Increased Protein Insolubility in Brains From a Subset of Patients With Schizophrenia
Publication: American Journal of Psychiatry
Abstract
Objective:
The mechanisms leading to schizophrenia are likely to be diverse. However, there may be common pathophysiological pathways for subtypes of the disease. The authors tested the hypothesis that increased protein insolubility and ubiquitination underlie the pathophysiology for a subtype of schizophrenia.
Methods:
Prefrontal cortex and superior temporal gyrus from postmortem brains of individuals with and without schizophrenia were subjected to cold sarkosyl fractionation, separating proteins into soluble and insoluble fractions. Protein insolubility and ubiquitin levels were quantified for each insoluble fraction, with normalization to total homogenate protein. Mass spectrometry analysis was then performed to identify the protein contents of the insoluble fractions. The potential biological relevance of the detected proteins was assessed using Gene Ontology enrichment analysis and Ingenuity Pathway Analysis.
Results:
A subset of the schizophrenia brains showed an increase in protein insolubility and ubiquitination in the insoluble fraction. Mass spectrometry of the insoluble fraction revealed that brains with increased insolubility and ubiquitination exhibited a similar peptide expression by principal component analysis. The proteins that were significantly altered in the insoluble fraction were enriched for pathways relating to axon target recognition as well as nervous system development and function.
Conclusions:
This study suggests a pathological process related to protein insolubility for a subset of patients with schizophrenia. Determining the molecular mechanism of this subtype of schizophrenia could lead to a better understanding of the pathways underlying the clinical phenotype in some patients with major mental illness as well as to improved nosology and identification of novel therapeutic targets.
Schizophrenia affects about 1% of the general population and is characterized by positive and negative symptoms as well as cognitive deficits (1). Although our understanding of the genetics related to schizophrenia has advanced (2), the mechanisms of the disease are still poorly understood and are likely to be diverse. In addition, the current nosology of psychiatric illness is in flux; the concept of a categorical description has been challenged (3, 4), and even under a categorical nosological scheme, schizophrenia may not represent a single disease entity (5). Therefore, differentiating subtypes of schizophrenia may be valuable in understanding the mechanisms underlying disease phenotypes. One approach to subtyping schizophrenia is by clinical symptoms or course. However, this may not fully capture the biology that distinguishes patient subtypes. An alternative approach to improving patient subtyping is to identify pathological processes present in subsets of patients.
Abnormalities of protein conformation and solubility have been implicated in many disorders, including cancer, cardiac and pulmonary diseases, muscle diseases, and neurodegenerative disorders (6). We and others have previously shown that the protein products of genes with rare variants linked to schizophrenia in unique pedigrees are prone to insolubility (7, 8). Alterations in ubiquitin signaling and ubiquitin proteins have been found in homogenized brain tissue from schizophrenia patients (9, 10). This is of considerable potential significance because ubiquitin acts as a key modulator of protein insolubility (11), and ubiquitin reactivity is considered a marker for insoluble protein aggregates in neurodegenerative disorders. Based on these observations, we sought to test the hypothesis that protein insolubility and ubiquitination are present in a subset of brains from patients with schizophrenia and, if these were detected, to determine the potential biological relevance of the insoluble proteins.
Methods
Autopsy Brains, Rat Brains, and Sample Preparation
We conducted a pilot study, a full-scale study, and a replication study in human brain tissue, as well as an experiment in rat brain tissue. For the pilot study, blocks from the prefrontal cortex (Brodmann’s area [BA] 10) of autopsy brains frozen at −80°C were obtained from the Harvard Brain Tissue Resource Center. For the full-scale study, autopsy brains were provided by the University of Pittsburgh Brain Bank. Gray matter was harvested as previously described (12): tissue slabs containing the superior temporal gyrus, excluding Heschl’s gyrus and planum temporale, were collected in 40-μm sections and frozen at −80°C. A sample of autopsy brains for a replication study was provided by the University of Texas Southwestern Medical Center (13): tissue slabs containing the prefrontal cortex (BA 46) were collected and frozen at −80°C. These brain regions were chosen for their relevance to schizophrenia (14–23). While the brain tissues are not all from the same area because of differences in the availability of tissue from the different brain banks, they represent a pan-cortical sample of brain tissue. For all three brain banks, available covariates were tested for statistically significant differences between patient and control tissues, using unpaired two-tailed t tests for continuous variables and Fisher’s exact test for discrete variables (see Tables S1, S3, and S4 in the online supplement ). Experimenters were blinded to patient or control status during all steps of sample preparation and analysis for all brain cohorts. For the rat brain experiments, male Sprague-Dawley rats were treated with haloperidol (1.5 mg/kg per day), risperidone (6 mg/kg per day), or water only in their drinking water continuously for 4.5 months, as described previously (24). The rats were then sacrificed by decapitation, and the dorsal striatum/caudate area was dissected and frozen by immersion in −40°C isopentane and stored at −80°C until use. Three rats were used for each condition. Experimenters were blinded to treatment group during all steps of sample preparation and analysis.
