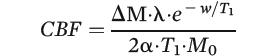Paranoia, defined as the “unfounded fear that others intend to cause you harm” (
1), is the most commonly reported delusional belief in individuals diagnosed with schizophrenia spectrum illnesses (
2). Such delusions are associated with significant distress and functional impairment (
3), often predict hospitalization (
4), and significantly predict conversion to psychosis in at-risk samples (
5). For these reasons, research has sought to understand the factors that contribute to and maintain paranoid ideation.
Whereas the psychological correlates of paranoia are fairly well understood (
2,
6), potential neurobiological bases remain unclear. Given the amygdala’s role in processing salience and threat (
7), investigators have postulated a link between amygdala activity and paranoia (
8–
10), and a review published in 2010 supports this relationship (
11). However, instead of seeing increased amygdala activation associated with paranoia as might be predicted, functional neuroimaging studies that have directly compared paranoid to nonparanoid individuals with schizophrenia have consistently reported amygdala hypoactivation in paranoid patients (
12–
17). In the studies of Williams et al. (
16,
17), decreased amygdala responses were paired with increased autonomic arousal (i.e., skin conductance), leading the investigators to hypothesize that a disjunction of arousal and amygdala-prefrontal neural circuits could lead to hypervigilance and maintenance of paranoid ideation. This is a viable explanation; however, an important caveat requires consideration. Each of the studies that have directly compared paranoid and nonparanoid patients has used the blood-oxygenation-level-dependent (BOLD) method. Estimates resulting from BOLD imaging are derived from a contrast between activity occurring during a specific event and that occurring during nonevents, or a baseline. These baselines cannot be directly measured and thus cannot be compared between groups. This raises the possibility that paranoid patients may actually show higher resting, or tonic, levels of amygdala activation and that the reported reductions in activation with task may result from high baseline activity that attenuates the event-evoked change detected by the BOLD method.
In the present study, we sought to test the hypothesis that amygdala hyperactivity would be present in paranoid individuals with schizophrenia by examining resting levels of cerebral blood flow (CBF) using arterial spin labeling imaging. Arterial spin labeling measures CBF using magnetically tagged arterial blood water as an endogenous tracer (
18,
19) and provides quantitative measurement of CBF that renders this method optimal for assessing baseline differences between groups. In order to replicate the previous BOLD findings, participants also completed a BOLD functional MRI (fMRI) trustworthiness rating task that we have used in our previous work and that robustly activates the amygdala (
13,
14,
20,
21). We predicted that the group of patients showing significant levels of paranoia at the time of scanning would show reduced task-related amygdala activation but greater resting CBF in the amygdala compared with both healthy comparison subjects and nonparanoid individuals with schizophrenia.
Method
Participants
The study included 57 participants from three groups: individuals with schizophrenia or schizoaffective disorder with prominent paranoid symptoms (paranoid schizophrenia, N=16), individuals with schizophrenia or schizoaffective disorder without paranoid symptoms (nonparanoid schizophrenia, N=16), and healthy community comparison subjects (N=25). All participants were between the ages of 18 and 55 years and proficient in English. To be eligible, participants could not report 1) a history of head injury or other medical conditions known to affect brain function (e.g., uncontrolled hypertension, diabetes mellitus, history of seizures), 2) pervasive developmental disorder, or 3) electroconvulsive therapy. Individuals also could not meet criteria for current substance abuse or dependence (except nicotine). The institutional review board of the University of Texas Southwestern Medical School approved the study protocol, and all participants provided written, informed consent.
Healthy comparison subjects were recruited through community advertisements in Dallas County and from other studies in our laboratory. Individuals were screened for personal and family history of psychopathology to ensure that they did not meet criteria for any DSM-IV axis I or II disorder and had no first-degree relatives who met criteria for a psychotic or affective disorder.
