Anorexia nervosa is a serious eating disorder characterized by restriction of energy intake relative to requirements, resulting in abnormally low body weight. It has a lifetime prevalence of approximately 1% and disproportionately affects females (
1,
2), and no well-replicated evidence of effective pharmacological or psychological treatments for it have been identified, despite high morbidity and mortality (
3,
4). Twin studies consistently support a genetic basis for the observed familial aggregation in anorexia nervosa, with heritability estimates in the range of 48%–74% (
5). Although initial genome-wide association studies (GWASs) were underpowered (
6,
7), the available evidence strongly suggested that signals for anorexia nervosa would be detected with increased sample size (
6).
Our aim in the present study was to combine existing samples to conduct a more powerful GWAS of anorexia nervosa. To further characterize the nature of the illness, we applied linkage disequilibrium (LD) score regression (
8) to calculate genome-wide common variant heritability (single-nucleotide polymorphism [SNP]-based heritability [h
2SNP]), partitioned heritability, and genetic correlations (r
g) between anorexia nervosa and other phenotypes. These include the other major psychiatric disorders with large GWASs, namely schizophrenia, bipolar disorder, major depressive disorder, autism, and attention deficit hyperactivity disorder (ADHD), as well as medical, educational, and personality phenotypes. We then used r
g estimates between anorexia nervosa and 159 additional phenotypes to characterize the phenome-wide genetic architecture of anorexia nervosa.
Discussion
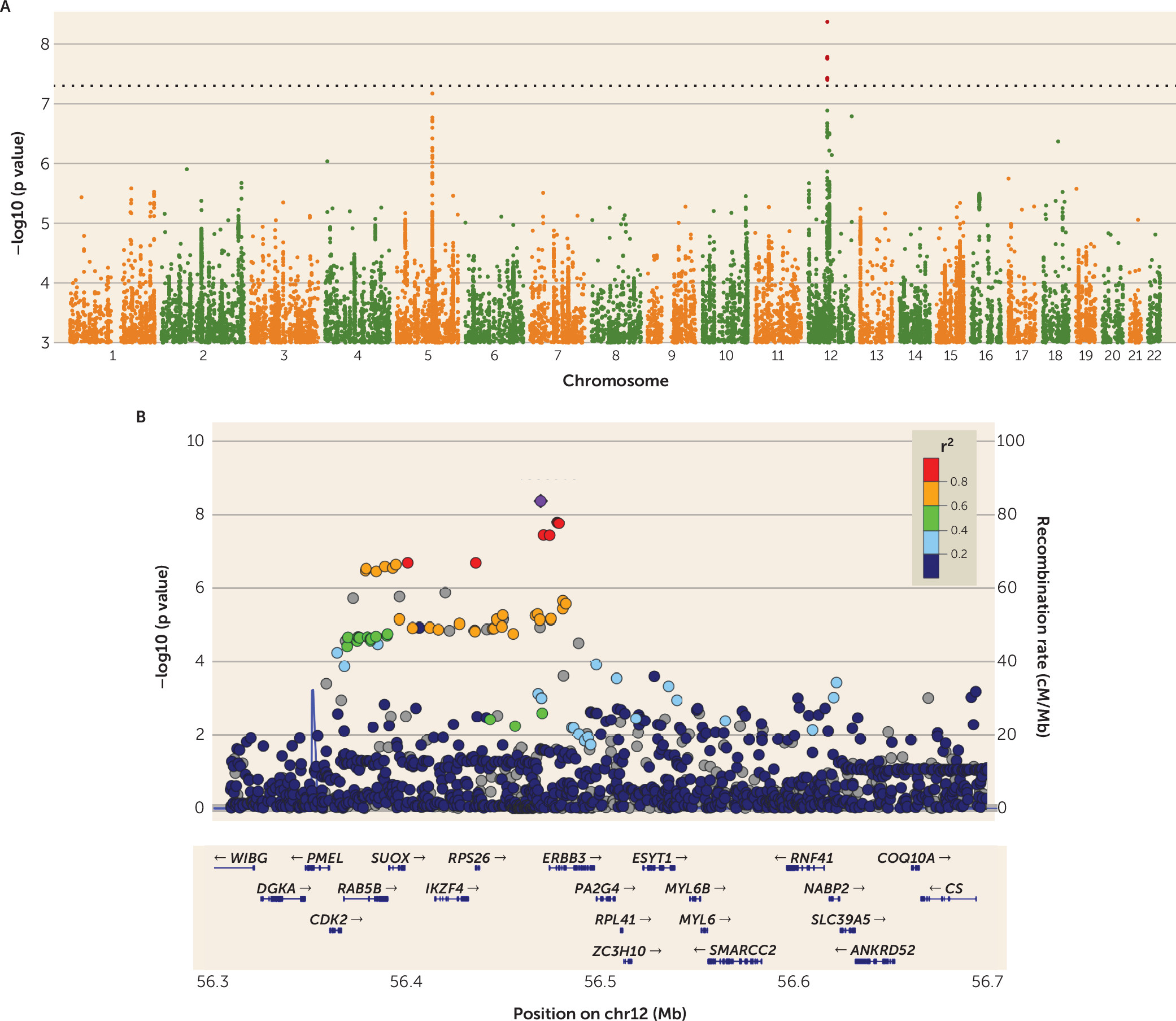
To our knowledge, this is the first report of a genome-wide significant association for anorexia nervosa. As is typical of many GWAS loci for complex disorders, the region has a common top variant (MAF
controls=0.44) that shows a modest odds ratio of 1.2 and implicates a broad region encompassing multiple genes (
25). Consistent with other GWAS (
26), our genome-wide h
2SNP estimate of 20% for anorexia nervosa supports a substantial role for common genetic variation, which accounts for a sizable portion of twin-based heritability (h
2Twin=48%–74%) (
6). Furthermore, these results fit with the expectation that h
2Twin should exceed h
2SNP, because the former captures the effects of all types of genetic variation (common and rare, as well as variation not captured with current methods).
