Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by pervasive abnormalities in reciprocal social communication and stereotyped, repetitive behaviors. Difficulties with attention are also common (
1), and they contribute to the cognitive phenotype of ASD (
2) and have significant implications for the diagnosis and management of ASD and associated comorbidities (
3).
Comorbid attention deficit hyperactivity disorder (ADHD) is common and persists with age in individuals with ASD (
4,
5). Previously, diagnostic guidelines precluded diagnosing ASD and comorbid ADHD. However, DSM-5 allows this, enabling greater understanding of comorbid attention difficulties. People with ASD have been found to have difficulties with sustained attention (
6–
9), although there have also been negative findings (
10). Furthermore, it has been suggested that sustained attention may be an endophenotype that could help identify the neurobiological causes of ASD (
11). Understanding the neurofunctional differences underlying attention problems in ASD is important, as it may help elucidate objective biomarkers of ASD and potential targets for treatment development.
Nevertheless, despite evidence of poor sustained attention in individuals with ASD, only one published functional MRI (fMRI) study of sustained attention has shown dorsolateral prefrontal, striato-thalamic, and cerebellar underactivation in children with ASD relative to comparison subjects (
12). However, ASD is a neurodevelopmental disorder that persists across the lifespan. A key question is therefore whether functional brain abnormalities are present during childhood and adulthood, and if so, whether they are due to abnormal functional maturation of these networks.
There is evidence of abnormal longitudinal age-related changes in the brain structure of individuals with ASD in early childhood (
13,
14,
15) and adolescence (
16,
17), and cross-sectional studies report abnormal age-related differences in brain structure from childhood to adulthood (
18–
21). However, no developmental fMRI study has investigated whether functional brain maturation between childhood and adulthood differs in people with ASD from typical development during attention or any other cognitive function.
The aim of this cross-sectional developmental fMRI study was to investigate 1) the neural substrates underlying performance on a sustained attention/vigilance task of parametrically modified attention load in a relatively large group of healthy male adolescents and adults with ASD and age-, sex-, and IQ-matched typically developing healthy comparison subjects; and 2) age-related differences in neurofunctional maturation between childhood and adulthood in people with ASD relative to comparison subjects. The task we used requires subjects to respond to a visual stimulus appearing in either predictable short intervals (0.5 seconds) or unpredictable long intervals (2, 5, or 8 seconds). Long, infrequent, unpredictable delays place a higher load on sustained attention/vigilance than short, predictable delays (
12,
22).
fMRI studies of other sustained attention/vigilance tasks (for example, the continuous performance task) in typically developing adolescents and adults have shown activation in inferior and dorsolateral prefrontal, striato-thalamic, parieto-temporal, and cerebellar regions (
23–
25), which progressively increases with age between childhood and adulthood (
24–
26). We hypothesized 1) that individuals with ASD would show lower activation in fronto-striato-cerebellar sustained attention networks than typically developing comparison subjects and 2) that these functional deficits would be associated with abnormalities in underlying functional brain development between childhood and adulthood.
Method
Participants
A total of 90 physically healthy, medication-naive, right-handed males participated; 46 had ASD diagnoses and 44 were typically developing comparison subjects. Participants’ ages ranged from 11 to 35 years, and all had an IQ ≥70 (
27). Comparison subjects were recruited locally by advertisement and scored below cut-off for pathology on the General Health Questionnaire (GHQ) (
28), the Strengths and Difficulties Questionnaire (
29), and Conners’ Parent Rating Scale–Revised, Long Version (
30). Parents of children with ASD completed the Strengths and Difficulties Questionnaire and the Conners scale, and parents of adults with ASD completed the Barkley Parent Report (
31). Participants with ASD were recruited with support from the National Autistic Society and the Maudsley Hospital. ASD diagnosis was made by a consultant psychiatrist using ICD-10 research diagnostic criteria and confirmed using the Autism Diagnostic Interview–Revised (ADI-R) (
32). The ADI-R and the Autism Diagnostic Observation Schedule (ADOS) (
33) were completed for all 46 participants with ASD; all 46 reached algorithm cut-offs for autism in all domains on the ADI-R (social, communication, restricted/stereotyped) and the ADOS (communication, social). Participants with ASD either fulfilled ICD-10 research diagnostic criteria for childhood autism (N=14) or fulfilled these criteria but had no history of language delay and therefore were subtyped with Asperger’s syndrome (N=32) (
Table 1).