Brain Fractionation Protocol
A 10% (weight/volume) homogenate was prepared from human brain tissue in 50 mM Hepes at pH 7.5, 250 mM sucrose, 5 mM MgCl2, 100 mM KCH3COO, 1% Triton X-100, and 1× protease inhibitor cocktail, then centrifuged (20,000 ×g, 20 minutes, 4°C). The pellet was resuspended and centrifuged (130,000 ×g, 50 minutes, 4°C) in 3 mL of high-sucrose buffer (50 mM Hepes at pH 7.5, 1.6 M sucrose, 100 mM KCH3COO, 0.5% Triton X-100, 1 mM phenylmethanesulfonyl fluoride) followed by overnight DNase treatment (100 units/mL) in 3 mL of high-salt buffer (50 mM Hepes at pH 7.5, 1 M NaCl, 20 mM MgCl2, 30 mM CaCl2, 100 Units/mL DNase, 1× protease inhibitor cocktail) at 4°C. The sample was centrifuged (130,000 ×g, 50 minutes, 4°C). The resultant pellet was resuspended and centrifuged (112,000 ×g, 50 minutes, 4°C) in 300 μL of sarkosyl buffer (50 mM Hepes at pH 7.5, 0.5% N-lauryl-sarcosine) to obtain the final insoluble fraction. Each of the above steps was repeated twice. The final pellet was resuspended in 100 μL of 2× Invitrogen NuPage LDS Sample Buffer (56 mM Tris HCl, 70.5 mM Tris base, 1% lithium dodecyl sulfate [LDS], 5% glycerol, 0.255 mM EDTA, 0.11 mM SERVA Blue G250, 0.0875 mM Phenol Red, pH 8.5, 3% 2-mercaptoethanol [BME]).
Western Blot and Quantification Protocol
All insoluble fractions were subjected to SDS-PAGE analysis followed by Coomassie/silver staining and Western blot analysis. The gels were processed on a Criterion TGX 4%−15% precast gel (Bio-Rad Laboratories, catalog no. 567-1084). For Coomassie staining, SimplyBlue SafeStain (Invitrogen, catalog no. LC6060) was used. For silver staining, the SilverQuest Staining Kit (Life Technologies, catalog no. LC6070) was used. Both Coomassie staining and silver staining were used to quantify levels of total homogenate protein and protein insolubility in the final pellet. For Western blot analysis, the gels were transferred to a 0.45 µM PVDF transfer membrane (Immobilon-P, Millipore). A polyclonal anti-ubiquitin antibody produced in rabbit (Dako, code Z0458) was used as the primary antibody for processing the Western blots at a concentration of 1:1000. An anti-rabbit peroxidase-linked secondary antibody (GE Healthcare) was used at a concentration of 1:5000.
Quantification of the protein insolubility was performed by measuring the protein signal of the entire lane for each sample processed on the Coomassie stained polyacrylamide gels and subtracting background signal. The final signal was corrected for the total homogenate protein concentration of each processed sample. The entire area of ubiquitin signal starting from the top of each lane was quantified. The final ubiquitin signal was corrected for total homogenate protein concentration of each processed sample. All gel experiments were repeated with consistent results.
Liquid Chromatography–Mass Spectrometry Data Acquisition
Insoluble pellets were solubilized in 4% SDS 100 mM Tris at pH 7.4 with protease and phosphatase inhibitors (Sigma), digested by trypsin using the FASP method (25), and analyzed by liquid chromatography–tandem mass spectrometry on an Orbitrap XL Hybrid Ion Trap-Orbitrap Mass Spectrometer (Thermo Scientific) operated in data dependent mode, with a nanoACQUITY UPLC system (Waters) and a picochip column (New Objective), as previously described (26).
Label-Free Quantification and Protein Identification
All preliminary data processing was performed in Proteome Discoverer, 2.2 beta versions, and finalized in version 2.20.388 (Thermo Fisher Scientific) using an ion current–based label-free quantification method similar to that previously described (27). Briefly, the total ion chromatogram was subjected to feature alignment using all present ions to compensate for chromatographic shift. Ion traces were stored and matched between runs if they met the post-alignment criterion that retention times within 120 seconds between runs for ions possessing at least three detectable isotopes fell within a 10-ppm mass tolerance. The area under the curve was extracted for the dominant isotope and the trace stored in the output file. Normalization was applied using the total ion current available in each output file and scaled to a total value of 100. All ion abundance measurements were adjusted by this scaling. The normalized protein output was visually inspected to verify algorithm accuracy. The initial analysis contained all pooled technical and digestion controls for further verification of fitness for quantification analysis.
Identification of peptides was performed with SequestHT using a maximum 10-ppm mass tolerance for the parent ion and a 0.6-Da fragment tolerance for tandem mass spectrometry. All data were searched against the cRAP contaminant database (http://www.thegpm.org/crap/) and the UniProt SwissProt Human canonical database (downloaded on April 10, 2017). Carbamidomethylation of cysteines was considered as a static modification; acetylation of the protein N-termini and oxidation of methionine were applied as potential variable modifications. Multiple testing correction was performed using false discovery rate calculations, as previously described (28). A 1% false discovery rate cutoff was applied to both the peptide spectral matches (calculated using Percolator [29]) and peptide group levels. Quantification ratios for each peptide were determined via pairwise analysis of individual peptides and then averaged for peptide group and protein levels. Significance was then determined by analysis of variance based on peptide background at both the peptide group and protein levels (30).