Individuals in the schizophrenia groups were recruited from the Southwestern Medical School Division of Translational Neuroscience of Schizophrenia and Metrocare Services, a nonprofit mental health services provider organization in Dallas County. Diagnoses were confirmed with the Structured Clinical Interview for DSM-IV (SCID-P). Severity of symptoms over the last week was assessed with the Positive and Negative Syndrome Scale (PANSS) (
22), and ratings on the suspiciousness/persecution item (item P6) were used to divide patients into paranoid and nonparanoid groups. Those scoring ≥4, indicating the presence of clinically significant paranoid ideation on the day of scanning, comprised the paranoid schizophrenia group, and those with a score of 1, indicating the absence of paranoid ideation, comprised the nonparanoid schizophrenia group. These groups did not differ on diagnosis (i.e., schizophrenia compared with schizoaffective; χ
2=2.0, p=0.16), medication type (i.e., typical compared with atypical; χ
2=0.96, p=0.62), or mean chlorpromazine equivalent dose (t=0.11, df=29, p=0.92) (
23). The paranoid schizophrenia group did show greater severity of positive (t=5.10, df=30, p<0.001) and general symptoms (t=3.52, df=30, p=0.001); however, these differences were no longer significant when controlling for the effects of paranoia. Severity of negative symptoms did not differ between groups (t=0.91, df=30, p=0.37). Finally, since previous work suggests that anxiety may be related to amygdala function (
24), we also compared groups specifically on level of PANSS-rated anxiety (item G2). The paranoid schizophrenia group (mean=3.94 [SD=1.53]) received higher ratings of anxiety than the nonparanoid group (mean=2.63 [SD=1.78]; t=2.24, df=30, p=0.03).
All three groups did not differ with regard to sex (χ
2=1.87, p=0.39), ethnicity (χ
2=6.50, p=0.17), age (F=1.34, df=2, 54, p=0.27), education (F=1.39, df=2, 54, p=0.26), or maternal education (F=1.24, df=2, 54, p=0.83). Paternal education significantly differed across groups (F=3.08, df=2, 54, p=0.049) such that the paranoid schizophrenia group showed lower attainment compared with the healthy individuals (p=0.04). Demographic and clinical characteristics of the study sample are presented in
Table 1.
Imaging Parameters and Procedures
Imaging was performed on a 3T Philips Achieva system (Philips Medical Systems, Best, the Netherlands) using the body coil for transmission and an eight-channel head coil for reception. A 4-minute magnetization-prepared, rapid acquisition gradient echo (160 slices, voxel size 1×1×1 mm, matrix=256×204, field of view=256×204, time to repeat/echo time=8.1 ms/3.7 ms) image was first acquired to obtain high-resolution anatomical images for spatial normalization. Pseudo-continuous arterial spin labeling was then used to obtain estimates of resting CBF (
25,
26). Forty control and label pairs of images were acquired with the following parameters: labeling duration=1,650 ms, postlabeling delay=1,525 ms, time to repeat/echo time=4,211 ms/14 ms, flip angle=90°, matrix size=80×79, voxel size=3×3×5 mm
3, 29 slices, no gap, duration=345 seconds. During scanning, participants were asked to relax, lie still, and keep their eyes open and focused on a small, white cross that was presented in the middle of a black screen.
Following the arterial spin labeling acquisition, participants completed an event-related BOLD imaging task comprised of 60 grayscale frontal images of faces taken from the Trustworthiness/Approachability Task (
27). Faces were displayed for 2 seconds and were followed by a variable interstimulus interval of 0.5 seconds–20.5 seconds during which participants fixated on a cross in the middle of a scrambled face. The total task duration was 8.5 minutes. As in previous work (
13,
14,
21), participants rated each face as either trustworthy or untrustworthy, and responses were recorded by button press. BOLD fMRI data were acquired during the task with the following parameters: 40 slices, voxel size=3.44×3.44×4 mm, matrix=64×64, field of view=220×220 mm, time to repeat/echo time=2,500 ms/30 ms.