All participants underwent a structured clinical examination to exclude comorbid medical disorders, major psychiatric disorders, and biochemical, hematologic, or chromosomal abnormalities that might affect brain function. Exclusion criteria were comorbid psychiatric or medical disorders affecting brain development (e.g., epilepsy or psychosis), psychotropic medication (antipsychotics, stimulants, mood stabilizers, antidepressants, benzodiazepines), substance dependence, history of head injury, genetic disorder associated with ASD (e.g., fragile X syndrome, 22q11 deletion syndrome), or abnormal findings on head MRI.
Forty participants (20 in the comparison group, 20 in the ASD group) also participated in our study of sustained attention in pediatric ASD relative to ADHD (
12).
The local Ethics Committee conferred ethical approval, and written informed consent or assent was obtained from each participant. Each participant received £30.
Sustained Attention fMRI Task
Each participant practiced the task once in a mock scanner before scanning. The 12-minute sustained attention task (
12) is a variant of psychomotor/vigilance and delay tasks (
12,
22,
34) and requires subjects to respond as quickly as possible, with a right-handed button press, to a visual stimulus (a timer counting up in milliseconds) within 1 second. Subjects obtain implicit feedback by seeing the timer displaying the number of milliseconds it took them to respond to the timer. The timer appears after either short, predictable, consecutive delays of 0.5 seconds in series of three to five stimuli (240 trials total) or after unpredictable delays of 2, 5, or 8 seconds (20 trials each), pseudorandomly interspersed into the 0.5-second series after at least three predictable short delays (see Figure S1 in the
data supplement that accompanies the online edition of this article).
Analysis of Performance Data
Multiple repeated-measures analyses of variance (ANOVAs) (with group as the independent measure and delay as the repeated measure) were conducted to test for group performance differences, including mean reaction time, intrasubject response variability of reaction time (intrasubject standard deviation), omission errors, and premature responses.
fMRI Image Acquisition
fMRI images were acquired on a 3-T General Electric Signa HDx Twinspeed scanner (Milwaukee, Wisc.) using a quadrature birdcage head coil. In each of 22 noncontiguous planes parallel to the anterior-posterior commissure, 480 T2*-weighted images depicting blood-oxygen-level-dependent (BOLD) contrast spanning the entire brain were acquired (TE=30 ms, TR=1.5 seconds, flip angle=60°, in-plane resolution=3.75 mm, slice thickness=5.0 mm, slice skip=0.5 mm). A whole-brain structural scan (inversion recovery gradient echo planar image), on which to superimpose activation maps, was acquired in the intercommissural plane (TE=40 ms, TR=3 seconds, flip angle=90°, 43 slices, slice thickness=3.0 mm, slice skip=0.3 mm).
fMRI Data Analysis
Event-related activation data were acquired in randomized trial presentation and analyzed using the XBAM method of nonparametric data analysis (
35,
36). After preprocessing (see the online
data supplement), a time-series analysis of individual subject activation was performed (XBAM, version 4) with a wavelet-based fMRI data resampling method (
36). Using rigid body and affine transformation, individual maps were then registered in Talairach standard space (
37).
A group brain activation map was then produced for each experimental condition (long delays of 2, 5, or 8 seconds), each contrasted with the implicit baseline (frequent 0.5-second intervals). Only correct trials were included in the analyses. Hypothesis testing was carried out at the cluster level. A voxel-wise test at p<0.05 was conducted to identify any voxels that might plausibly be activated, followed by a subsequent test at a cluster-level threshold of p<0.01 to remove false positive clusters produced by the voxel-level test. Combined voxel/cluster tests with permutation testing allow for excellent type I error control (
36). For the group activation analysis, less than one false positive activated three-dimensional cluster was expected at p<0.05 in voxel-level comparisons and at p<0.01 in cluster-level comparisons.
For between-group comparisons of brain activation, a split-plot ANOVA design (three time delays, two groups) was conducted using a randomization-based test for voxel- or cluster-wise differences (
36). Less than one false positive activation cluster was expected at p<0.05 at voxel level and at p<0.01 at cluster level.