Statistical Analysis
Baseline demographic and biochemical characteristics of brain samples from schizophrenia patients were compared with those of control subjects in each of the three brain bank cohorts. Differences between the groups were assessed using Student’s t test for continuous variables and Fisher’s exact test for categorical variables (unpaired and two-tailed). After biochemical analyses of protein insolubility and ubiquitin reactivity, hierarchical clustering analysis was performed on the z-score-normalized concentration of protein insolubility and ubiquitin reactivity in R (hclust function with “average” method; dist function with “euclidean” method; version 3.3.0). After dichotomization into brains with or without increased protein insolubility, the demographic characteristics were compared between patients with increased insolubility and patients and controls without increased insolubility. This was then repeated for patients with increased insolubility compared only to patients without increased insolubility. An unpaired two-tailed t test was used to determine whether the patients with increased insolubility were significantly different from the other patients and controls. The same analysis was applied to ubiquitination data.
Pathway and Cell Type Enrichment Analysis
In an effort to identify dysregulated pathways and cell types contributing to or resulting from protein insolubility, proteins identified by mass spectrometry were analyzed with enrichment tests. Pathways were assessed using gene set enrichment analysis (GSEA; Gene Ontology biological processes gene sets with PANTHER algorithm) and Ingenuity Pathway Analysis (IPA; Qiagen; accessed August 2015). Cell type enrichment was performed using the Expression Weighted Cell Type Enrichment (EWCE) R package (version 0.99.2) and two human cortical single-cell RNA sequencing (RNA-seq) data sets. The first data set, by Darmanis and colleagues (31), contains expression data for neurons, oligodendrocytes, oligodendrocyte progenitor cells, astrocytes, microglia, and endothelial cells. The second data set, by Lake and colleagues (32), contains expression data for inhibitory and excitatory neuron subtypes that they identified (N=8 each). EWCE determines the probability of enrichment and standard deviations from the bootstrapped mean for each cell type, using bootstrap significance testing with 20,000 randomly generated gene lists for comparison while controlling for transcript length and GC content. False discovery rates were calculated from the resulting p values using the Benjamini-Hochberg method, and cell types were considered enriched when the false discovery rate was <0.05.
Results
Demographic Information for the Pilot and Full-Scale Studies
For our pilot study, we analyzed samples from five schizophrenia patients and four control subjects from the Harvard Brain Tissue Resource Center matched for age, sex, and postmortem interval (PMI) (see Table S1 in the online supplement ). Because this was a pilot study, the demographic data were more limited than for the larger cohorts of the full-scale and replication studies.
For the full-scale study, we analyzed 19 brains from schizophrenia patients (brains S1–S19) and 19 brains from control subjects (brains C1–C19) from the University of Pittsburgh Brain Bank, matched for age, sex, race, PMI, RNA integrity number (RIN), and storage time (see Table S3 in the online supplement ). Brain pH was significantly lower in the patient group than in the control group (p=0.006). Patients S4, S6, and S10 died by suicide.
Increased Ratio of Insoluble Protein to Total Protein and Increased Ubiquitination in a Subset of Schizophrenia Brains in the Pilot and Full-Scale Studies
To determine whether protein solubility is disrupted in some patients with schizophrenia, we used two markers of protein abnormality, insolubility and ubiquitination. Protein insolubility is often a consequence of abnormal protein tertiary structure. One of the effects of ubiquitination (addition of ubiquitin to proteins) is to render proteins insoluble (11). Therefore, to examine protein insolubility and ubiquitination, we performed cold sarkosyl fractionation, an established method (7, 33–38) that uses detergent and gradient fractionation to separate soluble and insoluble protein fractions. We quantified the amount of protein insolubility as well as the amount of ubiquitin reactivity in the insoluble fraction. We initially performed this experiment in a small number of brains from the Harvard Brain Tissue Resource Center as a pilot study and determined that a subset of patients with schizophrenia have significant increases in these two markers (see Figure S1 in the online supplement ). Based on these results, we obtained a larger set of brain samples from the University of Pittsburgh Brain Bank. Using this sample, we first demonstrated that joint analysis of protein insolubility and ubiquitination detects a group of brains from schizophrenia patients that appear to separate from all other patients and control subjects based on these two markers (Figure 1A). Therefore, hierarchical analysis was performed. Unlike flat clustering algorithms, which try to divide a data set into a predefined number of groups, hierarchical clustering yields a tree diagram to show the “natural” grouping structure of the data. Our hierarchical clustering analysis identified two clusters based on protein insolubility and ubiquitination (cluster 1 and cluster 2) (Figure 1B). We determined that 10 out of 19 schizophrenia brains from the University of Pittsburgh Brain Bank (cluster 2, brains S10–S19) cluster separately from brains of all other patients and control subjects based on protein insolubility and ubiquitination (Figure 1B). A direct comparison demonstrates that brains in cluster 2 have significantly more protein insolubility and ubiquitination in their insoluble fractions than those in cluster 1 (p<0.001) (Figure 1C–E). Patient brain S1 was excluded from the positive cluster by hierarchical cluster analysis because it had high levels of protein insolubility but low levels of ubiquitination.