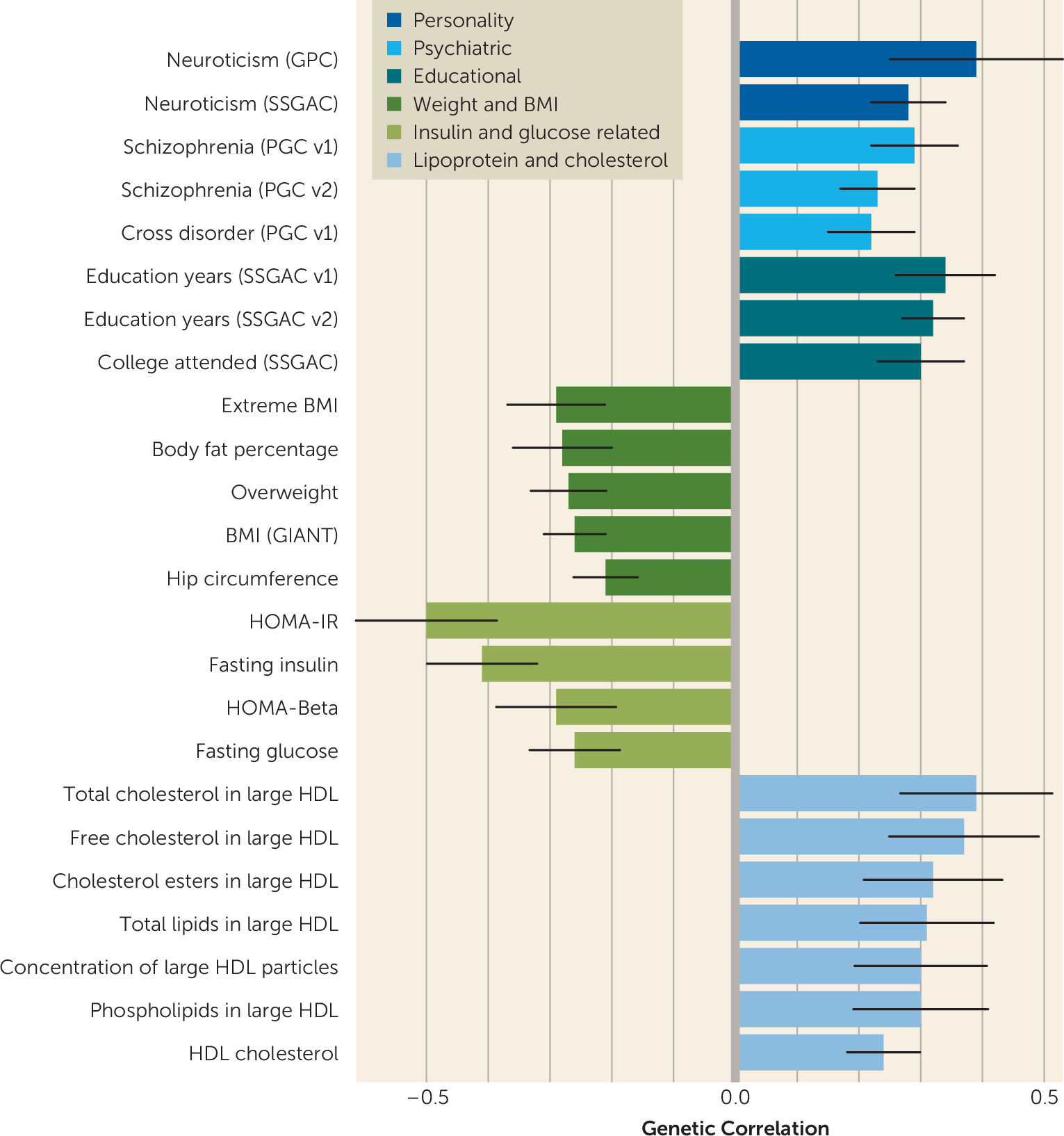
The observed pattern of genetic correlations with psychiatric, personality, educational, and metabolic phenotypes provides grounds for broadening our conceptualization of the disorder. First, the strong positive genetic correlations of anorexia nervosa with OCD and neuroticism reinforce clinical and epidemiological observations. Anorexia nervosa is commonly comorbid with OCD, and twin studies have reported high twin-based genetic correlations (
27). High neuroticism in adolescence predicts subsequent onset of anorexia nervosa (
1). In addition, anorexia nervosa is commonly comorbid with multiple anxiety phenotypes, which often predate the onset of anorexia nervosa (
28).