Data Processing
Custom scripts utilizing Statistical Parametric Mapping (SPM2, University College London) were used to calculate CBF. The pseudo-continuous arterial spin labeling image series was initially realigned to the first volume. Pairwise subtraction between the label and control images was then performed to yield 40 difference images that were averaged to create one CBF image per participant using the following perfusion kinetic model:
In this perfusion kinetic model, w is the postlabeling delay time (1,525 ms), λ is the blood-brain partition coefficient (0.9 mL/g), and α is the labeling efficiency (0.86). T
1 (1,279 ms) is the averaged T
1 value of blood and tissue because the labeled spins spend some time in the blood and some time in the tissue space. M
0 is the equilibrium magnetization of the tissue estimated from the control image while accounting for the T
1 relaxation of the static spins (
28).
To reduce the potential influence of individual physiological differences, the CBF values at each voxel were normalized against the whole-brain gray matter value for each participant (
25). Thus, CBF values are provided as relative CBF. These averaged relative CBF images were coregistered to their corresponding structural image and then normalized to Montreal Neurological Institute (MNI) space using the standard template provided by SPM2 and smoothed with an 8-mm full-width at half-maximum three-dimensional isotropic Gaussian kernel.
Preprocessing and analyses of BOLD fMRI data were performed with FEAT (fMRI Expert Analysis Tool), version 6.00 (from FMRIB’s Software Library [FSL]). Images were slice-time corrected, motion-corrected to the median image, high-pass filtered (100 seconds), and spatially smoothed (8-mm full width at half maximum, isotropic). The median functional and anatomical images were coregistered and then transformed into standard space (T1 MNI template) using trilinear interpolation. Subject-level time-series statistical analyses were conducted using FILM (FMRIB’s Improved General Linear Model) with local auto-correlation correction. The events comprising each condition (trustworthy and untrustworthy) were identified according to the individual judgment of each participant, and the two conditions were then modeled with a canonical hemodynamic response function and its temporal derivative. Six rigid body motion parameters were also included as nuisance covariates. The resulting contrast images revealing stimulus-dependent activation relative to fixation baseline were then used in group-level analyses.