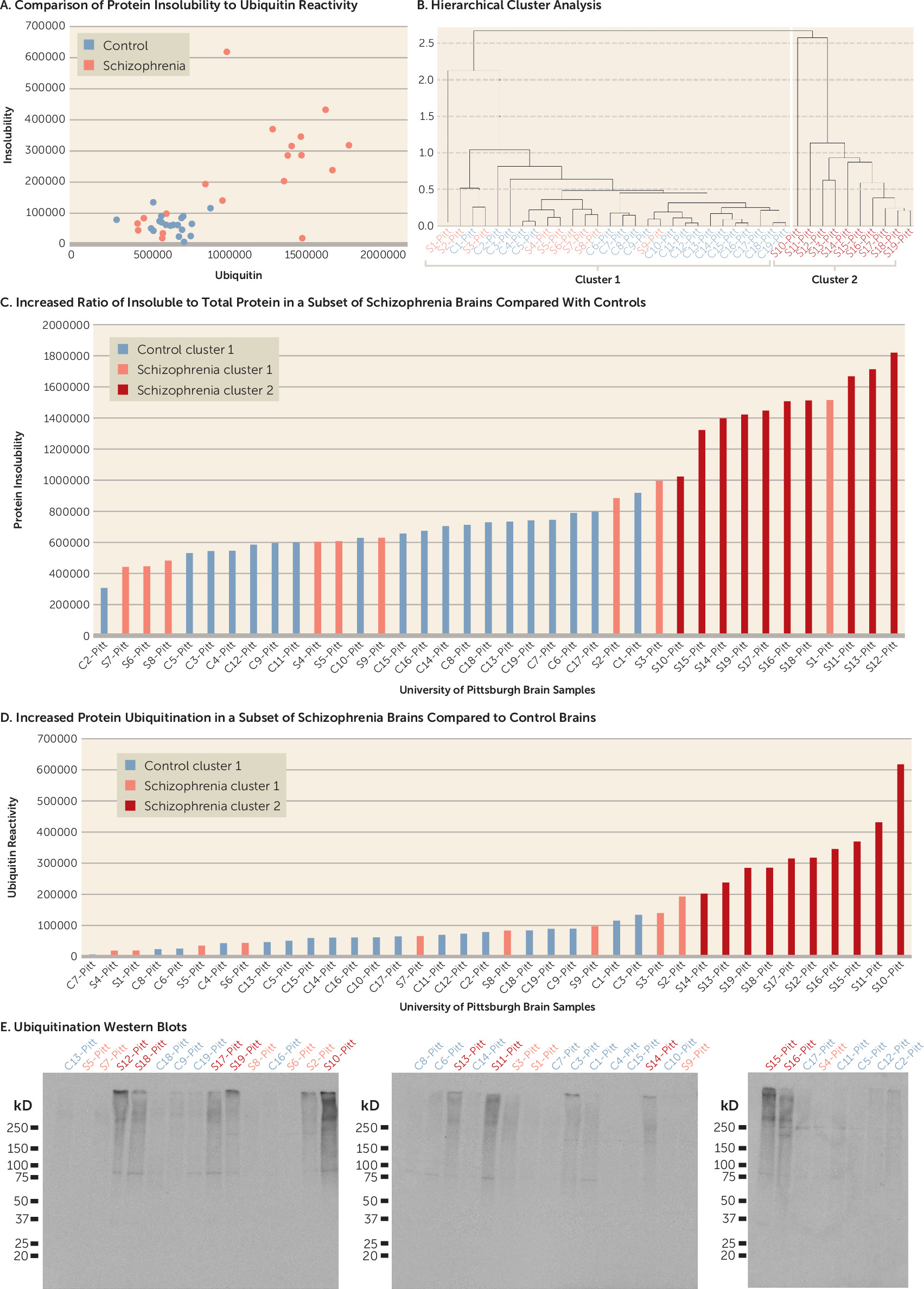
FIGURE 1. Increased ratio of insoluble protein and ubiquitination to total protein in a subset of schizophrenia brains compared with control brains from the full-scale study (University of Pittsburgh Brain Bank)a
a Cold sarkosyl fractionation was used to obtain insoluble fractions from postmortem brains of 19 schizophrenia patients and 19 control subjects from the University of Pittsburgh Brain Bank. In panel A, when insolubility and ubiquitination measurements were jointly analyzed, a subset of patient brains separated from other patient and control brains. In panel B, hierarchical clustering analysis determined that 10 of the schizophrenia brains (brains S10–S19, cluster 2) clustered separately from all other patient and control brains. The y-axis corresponds to dissimilarity. In panel C, schizophrenia brains in cluster 2 have significantly more protein insolubility than the patient and control brains in cluster 1 (p≤0.001). In panel D, the schizophrenia brains in cluster 2 were also significantly different from the patient and control brains in cluster 1 based on the amount of ubiquitination in the insoluble fraction (p≤0.001). Blue represents control brains with low levels of the markers of protein abnormality (insolubility and ubiquitination), pink indicates schizophrenia brains with low levels of these markers, and red indicates schizophrenia brains with high levels of these markers, based on hierarchical cluster analysis. Brain S1 was excluded from the positive cluster by hierarchical cluster analysis because it had a high level of protein insolubility but a low level of ubiquitination. In panel E, the ubiquitination was strikingly higher in the subset of schizophrenia brains with increased markers (cluster 2) than in the brains of the other patients or control subjects (cluster 1).
As an additional control experiment, we determined that the total amount of starting protein was not significantly different between the mean of three representative patients with increased protein insolubility and ubiquitination and the mean of three representative controls. We also detected a shift in the amount of protein from the soluble fraction to the insoluble fraction in the patient brains with increased protein insolubility and ubiquitination. When the soluble and insoluble fractions were combined, they added back to approximately the same amount of total starting material, consistent with the presence of an abnormality in protein insolubility in these brains rather than in the total amount of protein (see Figure S2 in the online supplement ).
