what was perceived was different in the two hemispheres (F=9.43, df=1, 7, p<0.02) for the interaction of state, color perception, and hemisphere. No other main effects or interactions were significant, including the state of being hypnotized.
Given the interactions with hemisphere, we next analyzed the two regions of interest separately.
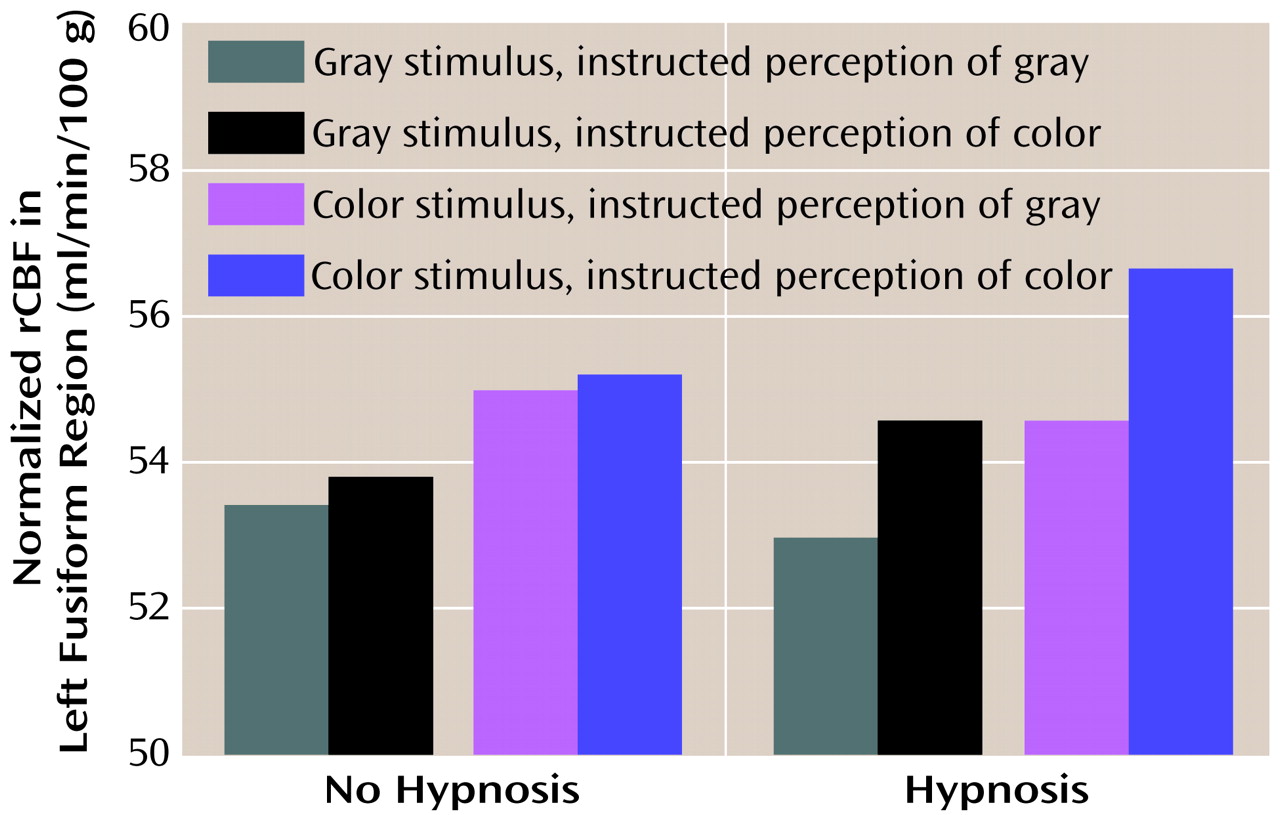
Figure 3 illustrates the results from the left region of interest. As in the initial analysis of the perception conditions, we found greater blood flow when color was displayed than when gray scale was displayed (F=11.96, df=1, 7, p=0.01), with means of 55.4 ml/min/100 g (SD=4.7) and 53.7 ml/min/100 g (SD=3.9), respectively. However, we also found greater blood flow when color was perceived than when gray scale was perceived, independent of what was actually presented (F=8.68, df=1, 7, p<0.03), with means of 55.1 ml/min/100 g (SD=4.6) and 54.0 ml/min/100 g (SD=4.0), respectively. In addition, we found that blood flow in the left region of interest varied with instruction to perceive color versus gray scale only when the subjects were formally hypnotized (F=21.48, df=1, 7, p<0.003, for the interaction of hypnotic state and stimulus perceived). Indeed, although the difference between the effects of instruction to perceive color versus gray scale was robust during hypnosis (t=7.8, df=7, p<0.0001), there was no effect when subjects were not hypnotized (t=1.3, df=7, p>0.25).