Second, the positive genetic correlations seen with schizophrenia and the cross–psychiatric disorder phenotype firmly anchor anorexia nervosa with other psychiatric disorders and reflect the substantial evidence for partially shared genetic risk across many psychiatric disorders (
29). Third, congruent with our results, positive associations between anorexia nervosa and educational attainment have been reported (
30) and have been conjectured to reflect greater internal and external demands for academic success in highly educated families. Our results, in contrast, suggest that genetic factors may partially account for these reported associations.
Fourth, the identification of significant negative correlations between anorexia nervosa and BMI-related and anthropometric measures could potentially serve as an important first step toward gaining a better understanding of the shared biology underlying extremes of weight dysregulation (i.e., obesity versus anorexia nervosa). This is of critical importance because adequate explanations for how individuals with anorexia nervosa reach, sustain, and revert to exceedingly low BMIs have been elusive. Clinically, one of the most perplexing features of anorexia nervosa is how patients’ bodies seem to revert rapidly to a “low set point” after renourishment, which may represent the biological inverse of the reversion to high set points commonly seen in the unsuccessful treatment of obesity (
31,
32). As noted by Bulik-Sullivan et al. (
23) and Hinney et al. (
33), these observations extend our understanding that the same genetic factors that influence normal variation in BMI, body shape, and body composition may also influence extreme dysregulation of these weight-related features in anorexia nervosa. This pattern of observations complements prior strong evidence for the involvement of neural mechanisms in obesity (
34). Finally, positive correlations with “favorable” metabolic phenotypes (i.e., HDL and lipid measures) and negative correlations with “unfavorable” metabolic phenotypes (i.e., fasting insulin level, fasting glucose level, HOMA-IR) encourage additional exploration of the role metabolic factors may play in extreme dysregulation of appetite and weight in anorexia nervosa.
The genome-wide significant locus we identify to be associated with anorexia nervosa is broad and multigenic (chr12:56,372,585–56,482,185). Mechanistic explanations about the role of the associated variant require additional functional data; nevertheless, we note the possible role for genes at this locus in the pathophysiology of anorexia nervosa.
PA2G4 is involved in growth regulation and acts as a co-repressor of the androgen receptor (
35).
ESYT1 (extended synaptotagmin-1, which binds and transports lipids [
36]) is enriched in the postsynaptic density, which is implicated in the etiology of schizophrenia (
37). Perhaps more convincing is that the sentinel marker for this locus, rs4622308, is in high LD with a known GWAS hit for type 1 diabetes (
20) and rheumatoid arthritis (
21), and the region around it harbors multiple other autoimmune associations. Multiple reports of shared effects between anorexia nervosa and immune phenotypes fit into a broader pattern of above-chance comorbidity across psychiatric and immune phenotypes (
38,
39). Evidence suggests that this shared risk is at least partly genetic in origin (
23,
39). A negative genetic correlation between anorexia nervosa and rheumatoid arthritis was previously reported (
23), and our LD score regression estimate—though only nominally significant—is in the negative direction as well (see Table S5 in the
data supplement).
The primary strength of this investigation is that it extends previous work by increasing sample size through collaboration. Nevertheless, contemporary understanding of complex trait genetics suggests that even larger samples are needed. Since our collection represents all of the currently GWAS-genotyped anorexia nervosa samples in the world, no known genotyped replication samples exist. We therefore expect this to be the beginning of genomic discovery in eating disorders (
25). Future work with additional and better-powered anorexia nervosa GWASs will clarify the magnitude of genetic relationships among metabolic and psychiatric phenotypes, and methods such as that proposed by Pickrell et al. (
40) will provide clues about the direction of causal relationships.
In summary, we identified the first robust genome-wide significant locus for anorexia nervosa, which is also a previously reported type 1 diabetes and general autoimmune disorder locus. Perhaps of greater importance is that we find anorexia nervosa to be a complex heritable phenotype with intriguingly large and significant genetic correlations not only with psychiatric disorders but also with multiple metabolic traits. This encourages a reconceptualization of this frequently lethal disorder as both psychiatric and metabolic. Just as obesity is increasingly considered to be both a metabolic/endocrine and psychiatric disorder, approaching anorexia nervosa as both a psychiatric and metabolic condition could ignite interest in developing or repositioning pharmacological agents for its treatment.