Correlations Between Demographic Characteristics and Increased Protein Insolubility and Ubiquitination in the Pilot and Full-Scale Studies
In our pilot study using the small Harvard Brain Tissue Resource Center sample, differences in age, sex, and PMI between patients with increased markers of protein abnormality and all other patients and control subjects, or between patients with increased markers and patients without increased markers, did not reach statistical significance (see Tables S2A and S2B in the online supplement ).
For our full-scale study with brains from the University of Pittsburgh Brain Bank, the differences in age, sex, race, PMI, RIN, or storage time between patient brains with increased insolubility and ubiquitination, compared with all other patient and control brains, did not reach statistical significance (Table 1). However, the pH in brains with protein abnormalities was slightly but significantly lower (p<0.001). The differences in age, sex, race, PMI, RIN, storage time, or antipsychotic use between patients with increased insolubility and ubiquitination compared with patients without increases in these markers did not reach statistical significance. Brain pH was again slightly lower (p=0.029) and anticonvulsant exposure was higher (p=0.033) in patient brains with protein abnormalities.
| All Brains: Positive vs. Negative Cluster | Schizophrenia Brains: Positive vs. Negative Cluster | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Characteristic | Cluster 1 (Negative) (N=28) | Cluster 2 (Positive) (N=10) | Cluster 1 (Negative) (N=9) | Cluster 2 (Positive) (N=10) | ||||||
| N | % | N | % | p | ||||||
| Group | <0.001 | |||||||||
| Control | 19 | 67.9 | 0 | 0.0 | ||||||
| Schizophrenia | 9 | 32.1 | 10 | 100.0 | ||||||
| Mean | SD | Mean | SD | p | Mean | SD | Mean | SD | p | |
| Age (years) | 46.3 | 11.2 | 49.6 | 13.6 | 0.453 | 47.8 | 12.4 | 49.6 | 13.6 | 0.765 |
| Postmortem interval (hours) | 18.1 | 7.0 | 17.4 | 7.8 | 0.789 | 19.2 | 8.1 | 17.4 | 7.8 | 0.627 |
| RNA integrity number | 8.3 | 0.5 | 8.0 | 0.7 | 0.171 | 8.3 | 0.6 | 8.0 | 0.7 | 0.446 |
| Brain pH | 6.7 | 0.2 | 6.3 | 0.3 | <0.001 | 6.6 | 0.3 | 6.3 | 0.3 | 0.029 |
| Storage time (months) | 150.2 | 45.2 | 147.2 | 27.9 | 0.845 | 143.6 | 47.7 | 147.2 | 27.9 | 0.839 |
| N | % | N | % | p | N | % | N | % | p | |
| Sex | 0.404 | 0.184 | ||||||||
| Male | 22 | 78.6 | 6 | 60.0 | 8 | 88.9 | 6 | 60.0 | ||
| Female | 6 | 21.4 | 4 | 40.0 | 1 | 11.1 | 4 | 40.0 | 0.650 | |
| Race | 0.832 | 5 | 55.6 | 7 | 70.0 | |||||
| White | 19 | 67.9 | 7 | 70.0 | 4 | 44.4 | 3 | 30.0 | ||
| Black | 8 | 28.6 | 3 | 30.0 | 0 | 0.0 | 0 | 0.0 | ||
| Other | 1 | 3.5 | 0 | 0.0 | 0.184 | |||||
| Medication use | ||||||||||
| Any | 8 | 88.9 | 10 | 100.0 | 0.474 | |||||
| Antipsychotic | 7 | 77.8 | 9 | 90.0 | 0.582 | |||||
| Anticonvulsant | 0 | 0.0 | 5 | 50.0 | 0.033 | |||||
| Antidepressant | 4 | 44.4 | 4 | 40.0 | 1.000 | |||||
| Benzodiazepine | 1 | 11.1 | 1 | 10.0 | 1.000 | |||||
| Other | 6 | 66.7 | 7 | 70.0 | 1.000 | |||||
a
Clusters were determined using hierarchical analysis.
Demographic Information for the Replication Study
We obtained a third set of brain samples from the University of Texas Southwestern Medical Center to replicate our findings from the University of Pittsburgh Brain Bank samples. The samples (18 schizophrenia brains, S20-S37, and 18 control brains, C20-C37) were matched for age, sex, race, RIN, pH, PMI, and storage time. As expected, psychotropic medication exposure was higher in the patient samples than in the control samples (see Table S4 in the online supplement ). Patients S20, S23, and S32 and control subject C23 died by suicide.
Increased Ratio of Insoluble Protein to Total Protein and Increased Ubiquitination in a Subset of Schizophrenia Brains in the Replication Study
Consistent with our findings in brains from the Harvard Brain Tissue Resource Center and the University of Pittsburgh Brain Bank, eight out of 18 brains from schizophrenia patients (cluster 2, brains S30–S37) clustered separately from the brains of all other patients and control subjects (Figure 2A and 2B). Again, the brains of cluster 2 tended to have the highest amount of protein insolubility and ubiquitination, and mean levels of protein insolubility and ubiquitination were significantly higher than in brains of cluster 1 (p<0.001 and p=0.002, respectively) (Figure 2C, 2D, 2E). Hierarchical cluster analysis excluded brains S23 and S24 because they had high levels of protein insolubility but low levels of ubiquitination, and brain S25 because it had a high level of ubiquitination but a low level of protein insolubility.