We next performed least squares means comparisons (Bonferroni-corrected to an alpha level of 0.008) on the interaction between hypnotic state and stimulus perceived. These tests revealed significant differences for three pairs of comparisons: “hypnosis, color perceived” versus “hypnosis, gray scale perceived”; “hypnosis, color perceived” versus “no hypnosis, gray scale perceived”; and “hypnosis, color perceived” versus “no hypnosis, color perceived” (t=7.8, df=7, p=0.0001; t=6.2, df=7, p=0.0004; and t=5.0, df=7, p=0.002, respectively). Thus, the “hypnosis, color perceived” condition resulted in significantly greater blood flow to the region of interest in all comparisons where it was examined. Not significant after correction were the “hypnosis, gray scale perceived” versus “no hypnosis, color perceived” (t=2.8, df=7, p<0.03), the “no hypnosis, color perceived” versus “no hypnosis, gray scale perceived” (t=1.2, df=7, p>0.25), and the “hypnosis, gray scale perceived” versus the “no hypnosis, gray scale perceived” (t=1.6, df=7, p>0.15) comparisons.
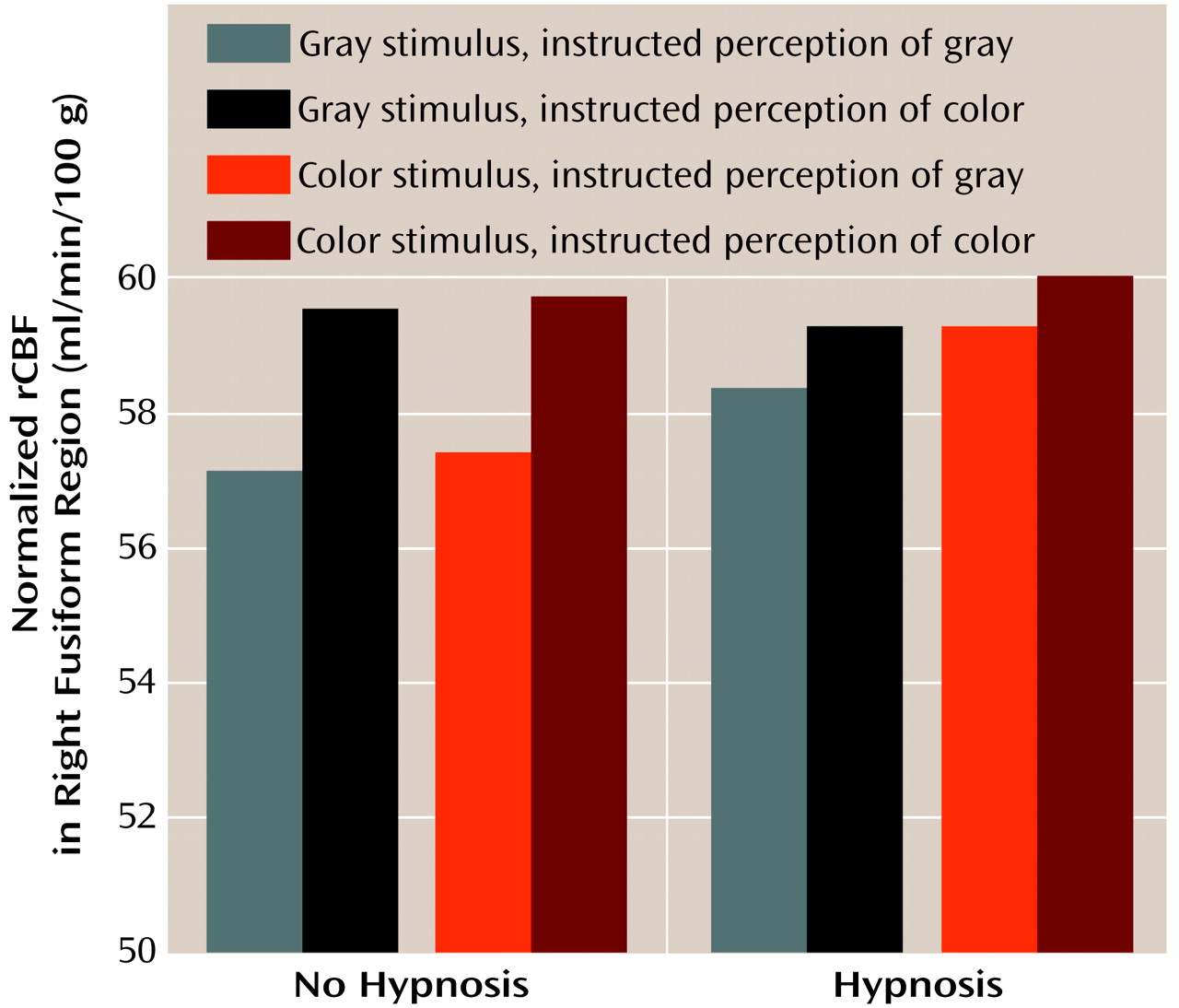
As indicated by the interactions with hemisphere in the main analysis, we found some different results in the right hemisphere color processing region. As illustrated in
Figure 4, the only factor that affected blood flow in the right region of interest was what subjects were asked to perceive (F=7.76, df=1, 7, p<0.03), with means of 58.1 ml/min/100 g (SD=3.5) for gray scale and 59.7 ml/min/100 g (SD=2.5) for color. No other main effect or interaction, including those with the state of being hypnotized, approached significance (p>0.13). The instruction to perceive the stimulus as color or gray scale overrode the effect of the actual stimulus presented, with means of 58.7 ml/min/100 g (SD=2.9) and 59.1 ml/min/100 g (SD=2.9) for gray scale and color stimuli, respectively (F=3.6, df=1, 7, p>0.20).
The means in the interaction between displayed and perceived colors revealed that in all cases asking the subjects to perceive color produced more activation than asking them to perceive gray scale. Least squares means contrasts (Bonferroni-corrected to an alpha level of 0.008) for multiple comparisons performed on all possible pairs revealed that these differences were all significant, with the exception of the “gray scale displayed, color perceived” versus “color displayed, gray scale perceived” comparison, which was just below the significance threshold after correction for multiple comparisons (t=3.3, df=7, p=0.01). For all other comparisons between conditions where subjects were asked to perceive color or gray scale, t values ranged from 4.34 to 5.98 and p values from 0.003 to 0.0006, df=7. Moreover, the amount of activation was the same when subjects perceived color that was presented versus when they visualized it on seeing gray scale (t=1.0, df=7, p>0.33) and when they perceived gray scale that was presented versus when they visualized it on seeing color (t=1.6, df=7, p>0.14).