Data Analysis
Primary relative CBF analyses.
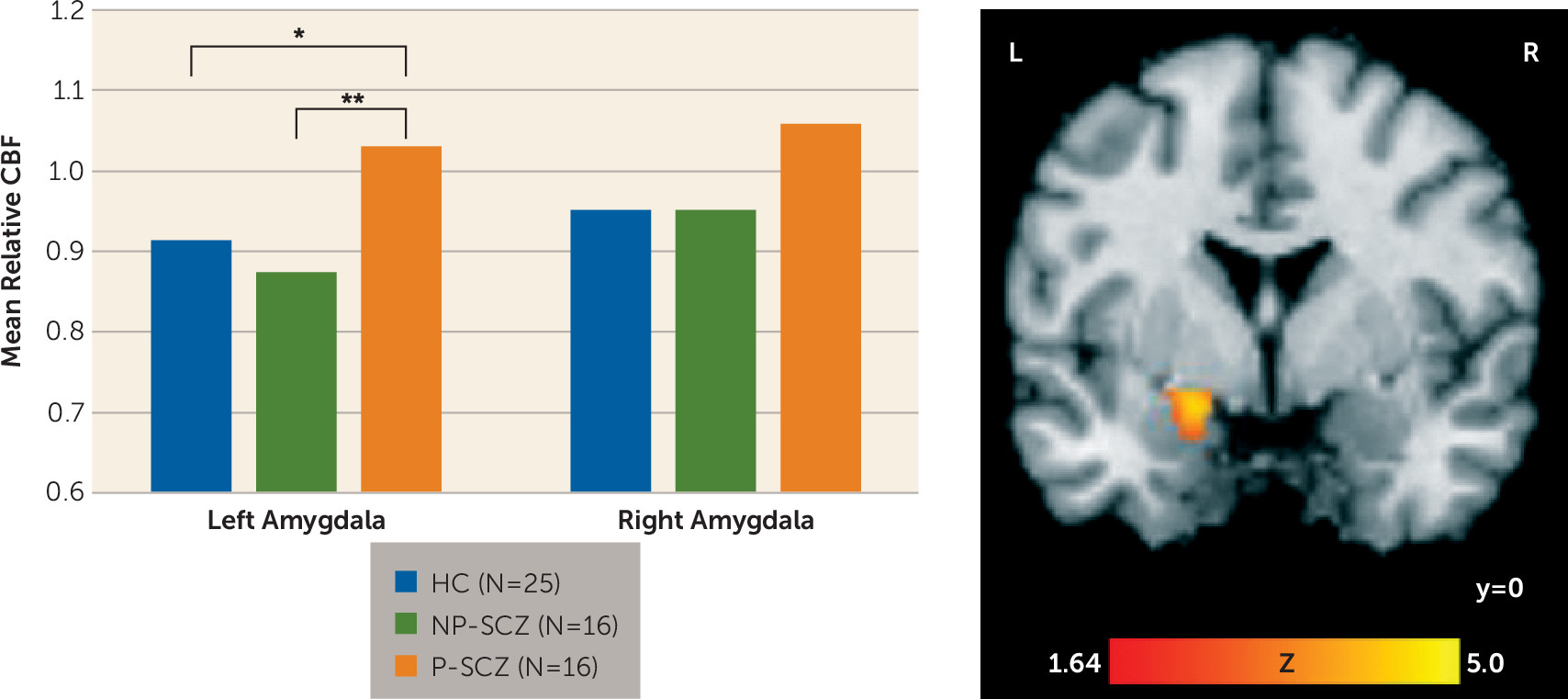
Both region-of-interest and exploratory whole-brain analyses were conducted. To examine activation in our a priori region of interest, the right and left amygdala were first anatomically defined using the Wake Forest University PickAtlas. Averaged estimates of relative CBF within the right and left amygdala were then extracted and entered into a repeated-measures analysis of variance (ANOVA) with hemisphere (right versus left) as the within-subject factor and group (healthy comparison compared with nonparanoid schizophrenia compared with paranoid schizophrenia) as the between-subject factor. Age, sex, and race were entered as covariates. The threshold for statistical significance was set at a p value <0.05, and significant group effects were probed with Tukey’s honest significant difference test.
In order to examine regions other than the amygdala in which paranoid and nonparanoid groups may differ in resting relative CBF, exploratory whole-brain voxel-wise analyses were also conducted with a one-way ANOVA (group: healthy comparison compared with nonparanoid schizophrenia compared with paranoid schizophrenia) conforming to random-effects analyses. As above, age, sex, and race were entered as nuisance covariates. Monte Carlo cluster correction (AlphaSim [
29]) at a p value <0.05 was used to correct for multiple comparisons. For interpretation, relative CBF values were extracted from clusters showing a main effect of group, and these values were analyzed with a one-way ANOVA with Tukey’s post hoc comparisons.
BOLD analyses.
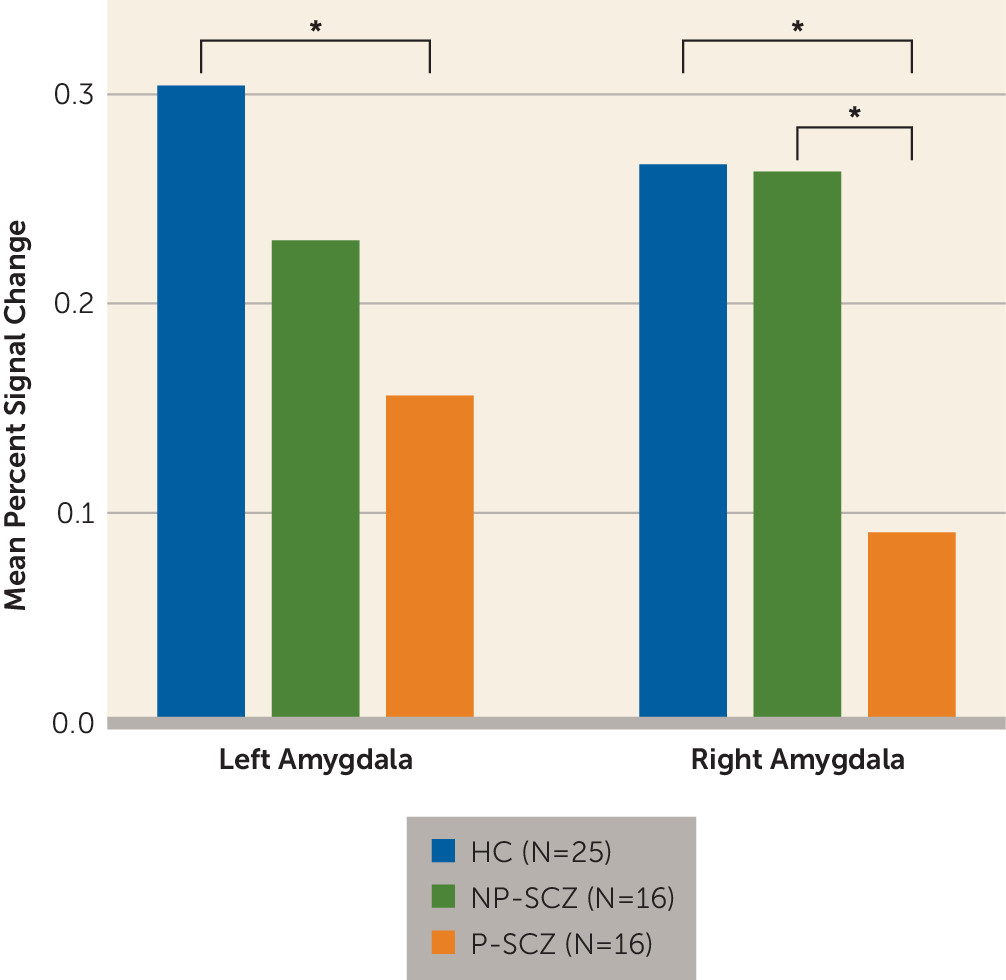
To examine group differences in task-related activation, mean percent signal change was extracted from the same regions of interest that were applied to the CBF data. These values were entered into a repeated-measures ANOVA with stimulus type (trustworthy versus untrustworthy) and hemisphere (right versus left) as within-subject factors and group (healthy comparison compared with nonparanoid schizophrenia compared with paranoid schizophrenia) as the between-subject factor. Effects with statistical probabilities with a p value <0.05 were considered significant and were followed up with Tukey’s post hoc tests. Behavioral data from the trustworthiness task are presented in the data supplement accompanying the online version of this article.