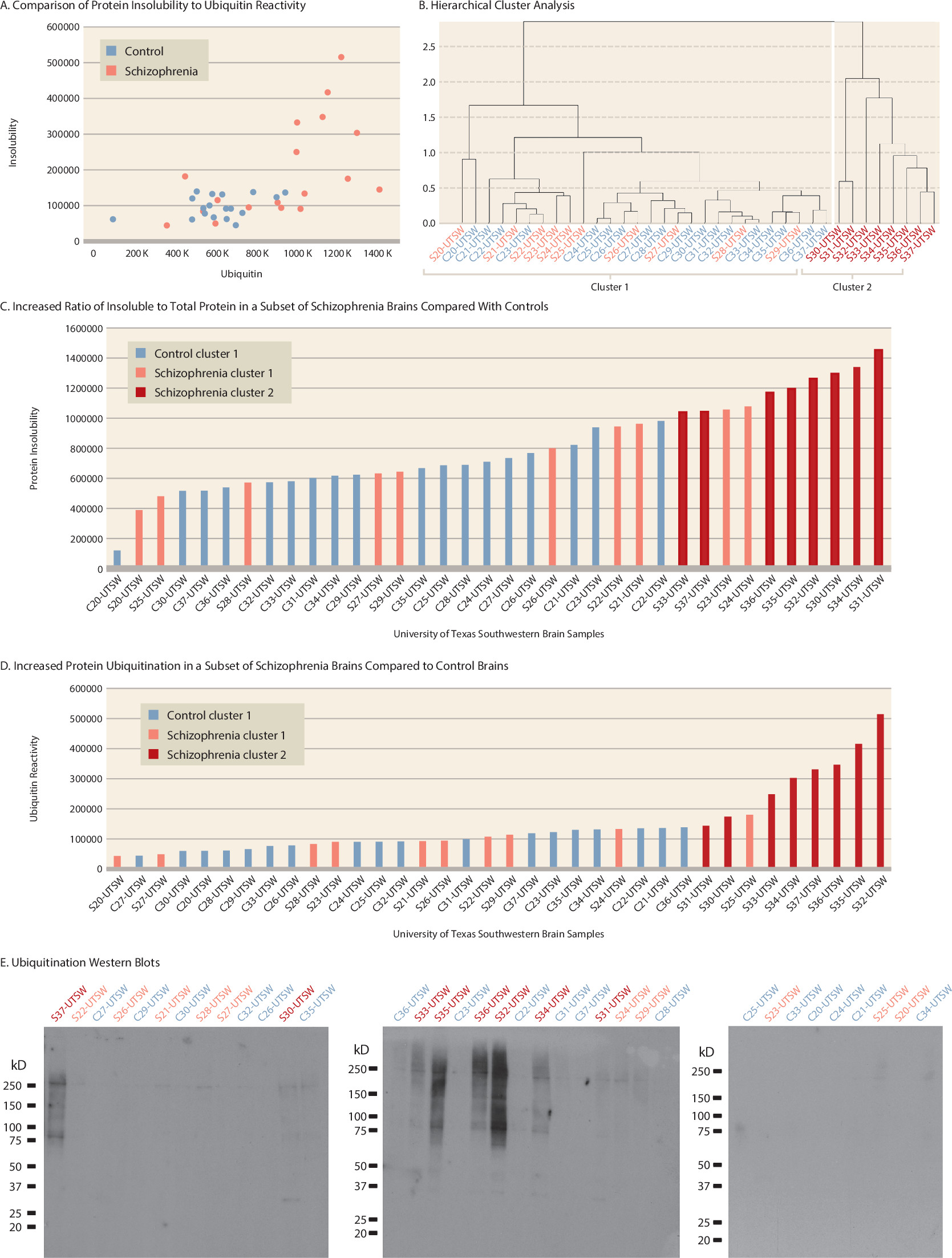
FIGURE 2. Increased ratio of insoluble protein and ubiquitination to total protein in a subset of schizophrenia brains compared with control brains from the replication study (University of Texas Southwestern Medical Center)a
a Cold sarkosyl fractionation was used to obtain insoluble fractions from postmortem brains of 18 schizophrenia patients and 18 control subjects from the University of Texas Southwestern Medical Center (UTSW). In panel A, when insolubility and ubiquitination measurements were jointly analyzed, a subset of schizophrenia brains separated from all other patient and control brains. In panel B, hierarchical cluster analysis determined that eight of the schizophrenia brains (brains S30–S37, cluster 2) clustered separately from all other patient and control brains. The y-axis corresponds to dissimilarity. In panel C, schizophrenia brains in cluster 2 have significantly more protein insolubility than the patient and control brains in cluster 1 (p<0.001). In panel D, the schizophrenia brains in cluster 2 were also significantly different than the patient and control brains in cluster 1 based on the amount of ubiquitination in the insoluble fraction (p<0.001). Blue represents control brains with low levels of the markers of protein abnormality (insolubility and ubiquitination), pink indicates schizophrenia brains with low levels of these markers, and red indicates schizophrenia brains with high levels of these markers based on hierarchical clustering analysis. Hierarchical clustering analysis excluded brains S23 and S24 because they had high levels of protein insolubility but low levels of ubiquitination, and brain S25 because it had a high level of ubiquitination but a low level of protein insolubility. In panel E, the ubiquitination was strikingly higher in the subset of schizophrenia brains with increased markers (cluster 2) than in the brains of the other patients or control subjects (cluster 1).