Whole-Brain Correlations Between Activation and Age Within Groups and Group Differences Between Whole-Brain Age Correlations
Because ANOVA showed that group differences increased progressively with delay, we used the longest delay (8 seconds) for the whole-brain age correlation analyses. To test for a linear correlation between whole-brain activation and age, the Pearson product-moment correlation coefficient was first computed at each voxel in standard space between age data and signal change across all subjects. Correlation coefficients were recalculated after randomly permuting the ages, but not the fMRI data. Multiply repeating the second step (1,000 times per voxel, then combining across all voxels) gives the distribution of correlation coefficients under the null hypothesis of no association between specific ages and specific BOLD effects. This null distribution can then be used to assess the probability of any particular correlation coefficient under the null hypothesis. The critical value of the correlation coefficient at any desired type I error level in the original (nonpermuted) data could be determined by reference to this distribution. Statistical analysis was extended to cluster level (
36).
To test whether group had differential effects on linear age correlations across the whole brain, group differences were examined in the correlation coefficients of brain activation with age. For each group independently, the average Pearson correlation coefficient between subject age and fMRI response was computed and group differences in correlation were calculated. To determine the significance of this difference, the appropriate null distribution was generated by randomly permuting subjects and ages between groups, thus scrambling any group differences. For each permutation, the correlation difference between scrambled groups was calculated and the resulting values were combined across all voxels to produce a whole-brain null distribution of differences in correlation. Testing was then extended to cluster level, with the cluster probability under the null hypothesis chosen to set the level of expected type I error clusters at less than one. Less than one error cluster was observed (at p<0.05 for voxel and p<0.01 for cluster analyses). Areas where either group showed exclusive significant progressive or regressive changes are reported.
To determine the direction of group differences in age correlations, post hoc analyses were conducted on the statistical measures of the BOLD response extracted for each subject in these regions, and age correlations were then performed for all clusters within each group.
Conjunction Analysis
To test whether areas of group differences were associated with differential neurofunctional development, we performed a conjunction analysis between the ANOVA group difference analysis and differences in whole-brain age correlations. Thus, we identified voxels where both the ANOVA group effect and between-group differences in whole-brain age correlations were significant (
38).
Results
There were no significant differences in age or IQ between the ASD and comparison groups.
Performance
Across all participants, there was a significant effect of delay on reaction time (F=124, df=3, 86, p<0.001), intrasubject standard deviation (F=8, df=3, 86, p<0.001), and omission errors (F=3, df=3, 86, p<0.036) (see Table S1 in the online
data supplement).
This analysis was repeated with age as a covariate. There was a significant effect of group on reaction time (F=12, df=1, 87, p<0.001) and intrasubject standard deviation (F=10, df=1, 87, p<0.002) as a result of the ASD group having a slower reaction time and greater intrasubject variability than the comparison group. No significant group-by-delay effect was observed.
Age Effects on Performance
Reaction time, intrasubject standard deviation, and premature responses were all significantly negatively correlated with age in both the ASD and comparison groups (r=−0.463, p<0.001) (see Table S1 in the
data supplement). There was a significant between-group difference in age correlation with reaction time only (p<0.001), because of significantly stronger age correlations in the comparison group (r=−0.645, p<0.001) relative to the ASD group (r=−0.409, p<0.005).
Movement
There was no significant group effect in the extent of three-dimensional motion as measured by maximum, minimum, or median displacement in the x, y, and z axes (see Table S2 in the online
data supplement).
fMRI Results
Brain activations within each group are shown in Figure S2 in the online
data supplement.