Discussion
The results of this study revealed that people who are highly hypnotizable can modulate the neural responses underlying color perception by both enhancing these responses and diminishing them. The left hemisphere color area registered what they were told to see only when subjects were formally hypnotized. In contract, the right hemisphere region of interest registered what the subjects were told to see (independently of what was actually presented), whether or not they were formally hypnotized. The right hemisphere appeared to respond to imagery per se, whereas the left required the additional boost provided by hypnosis.
Our findings are consistent with those of Howard et al.
(34), who reported that ordinary mental imagery of color activated only the right fusiform area, not the left. Thus, our findings in the left hemisphere could not have been produced by mental imagery alone. If we had found only results in the right color area, we would simply have extended the findings of Howard et al.
(34), showing not only that imagery can activate these areas but also that it can literally override perceptual input. However, our results are stronger than that. We also found that hypnosis can activate the left color area in a way that imagery alone does not. This finding is consistent with results from a recent study by Maquet et al.
(15), who used PET to examine the functional aspects of the hypnotic state. They reported a predominance of left-side activation during hypnosis, as well as many areas that overlap with those found in studies of visual mental imagery
(11; see also reference
35). They concluded that multimodal mental imagery is an integral part of the hypnotic experience. Rainville et al.
(16) reached similar conclusions about the role of imagery and proposed that the predominance of left hemisphere activations may be due to verbal mediation of hypnotic suggestions, working memory functions, and top-down processes that may reinterpret the sensory experience.
The results in the right hemisphere are not as easily interpreted as those in the left and must therefore be viewed with caution. It is possible that the right-side activation across hypnotic and nonhypnotic conditions reflects not only the effects of imagery per se but also the trait of hypnotizability; however, our data do not allow us to address this issue directly, and so the resolution of this important question of distinctions between state and trait must be left to future investigations.
We conclude that hypnosis is not simply role enactment. Consistent with other recent work
(19), we find that the vivid changes in subjective experience attained under these hypnotic conditions are associated with changes in brain function that are typical of differences in actual perception among highly hypnotizable subjects.