Appreciation of the likely dimensional nature of autistic symptoms
(1 –
3) has led to the development of measures to evaluate autism-related social traits in the general population
(2 –
4) . The Social Responsiveness Scale (SRS) quantifies social, communicative, and other characteristic impairments of autism spectrum disorders. In the present article, the term autism refers to autistic disorder, Asperger’s syndrome, and pervasive developmental disorders not otherwise specified
(3) . The SRS, which has been extensively validated in both child and adult populations
(3,
5,
6), provides a single measure of autistic traits that, although heavily weighted on social-communicative impairment, represent a “singular, continuously distributed underlying factor” in the population
(7) . As such, the SRS may allow investigators to study the neurobiology of autistic traits in both clinical and nonclinical samples.
In the present study, we developed a protocol for examining the neural correlates of autistic traits by measuring the relation between informant-provided SRS scores and resting state functional connectivity measures in neurotypical individuals. Resting state functional magnetic resonance imaging (fMRI) has emerged as a powerful means of mapping and characterizing functional connectivity without the constraints of task-based approaches
(8 –
10) . Imaging and electrophysiological studies
(11,
12) examining intrinsic brain activity at rest in individuals with autism support a model of compromised long-range cortico-cortical connectivity
(13) . Recently, two resting state fMRI studies
(14,
15) that compared high functioning adults with autism with neurotypical comparison subjects revealed reduced intrinsic connectivity of long-range circuits based in pregenual portions of the anterior cingulate cortex.
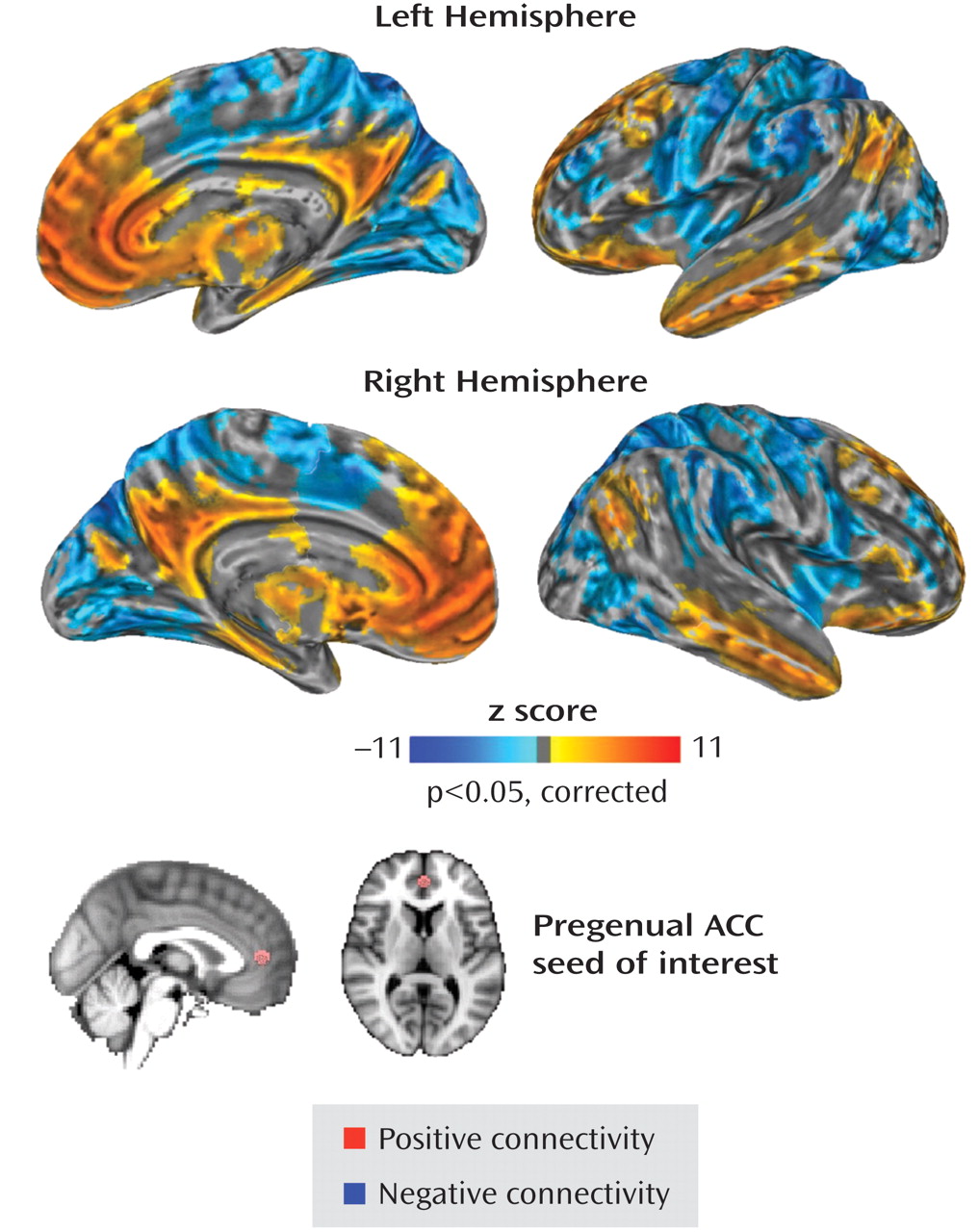
Involvement of the pregenual anterior cingulate cortex in autism is consistent with this region’s role in an individual’s capacity to reason about the thoughts and beliefs of others—also referred to as “theory of mind”