Supplemental analyses.
Given the link between anxiety and amygdala activation (
24) and the difference between patient groups in clinician-rated levels of anxiety on the PANSS, we also repeated the CBF region-of-interest analysis while controlling for this variable. Because healthy comparison subjects did not complete the PANSS, these analyses were restricted to the patient groups. Additionally, to assess the potential effects of morphological differences on the CBF results, a supplemental voxel-based morphometry analysis was also completed. Details of this analysis, including full results, are presented in the
data supplement.
Discussion
This study used arterial spin labeling imaging to assess potential differences in resting cerebral blood flow between paranoid and nonparanoid individuals with schizophrenia in an effort to clarify the association between amygdala activity and paranoid ideation. Patients who were actively paranoid at the time of scanning showed significantly increased blood flow in the left amygdala compared with both nonparanoid individuals with schizophrenia and healthy comparison subjects. Increases in resting CBF for paranoid individuals were also evident in the right amygdala, although to a lesser degree. Additionally, these same paranoid individuals showed reduced task-related amygdala activity compared with healthy comparison subjects in the bilateral amygdala and compared with nonparanoid patients in the right amygdala. The finding of increased baseline amygdala activity paired with decreased stimulus-driven activity may explain why previous BOLD comparisons of paranoid and nonparanoid patients have consistently, and counterintuitively, shown decreased amygdala activation in paranoid individuals. A higher resting baseline may create a ceiling effect that limits the signal change detectable by the BOLD method. Thus, the present findings suggest that paranoia is not related to reduced amygdala activity but rather that increased tonic activity of the amygdala may serve as a mechanism for paranoid ideation. This interpretation is consistent with the known link between the amygdala and threat perception and the conceptualization of paranoia as the continual perception of environmental threat.