ANOVA group difference effect across all delays
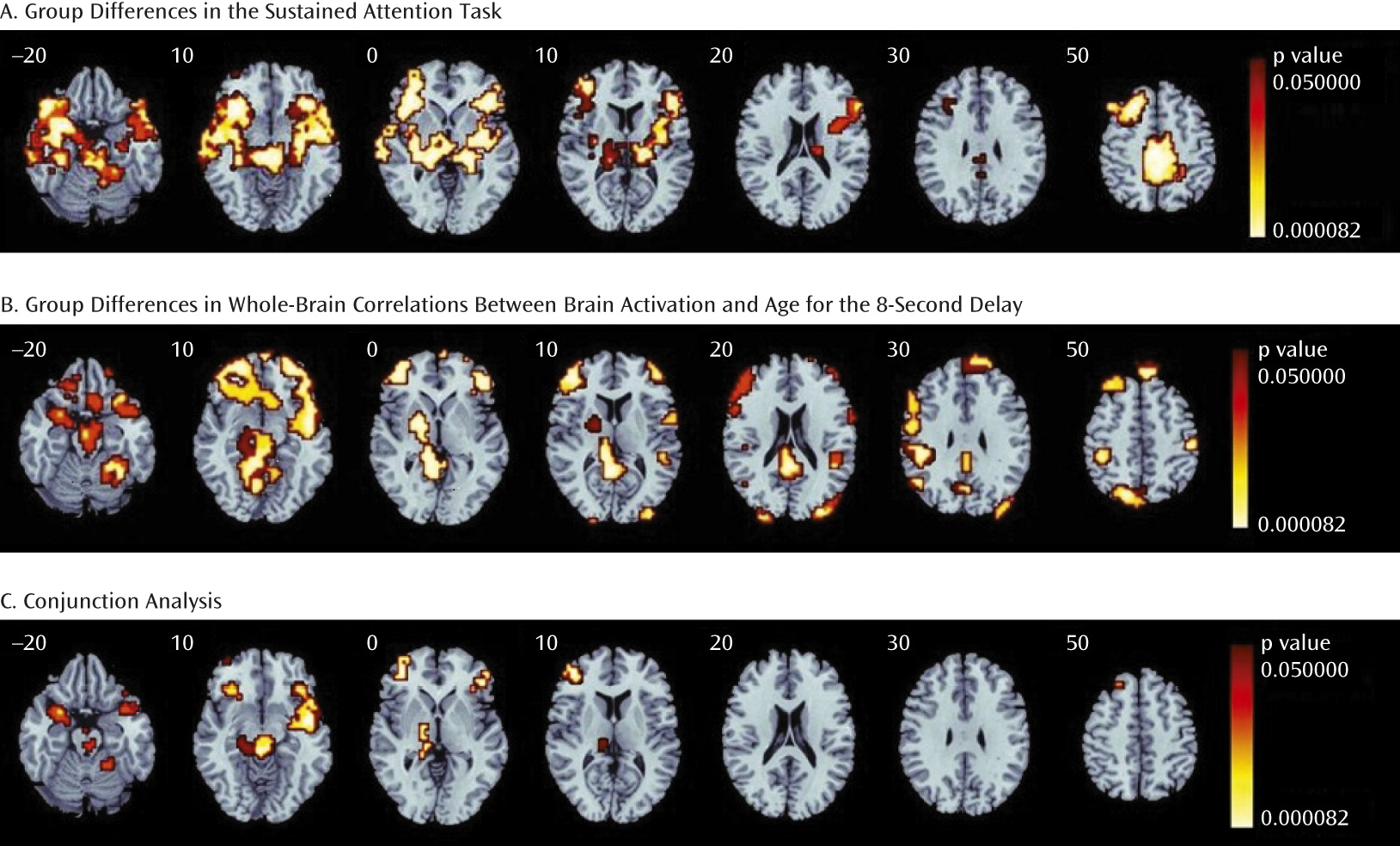
Across all delays, the comparison group, relative to the ASD group, showed higher activation in the left and right inferior frontal cortex reaching into the superior temporal lobe, left middle frontal cortex, insula, superior parietal cortex, posterior cingulate/precuneus, supplementary motor area, caudate, putamen, globus pallidus, thalamus, midbrain, and left and right cerebellar vermis and hemispheres (
Table 2,
Figure 1A).
Group differences in whole-brain correlations between brain activation and age for the longest delay.
Whole-brain correlations between age and brain activation individually for each group for the 8-second delay are shown in Figure S2 in the online
data supplement.
The group difference analysis between whole-brain age correlations showed that in several clusters, the comparison group had higher activation with increasing age, whereas in the ASD group, either this effect was diminished or negative correlations with age were observed in the left and right inferior frontal cortex-superior temporal junction, the left middle frontal gyrus/inferior frontal cortex, left and right superior and middle temporal and inferior and superior parietal regions, left striato-thalamic areas, the posterior cingulate, the midbrain, and the right cerebellar hemispheres and vermis (
Table 3,
Figure 1B).
Conjunction Analysis
The conjunction analysis showed five areas of overlap that both differed significantly between groups and increased significantly with age in the comparison group but not the ASD group: the right inferior frontal cortex-superior temporal junction reaching into the putamen; the left inferior frontal cortex reaching into the superior temporal lobe; the left middle frontal cortex; a cluster including the right thalamus, globus pallidus, and midbrain; and a right lateral cerebellar cluster (
Table 4,
Figure 1C).
Brain-Performance and Brain-Behavior Correlations
To investigate whether regions that survived the conjunction analysis were associated with performance or behavior, the statistical BOLD response in these regions was extracted for each subject and then correlated with reaction time for each group and, in the ASD group, with ADOS (communication, social) scores and z-transformed ADHD scores (z-transformed Conners’ Parent Rating Scale–Revised hyperactivity T-score [children] and Barkley self-report hyperactivity score [adults]).
In the comparison group, reaction time was significantly negatively correlated with activation in the right inferior frontal cortex-superior temporal lobe (r=−0.3, p<0.03), the left inferior frontal cortex (r=−0.3, p<0.05), and clusters in the left middle frontal cortex (r=−0.3, p<0.03). No significant correlations between performance and activation were observed in the ASD group.
In the ASD group, underactivation of the left inferior frontal cortex was negatively correlated with ADOS communication scores (r=−0.32, p<0.03), and underactivation of the right pallido-thalamic cluster was negatively correlated with z-transformed ADHD scores (r=−0.39, p<0.008).
To investigate the possible impact of group performance differences on group differences in brain activation and age correlations, we repeated both analyses using performance-matched subgroups (N=78); all findings remained identical except that the left middle frontal cortex and a small left temporal cluster no longer differed between subgroups (see Figures S4 and S5 in the online
data supplement).
Although motion did not differ between groups, to assess the possible impact of motion on group differences in activation or age correlation, we conducted a whole-brain correlation analysis between maximum motion displacement in the x, y, and z dimensions and brain activation for the 8-second versus 0.5-second contrast across all subjects. Two clusters were correlated with motion, but they did not overlap with the group differences in activation or age correlations (see the online
data supplement, including Figure S6).
Discussion
To our knowledge, this is the first fMRI investigation of differences in brain activation and cross-sectional functional brain development between healthy adolescents and adults with ASD relative to age- and IQ-matched typically developing comparison subjects during a sustained attention task. Relative to comparison subjects, individuals with ASD exhibited underactivation of left and right inferior frontal cortical/superior temporal cortical, left middle frontal cortical, striato-thalamic, and right lateral cerebellar regions. Our key novel finding is that abnormal activation in individuals with ASD in these areas was concomitant to underlying abnormalities in their functional brain maturation; with increasing age, the comparison group, but not the ASD group, showed progressively greater activation in these regions. Moreover, abnormal activation and functional maturation in inferior frontal cortical/middle frontal cortical/striato-thalamic regions in people with ASD was associated with poorer task performance and clinical measures of ASD and attention. These findings offer cross-sectional evidence that functional abnormalities in individuals with ASD during attention tasks may be related to underlying differences in the dynamic maturation of these functional networks between adolescence and adulthood.
The ASD group had significantly slower reaction times and larger intrasubject response variability, consistent with previous findings in children and adults with ASD during attention tasks (
6,
9). Inconsistent response times are associated with sustained attention problems (
39), while slower reaction times with increasing task delays suggest difficulties with sustained attention and stimulus anticipation (
40).
While comparison subjects showed progressively higher activation across all delays in a typical sustained attention network (left and right inferior frontal cortex; left middle frontal cortex; striato-thalamic, superior temporal, and inferior/superior-parietal regions; and lateral cerebellum) (
23), individuals with ASD had reduced activation in these regions across all delays, a finding that extends to adulthood previous evidence for lower activation in these regions in children with ASD during sustained attention (
12).