(16,
17) —which is qualitatively impaired in individuals with autism. Furthermore, hypofunction of this region in autism was revealed by a recent voxel-wise meta-analysis of functional imaging studies of social processing
(18) . This accumulating evidence suggests that disruption of the pregenual anterior cingulate cortex-based network may underlie autistic traits. Other likely nodes within this network are the posterior cingulate cortex, which is associated with self-referential processing and mentalizing
(19,
20), and the anterior insula, which is linked to representing and/or monitoring the salience of one’s own emotions and the emotions of others
(21 –
23) . Hypoactivation of both these regions has also been found in individuals with autism relative to neurotypical comparison subjects
(18) .
In the present study, we hypothesized that interindividual differences in autistic traits would be related to differences in the functional connectivity of the pregenual anterior cingulate cortex, even in a nonclinical population. Specifically, we predicted that higher SRS scores, indexing greater autistic traits, would be inversely related to the strength of functional connectivity of the pregenual anterior cingulate cortex with the 1) posterior cingulate cortex and 2) anterior insula.
Discussion
The present study represents an initial application of a protocol developed to examine the neural correlates of autistic traits, as indexed by the SRS-A, in neurotypical individuals using resting state fMRI. As hypothesized, we found that individual differences in autistic traits in a nonclinical population were related to pregenual anterior cingulate cortex functional connectivity. Several studies have linked individual behavioral performance to patterns of functional connectivity observed during rest (e.g.,
35 ). The present study extends this finding by relating an informant-based measure of social skills related to autistic traits
(3,
5 –
7) directly to pregenual anterior cingulate cortex connectivity.