Relating our findings to the broader literature on amygdala function in schizophrenia requires caution given that the vast majority of studies have not differentiated patients based on the presence of paranoia. However, one previous arterial spin labeling study did report hyperperfusion in the left amygdala of nonmedicated individuals with schizophrenia (
30), and positron emission tomography studies, which are not constrained by the same limitations as BOLD imaging studies, also provide support for increased amygdala activation at baseline in schizophrenia patients. Specifically, both Taylor et al. (
31) and Fernandez-Egea et al. (
32) reported hyperactivation of the amygdala in schizophrenia patients compared with healthy individuals. A meta-analysis of primarily BOLD studies also suggests that schizophrenia patients may show more amygdala activation than healthy individuals when processing neutral stimuli (
33), but how increased responding to neutral stimuli and increased baseline activity may fit together and how these relationships may differ between paranoid and nonparanoid patients requires further investigation. Liu et al. (
34) demonstrated that baseline CBF does influence the BOLD signal and that including estimates of baseline CBF can increase the sensitivity of BOLD analyses. Future investigations of amygdala function in schizophrenia may therefore benefit from concurrently measuring CBF and blood oxygenation.
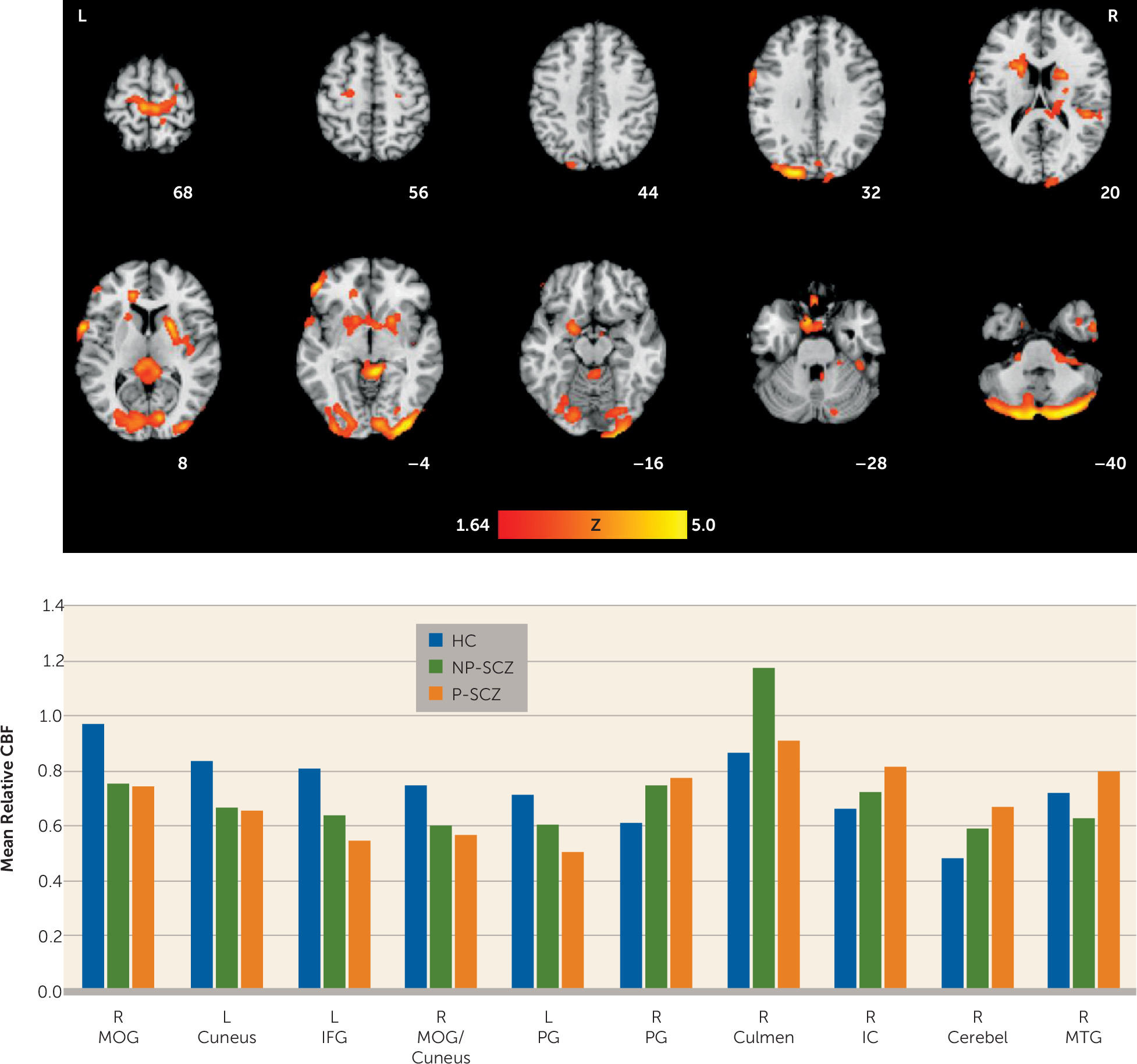
Results of the whole-brain analysis on CBF largely replicate previous arterial spin labeling studies that have compared schizophrenia patients with healthy individuals. As reported in the present study, increased CBF for healthy individuals compared with schizophrenia patients has previously been found in the middle occipital gyrus (
35,
36), fusiform gyrus (
37,
38), cuneus (
30,
37), and inferior frontal gyrus (
35,
36). Similarly, increased CBF for schizophrenia patients was noted in the precentral gyrus, cerebellum, and corpus callosum, all of which are consistent with previous reports (
30,
36). The whole-brain direct comparison between paranoid and nonparanoid patients is novel to the present study and extends the literature by highlighting the potential importance of white matter function for this symptom dimension. Paranoid patients, compared with nonparanoid patients, showed greater white matter CBF in the internal capsule, corpus callosum, and white matter portions of the inferior temporal and fusiform gyri. Collectively, these findings suggest that increased neuronal signaling at rest, particularly from sensory areas to frontal regions, may be associated with paranoid ideation. This interpretation is also consistent with our previous work showing a positive correlation between CBF in the corona radiata and the severity of positive symptoms (