No Correlation of Demographic Variables With Increased Protein Insolubility and Ubiquitination in the Replication Study
Age, sex, race, PMI, RIN, brain pH, and storage time did not differ between patient brains with increased insolubility and ubiquitination compared either to all other patient and control brains or to brains of patients without increased markers (Table 2).
| All Brains: Positive vs. Negative Cluster | Schizophrenia Brains: Positive vs. Negative Cluster | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Characteristic | Cluster 1 (Negative) (N=28) | Cluster 2 (Positive) (N=8) | Cluster 1 (Negative) (N=10) | Cluster 2 (Positive) (N=8) | ||||||
| N | % | N | % | p | N | % | N | % | p | |
| Group | 0.002 | |||||||||
| Control | 18 | 64.3 | 0 | 0.0 | ||||||
| Schizophrenia | 10 | 35.7 | 8 | 100.0 | ||||||
| Mean | SD | Mean | SD | p | Mean | SD | Mean | SD | p | |
| Age (years) | 49.6 | 13.9 | 55.1 | 17.5 | 0.360 | 47.2 | 12.9 | 55.1 | 17.5 | 0.285 |
| Postmortem interval (hours) | 19.9 | 7.3 | 15.6 | 7.6 | 0.157 | 21.6 | 8.4 | 15.6 | 7.6 | 0.137 |
| RNA integrity number | 8.7 | 0.9 | 8.2 | 1.3 | 0.216 | 9.1 | 0.9 | 8.2 | 1.3 | 0.115 |
| Brain pH | 6.6 | 0.4 | 6.4 | 0.2 | 0.317 | 6.7 | 0.3 | 6.4 | 0.2 | 0.093 |
| Storage time (months) | 147.4 | 20.4 | 149.6 | 24.8 | 0.799 | 154.0 | 21.5 | 149.6 | 24.8 | 0.695 |
| N | % | N | % | p | N | % | N | % | p | |
| Sex | 0.107 | 0.145 | ||||||||
| Male | 20 | 71.4 | 3 | 38.0 | 8 | 80.0 | 3 | 37.5 | ||
| Female | 8 | 28.6 | 5 | 62.0 | 2 | 20.0 | 5 | 62.5 | ||
| Race | 0.063 | |||||||||
| White | 21 | 75.0 | 8 | 100.0 | 0.832 | 5 | 50.0 | 8 | 100.0 | |
| Black | 3 | 10.7 | 0 | 0.0 | 3 | 30.0 | 0 | 0.0 | ||
| Other | 4 | 14.3 | 0 | 0.0 | 2 | 20.0 | 0 | 0.0 | ||
| Medication use | ||||||||||
| Any | 9 | 90.0 | 6 | 75.0 | 0.559 | |||||
| Antipsychotic | 8 | 80.0 | 5 | 62.5 | 0.608 | |||||
| Anticonvulsant | 3 | 30.0 | 2 | 25.0 | 1.000 | |||||
| Antidepressant | 4 | 40.0 | 4 | 50.0 | 1.000 | |||||
| Benzodiazepine | 2 | 20.0 | 3 | 37.5 | 0.608 | |||||
| Other | 6 | 60.0 | 5 | 62.5 | 1.000 | |||||
a
Clusters were determined using hierarchical analysis.
Combined Data From the Three Studies
Since insolubility and ubiquitin levels for all three brain sets were normalized to total protein, the data from the pilot, full-scale, and replication studies could be directly combined (Figure 3). A hierarchical cluster analysis on the combined sample yielded a clustering pattern consistent with the individual brain sets alone (Figure 3A). In the combined sample, 20 schizophrenia brains had increased protein insolubility and ubiquitination compared with 22 schizophrenia brains and 41 control brains without these abnormalities. The same brains that clustered together in the individual brain sets clustered together in the combined sample, with the exception of brain S1, which switched to the positive cluster, and brain S33, which dropped out of the positive cluster. Brains in cluster 2 had significantly greater protein insolubility and ubiquitination than brains in cluster 1 (Figure 3B and 3C) (p<0.001 for both).