Most of these activations in left and right inferior frontal cortical, left middle frontal cortical, striato-thalamic, and cerebellar regions also showed a significant linear age correlation in the comparison group, supporting previous developmental fMRI attention findings (
24–
26,
41), but not in the ASD group. This demonstrates first cross-sectional evidence that abnormal inferior frontal cortical/middle frontal cortical-striato-thalamic and cerebellar brain activation in people with ASD during an attention task may be caused by abnormal functional maturation of these regions between childhood and adulthood. Furthermore, left and right inferior frontal cortical/superior temporal and left middle frontal cortical clusters that had lower activation and were functionally more immature in the ASD group were associated with better performance (shorter reaction time) in the comparison group, but not the ASD group. This suggests that in typically developing individuals these regions are recruited to perform better with increasing sustained attention/vigilance load, while poor task performance in ASD may be due to poor recruitment and “dysmaturation” of these frontal regions. Specifically, the left middle frontal cortex activation cluster was likely associated with performance differences, as it was no longer observed in performance-matched subgroups, while the bilateral underactivation and dysmaturation in the inferior frontal cortex survived the performance-matched subgroup analysis and may be more closely related to the ASD phenotype. Finally, fMRI evidence for abnormal functional maturation is reinforced by performance data, which showed a significantly stronger negative correlation between age and reaction time in the comparison group but not the ASD group.
These inferior frontal cortical/middle frontal cortical, superior temporal, striato-thalamic, and cerebellar attention regions that showed abnormal activation and functional maturation have also been shown to have abnormal structure and development in ASD (
19,
20,
42–
44), with evidence of time-specific regional changes—early abnormal overgrowth, followed by delayed growth in adolescence and adulthood, particularly in frontal and temporal regions (
18,
45).
Our cross-sectional findings of abnormal functional maturation in inferior frontal cortical/middle frontal cortical, superior temporal, pallido-thalamic, and lateral cerebellar regions in ASD are the first to extend evidence of a postadolescent abnormal structural maturation in these regions in ASD (
17,
19,
20,
44) to functional brain maturation. Interestingly, the regions found to be delayed in functional maturation in ASD are more susceptible to developmental delay, since they continue to develop across late adolescence into mid-adulthood (
46).
We also showed a correlation between the attention-mediating left middle frontal cortex (lower in both activation and neurofunctional development in the ASD group) and ADOS communication score, supporting the suggestion that attention contributes to the cognitive-behavioral phenotype of ASD (
2). Additionally, the underactivated left middle frontal cortical and left and right inferior frontal cortical-striatal clusters were correlated with better attention performance (shorter reaction time) in comparison subjects, suggesting an association in ASD between communication difficulties, poor attention performance, and underactivation and dysmaturation in the left middle frontal cortex/inferior frontal cortex. Intriguingly, the left middle frontal cortex, while crucial for attention (
47), has also been associated with social cognition and communication (
48).
Interestingly, only the underactivation of the right pallido-thalamic cluster correlated negatively with z-transformed ADHD scores in the ASD group. Striatal dysfunctions are key to ADHD (
49) and may be shared between ADHD and ASD (
12), possibly explaining the high overlap of attention problems. Furthermore, left middle frontal cortical and striato-thalamic underactivation was observed during the same task in both children with ADHD and children with ASD (
12). Sustained attention has major implications for learning and executive function (
50), matures late in adolescence (
51), and is a key cognitive impairment not only in ASD but also in numerous psychiatric disorders, particularly ADHD (
49). To our knowledge, however, there have been no cross-sectional or longitudinal fMRI studies in related psychiatric disorders. Future cross-sectional and longitudinal studies should investigate the specificity of the developmental underpinnings of attention deficits across disorders.
Limitations
Study limitations include the cross-sectional design, which is confounded by cohort effects; possible task confounding between trial duration and predictability; and a lack of measures of pubertal development. Only high-functioning males with ASD were included, which limits generalizability, and some people in our ASD group scored high on ADHD measures despite not having a clinical ADHD diagnosis. Although the findings survived the performance-matched subanalysis, we cannot rule out confounding due to subtle nonsignificant group differences in performance or strategies. A strength of the study is the relatively large sample of physically healthy, medication-naive, clearly diagnosed individuals with ASD without other psychiatric diagnoses and a 25-year age range from childhood to adulthood.