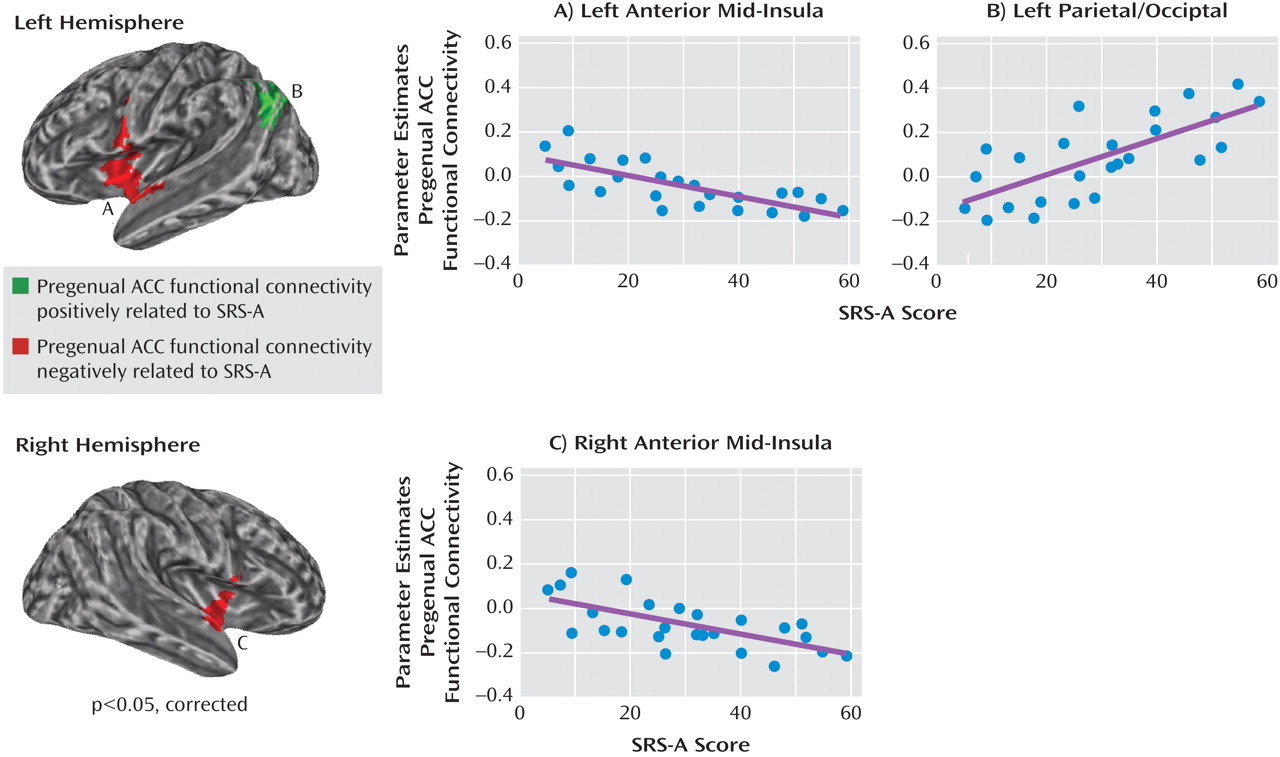
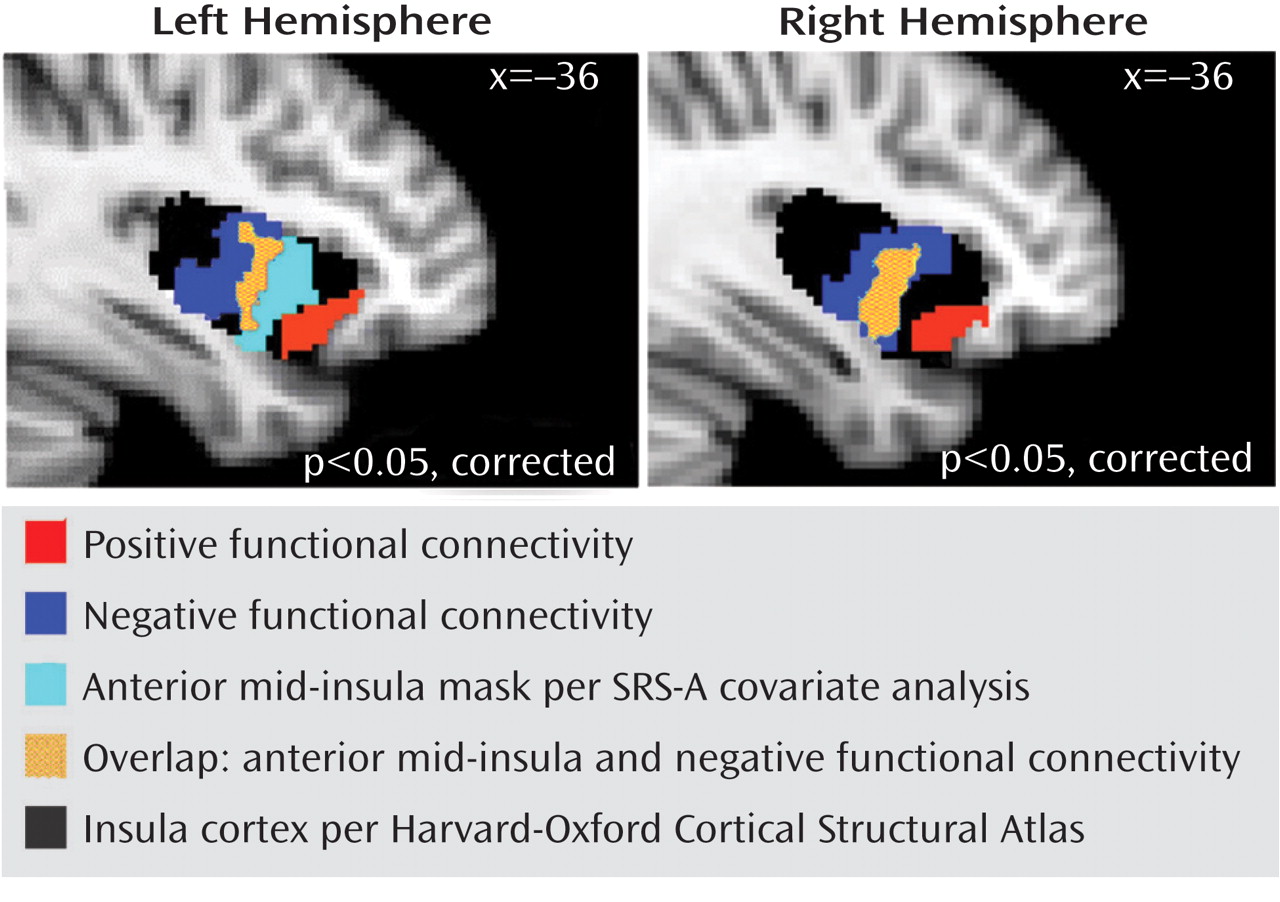
Consistent with one of our two predictions, autistic traits were related to pregenual anterior cingulate cortex connectivity with the insula, although this relationship was specifically limited to the anterior mid-insula rather than anterior insula per se. Discussing the implications of this finding requires considering the pattern of pregenual anterior cingulate cortex connectivity with various insula subregions. The insula can be subdivided broadly into the anterior, middle, and posterior, with the anterior insula containing the ventral and dorsal regions
(36) . The posterior insula, commonly involved in primary representations of visceral and somatic sensation
(33), showed no connectivity with the pregenual anterior cingulate cortex, which is consistent with a recent report
(23) . In contrast, the anterior and mid-insula subregions, implicated in maintaining higher order representations of sensation and emotion
(21,
33,
37), exhibited a significant pattern of pregenual anterior cingulate cortex functional connectivity along an anterior/posterior gradient, extending from positive to negative connectivity. At the anterior-most extent of the insula, the ventral anterior insula was positively connected with the pregenual anterior cingulate cortex along with a broader network of structures implicated in social cognition, such as the retrosplenial complex. By contrast, at the posterior extent of this gradient, the posterior mid-insula was negatively connected with the cingulate region of interest along with a network of structures implicated in somatic and self-focused attention (primary somatosensory cortices, superior parietal cortices, and primary motor cortices). Between these, an intervening anterior mid-insula region exhibited variable patterns of functional connectivity across subjects. This pattern of differential pregenual anterior cingulate cortex connectivity in anterior and middle insula subregions is consistent with the notion that the ventral anterior and posterior insula underlie social cognition and somatic attention, respectively
(21,
33,
37) .
These findings of differentiable mid-insula subregions are noteworthy when considered in the context of current models of insula organization, which tend to treat the mid-insula as a single subregion
(38) . Specifically, we found that the anterior mid-insula appears to function as a transition zone between the ventral anterior insula, which is related to social cognition
(21), and the posterior mid-insula, which is likely involved in somatic attention
(33,
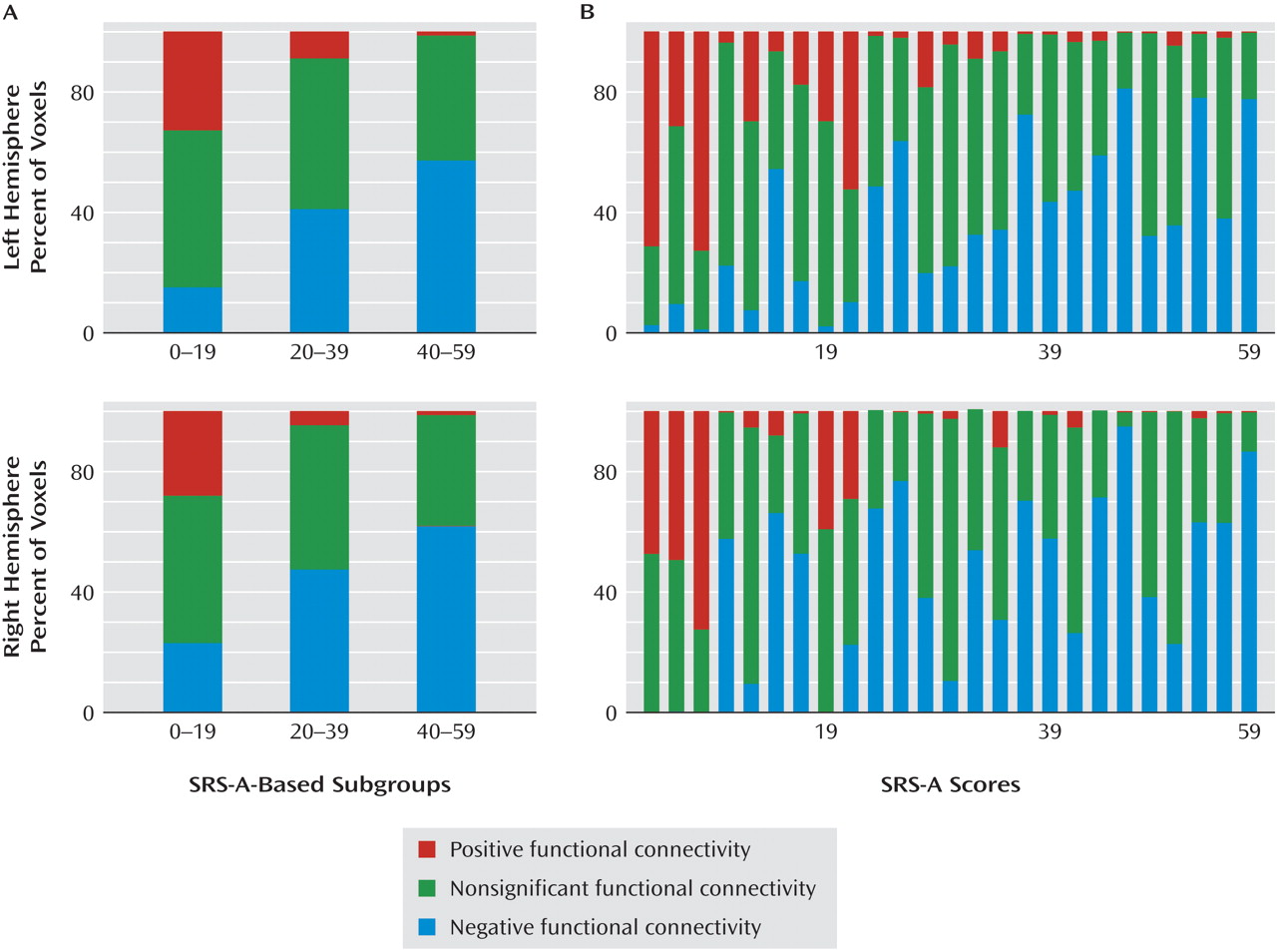
37) . As indicated by secondary analyses, in the subgroup with low levels of autistic traits, a substantial portion of the anterior mid-insula showed a positive connectivity pattern similar to that observed in the ventral anterior-insula. However, in the subgroup with elevated levels of autistic traits, the valence of functional connectivity in the anterior mid-insula resembled that of the posterior mid-insula (i.e., negative correlation with the pregenual anterior cingulate cortex). These results have broad implications for future efforts to relate interindividual differences in behavior to functional connectivity. Specifically, they highlight the need to interrogate connections exhibiting a high degree of variability across subjects as opposed to limiting analyses to a priori connections of interest or to only those analyses showing consistent patterns of connectivity across subjects.
We did not detect a significant relationship between SRS-A scores and pregenual anterior cingulate cortex functional connectivity with the posterior cingulate cortex, a region implicated in the development of theory of mind
(20) . A meta-analysis
(18) of functional imaging studies of autism found the posterior cingulate cortex to be hypoactive in autism during social tasks. Further, initial studies of functional connectivity in autism
(14,
15) reported decreased connectivity between the posterior cingulate cortex and pregenual anterior cingulate cortex. Beyond potential power concerns related to our sample size, we used a nonclinical screened sample that may not recapitulate brain-behavior relationships encountered in clinically diagnosed individuals with higher SRS-A scores. Future studies with affected individuals will test whether greater severity of autistic traits is related to functional connectivity between anterior and posterior cingulate regions implicated in social cognition
(18) .
We also found that increased SRS-A ratings were related to increased functional connectivity between the pregenual anterior cingulate cortex and higher order sensory processing regions (superior parietal gyrus, lateral occipital cortex, and angular gyrus). These regions are often abnormally hyperactive in individuals with autism
(18) . As such, although not hypothesized, our findings suggest the need to consider abnormal patterns of connectivity beyond hypoconnectivity.
In considering the imaging approach used in the present study, we note that although SRS-A scores may also be related to evoked brain activation using task-based approaches, examining the relationship between SRS-A scores and measures of resting state functional connectivity has several advantages. First, practice effects, floor/ceiling effects, and the need for compliance with task demands are minimized
(9) . This latter point is crucial for studies with behaviorally and intellectually challenged populations. Second, resting state analyses can be used to simultaneously delineate entire multiple networks, which are usually only partially observable in task-based studies, depending on the selected contrasts
(8) . Third, despite concerns about the unconstrained nature of the resting state, patterns of intrinsic connectivity during rest are remarkably replicable across individuals and across research labs
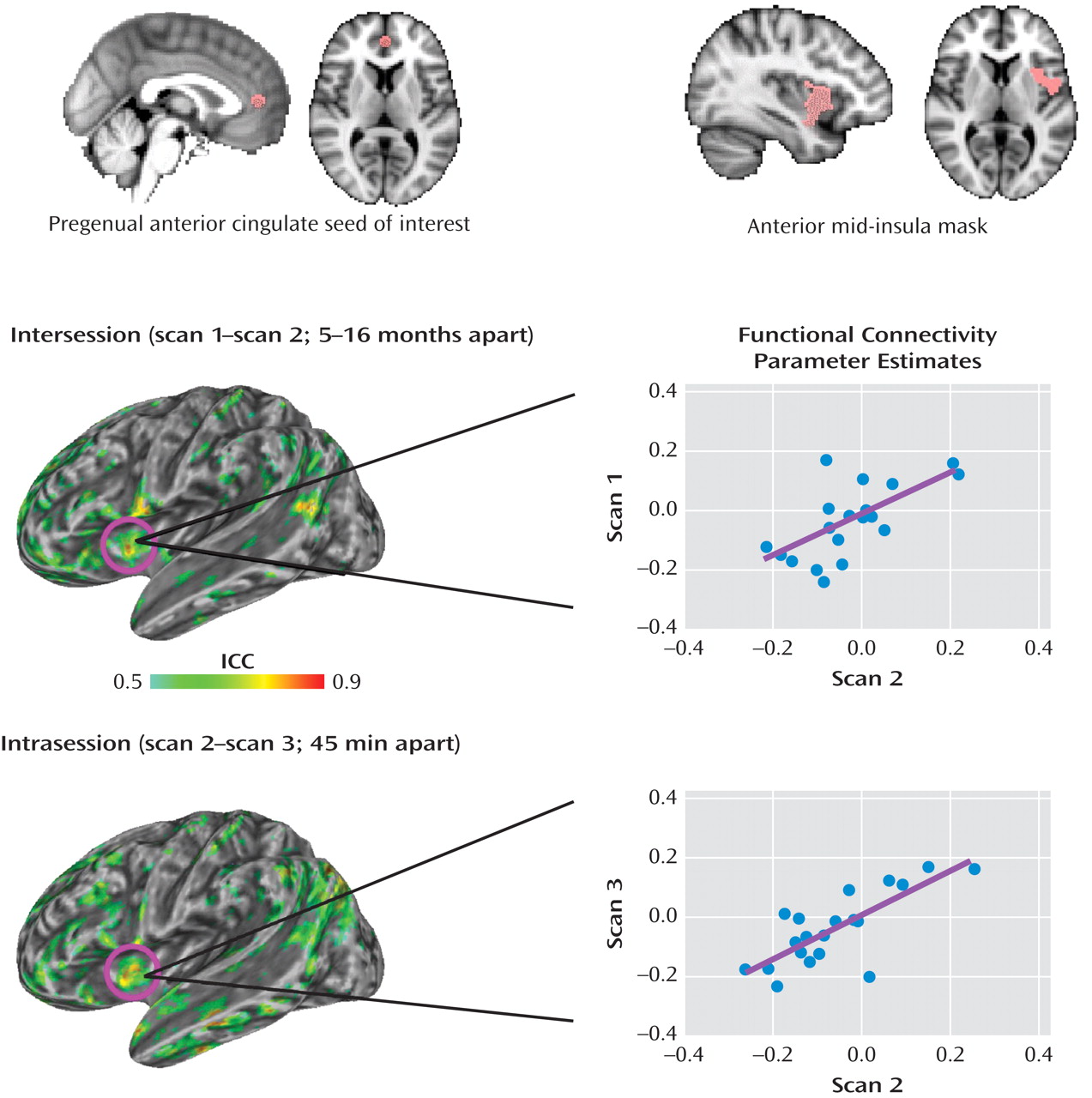
(39) . Further, our examination of the voxel-wise reliability of measures of functional connectivity within the pregenual anterior cingulate cortex network demonstrated moderate (intraclass correlation values >0.5) to high (intraclass correlation values between 0.7 and 0.95) test-retest reliability, a finding consistent with the stability of other resting state networks
(28) . Nevertheless, we cannot exclude the possibility that interindividual differences in responding to the scanner environment (e.g., scanner noise, experience of remaining still in the scanner) may be related to both patterns of functional connectivity and autistic traits assessed by the SRS-A. Future studies of the resting state using a combination of objective (e.g., physiological) and subjective (e.g., self-report) indices will be needed in order to clarify this issue.
Our findings should be interpreted in light of several limitations. Seed-based approaches to mapping functional connectivity require a priori selection of a region of interest. We selected the pregenual anterior cingulate cortex because of its prominence in models of autism-related abnormalities and its emergence in our recent meta-analysis of autism
(18) . Future studies will entail investigating the intrinsic connectivity of other regions of interest that may be implicated in the social impairments associated with autism
(18) . On a related note, although the SRS-A primarily emphasizes social and communication impairments in characterizing autistic traits, specifically focusing on other facets of autistic traits such as restricted/repetitive behaviors or interests may be helpful in future studies. We maximized the representativeness of individual parameter estimates by combining data from all available scans. Since only 80% of the study sample provided three scans, we considered the possibility that differences in the number of scans might have influenced our results. Accordingly, we accounted for potential differences at the group level by including the number of scans as a covariate. In addition, we conducted confirmatory analyses limited to 1) a single scan per subject and 2) the 20 participants who provided three resting state scans, and neither produced results that differed substantially from our main findings (see the data supplement accompanying the online version of this article). Finally, significant controversy continues to surround the interpretation of negative relationships in functional connectivity
(40) . Thus, although patterns of positive and negative connectivity differentiate insula subregions, caution should be taken when interpreting the meaning of negative relationships.
As has recently been noted
(41), when brain regions are identified by virtue of their correlation with behavioral measures, the corresponding correlation coefficients between the identified regions and the behavioral variables are generally somewhat inflated and should not be interpreted as unbiased. However, they do serve an illustrative purpose and are meaningful.
In summary, the present study demonstrates the utility of resting state fMRI for mapping functional connectivity in relation to the SRS-A, a continuous measure of autistic traits in the general population. This approach led us to identify the pregenual anterior cingulate cortex connectivity with the anterior mid-insula as a candidate marker of social competence related to autistic traits in a neurotypical sample. Application of this method in future studies examining patients with autism should allow confirmation of whether this circuit is a locus of dysfunction in autism that is dimensionally related to the severity of autistic traits.