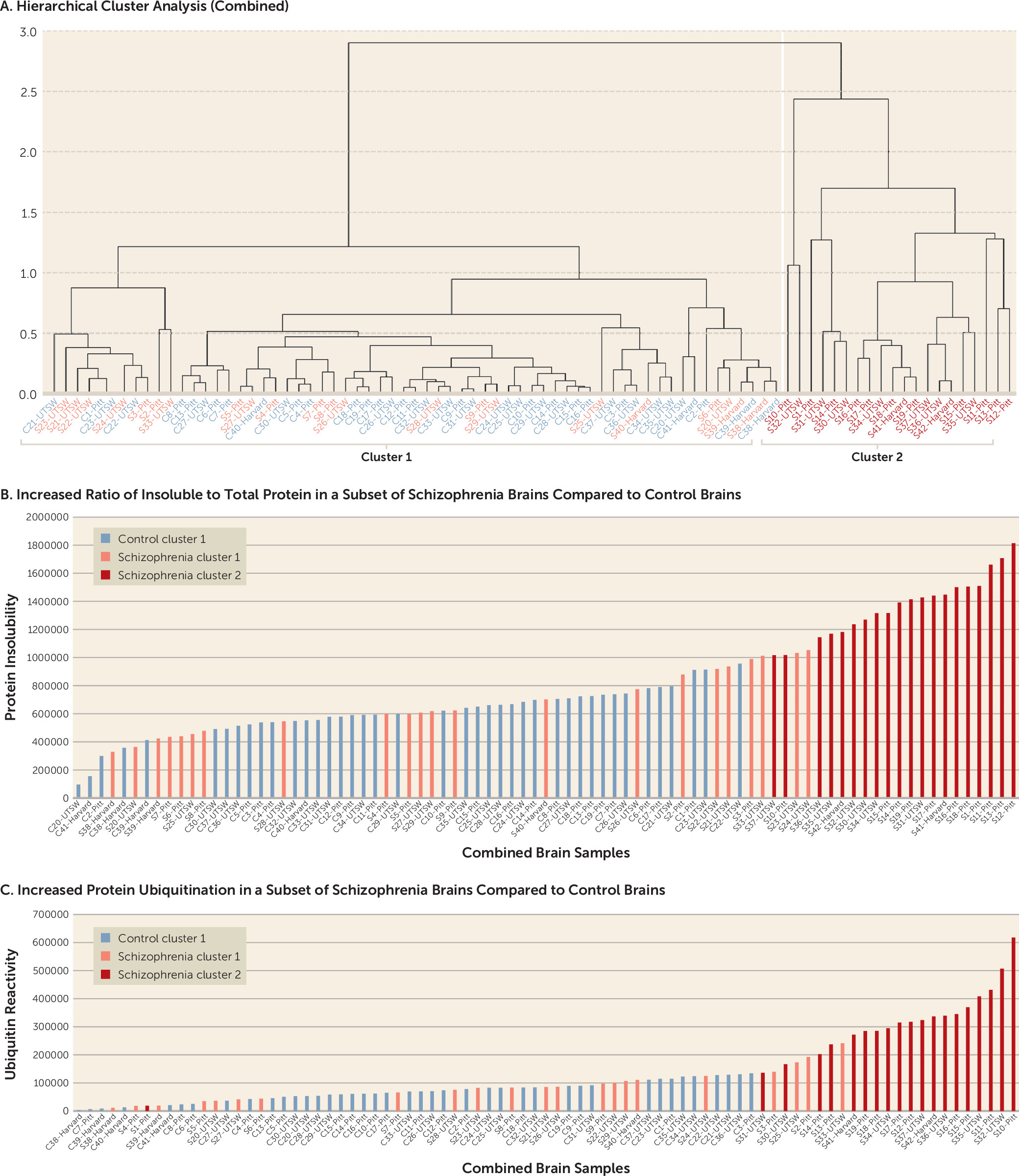
FIGURE 3. Increased protein insolubility and ubiquitination in a subset of schizophrenia brains compared with control brains in the combined sample of three brain banksa
a Data obtained from all 42 schizophrenia brains and 41 control brains were compared together. In panel A, hierarchical cluster analysis on all patient and control brains is similar to the separate analyses of the Harvard Brain Tissue Resource Center, University of Pittsburgh Brain Bank, and University of Texas Southwestern Medical Center samples. The same patient brains that clustered in the samples analyzed separately formed a larger cluster in the combined analysis, except that brain S1 became part of the positive cluster and brain S33 dropped out. The y-axis corresponds to dissimilarity. In panel B, protein insolubility was greater in brains from cluster 2 than in brains from cluster 1 (p>0.001). In panel C, ubiquitination in brains from cluster 2 was greater than in brains from cluster 1 (p<0.001).
Correlations Between Demographic Characteristics and Increased Protein Insolubility and Ubiquitination in the Three Studies Combined
While baseline demographic and other characteristics showed a significant difference only in the use of psychiatric medications between patients and control subjects (see Table S5 in the online supplement ), there was a significant difference in brain pH (p=0.002) (Table 3) between patient brains with increased protein insolubility and ubiquitination and brains of all other patients and control subjects in the combined sample. Furthermore, when comparing patient brains with increased markers to other patient brains, the former were likelier to be female (p=0.019) and to have lower brain pH (p=0.004).
| All Brains: Positive vs. Negative Cluster | Schizophrenia Brains: Positive vs. Negative Cluster | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Characteristic | Cluster 1 (Negative) (N=56, bN=63) | Cluster 2 (Positive) (N=18, bN=20) | Cluster 1 (Negative) (N=19, bN=22) | Cluster 2 (Positive) (N=18, bN=20) | ||||||
| N | % | N | % | p | N | % | N | % | p | |
| Groupb | <0.001 | |||||||||
| Control | 41 | 65.1 | 0 | 0.0 | ||||||
| Schizophrenia | 22 | 34.9 | 20 | 100.0 | ||||||
| Mean | SD | Mean | SD | p | Mean | SD | Mean | SD | p | |
| Ageb (years) | 48.6 | 12.1 | 52.4 | 14.5 | 0.240 | 48.3 | 11.6 | 52.4 | 14.5 | 0.312 |
| Postmortem intervalb (hours) | 19.5 | 7.1 | 17.1 | 7.3 | 0.194 | 20.6 | 7.5 | 17.1 | 7.3 | 0.134 |
| RNA integrity number | 8.5 | 0.7 | 8.1 | 1.0 | 0.069 | 8.7 | 0.9 | 8.1 | 1.0 | 0.064 |
| