The Interpersonal Dimension of Borderline Personality Disorder: Toward a Neuropeptide Model
Abstract
Abstract
Phenomenology of Borderline Personality Disorder
Interpersonal Dysfunction as a Core Component of Borderline Personality Disorder
Models of Interpersonal Dysfunction in Borderline Personality Disorder
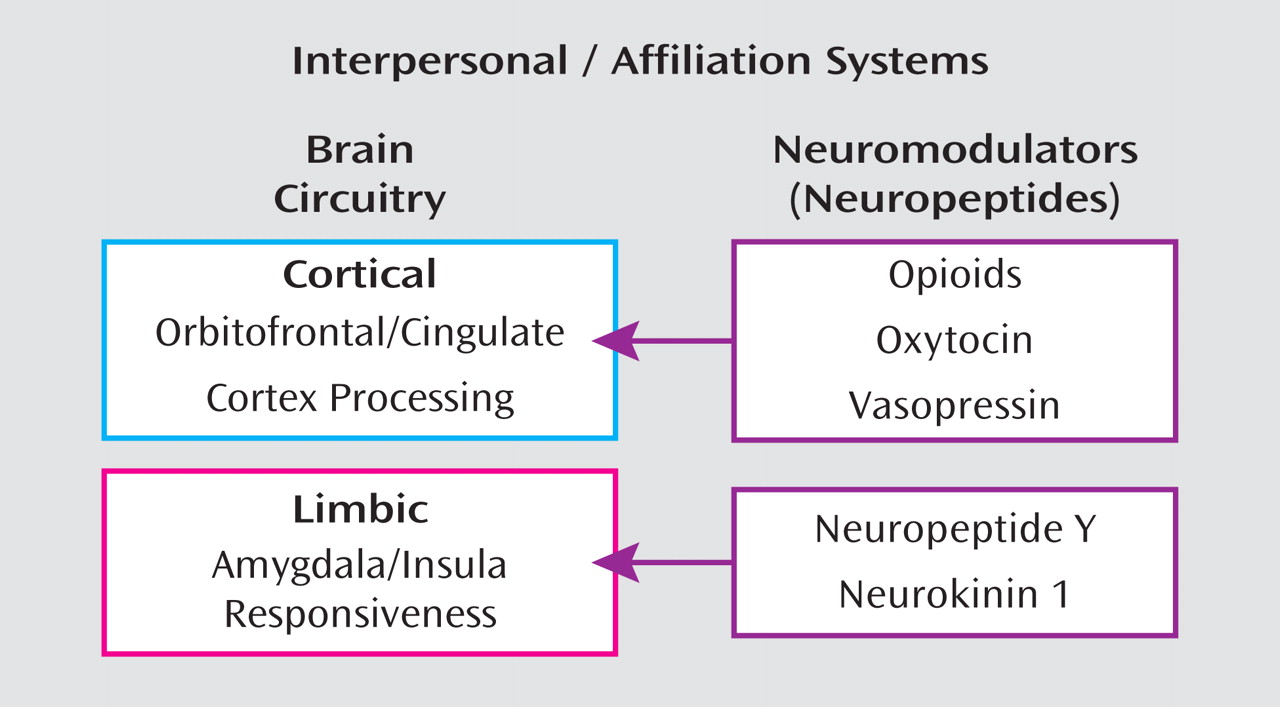
The Opioids and Borderline Personality Disorder
Neuropharmacology and Neurophysiology of the Opioid System
Opioids, Emotions, and Social Behavior
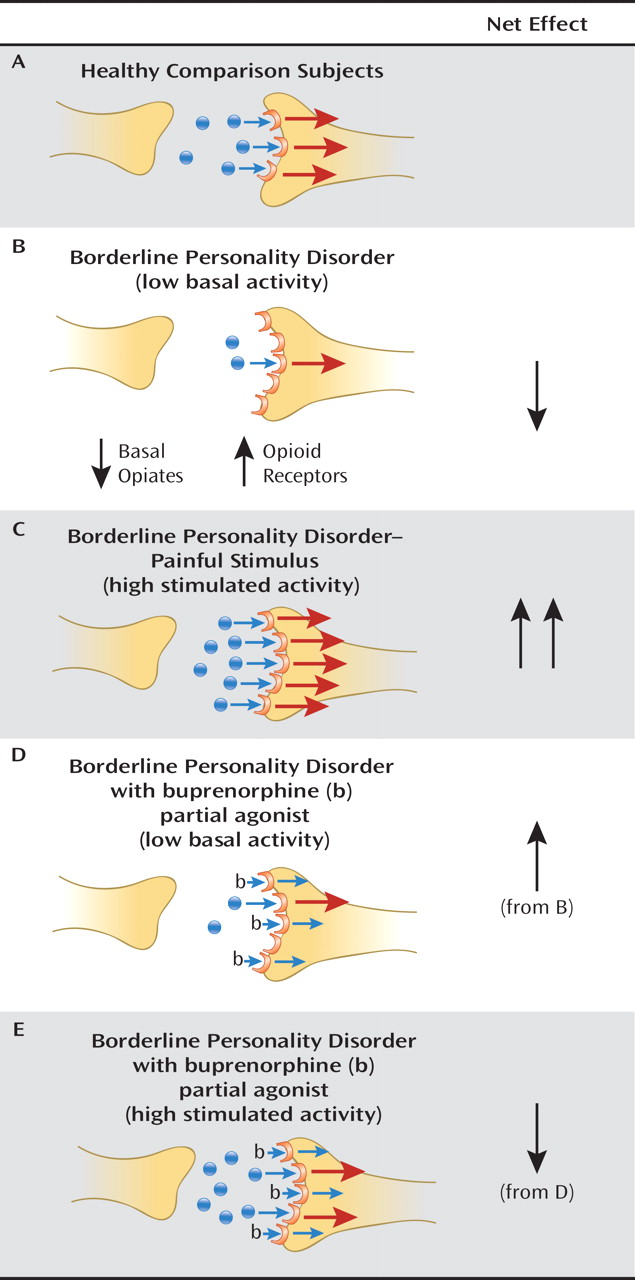
Opioid Dysfunction and Borderline Personality Disorder
Borderline personality disorder and physical pain.
Borderline symptoms and endogenous opioids.
μ-Opioid Imaging Studies in Borderline Personality Disorder
Individual Differences in Opioid Activity
Genetic Studies of the μ-Opioid Receptor, Attachment Behavior, and Borderline Personality Disorder
Model of Opioid Dysfunction in Borderline Personality Disorder
Oxytocin and Borderline Personality Disorder
Neuroanatomy and Neuropharmacology of the Oxytocin System
Oxytocin and Stress
Oxytocin, Affiliation, and Bonding
Oxytocin and Trust
Oxytocin Dysfunction in Borderline Personality Disorder
Individual Differences in Oxytocin Activity
Oxytocin, Genetics, and Borderline Personality Disorder
The Vasopressin System and Borderline Personality Disorder
Neuroanatomy and Neuropharmacology of the Vasopressin System
Vasopressin and Social Behavior
Vasopressin and Aggressive Behavior
Individual Differences in Vasopressin Activity
Model of Oxytocin and Vasopressin Activity in Borderline Personality Disorder
Implications for Research and Treatment
Research Implications
Treatment Implications
Conclusions
Acknowledgments
Footnote
References
References
Information & Authors
Information
Published In


History
Authors
Competing Interests
Funding Information
Metrics & Citations
Metrics
Citations
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
For more information or tips please see 'Downloading to a citation manager' in the Help menu.
View Options
View options
PDF/EPUB
View PDF/EPUBLogin options
Already a subscriber? Access your subscription through your login credentials or your institution for full access to this article.
Personal login Institutional Login Open Athens loginNot a subscriber?
PsychiatryOnline subscription options offer access to the DSM-5-TR® library, books, journals, CME, and patient resources. This all-in-one virtual library provides psychiatrists and mental health professionals with key resources for diagnosis, treatment, research, and professional development.
Need more help? PsychiatryOnline Customer Service may be reached by emailing [email protected] or by calling 800-368-5777 (in the U.S.) or 703-907-7322 (outside the U.S.).