36).
While our study establishes a link between increased amygdala activation and paranoia, some limitations should be considered. First, the results are correlational and do not provide a temporal ordering between amygdala activity and paranoid thinking. They also do not address the question of whether increased amygdala CBF is a state or trait characteristic of paranoid patients (i.e., does resting CBF fluctuate with paranoia or would it be evident in individuals with a history of paranoia but who are not currently paranoid). Longitudinal studies that can track CBF and symptom changes over time are needed. Second, the paranoid schizophrenia group reported higher levels of anxiety than the nonparanoid schizophrenia group. It is possible that increased anxiety may have contributed to the present results; however, this seems unlikely given that the difference between the patient groups in amygdala CBF was strengthened when statistically controlling for levels of anxiety. Nevertheless, anxiety has been shown to be a strong correlate of paranoia (as it was in our study [r=0.41 across all patients]) that could serve as a contributing factor or as a consequence (
6), suggesting that the potential role of anxiety should not be dismissed completely. Third, it could be argued that CBF differences between groups resulted from volumetric reductions in the patient groups. The limited overlap between our voxel-based morphometry and CBF results suggests that this is not the case. One possible exception could be the findings in the internal capsule. However, in this region, volumetric reductions were evident for both patient groups compared with healthy comparison subjects, whereas CBF differences were limited to the paranoid group.
Notwithstanding these limitations, the present findings identify increased resting cerebral blood flow in the amygdala in paranoid, compared with nonparanoid, individuals with schizophrenia and suggest that amygdala hyperactivity may subserve paranoid ideation. Given the negative effect of paranoia on the social functioning of individuals with schizophrenia, attenuating amygdala activation, through pharmacology for example, may improve quality of life. These results also highlight the need to consider symptom-based subgroups in future investigations of amygdala functioning in schizophrenia.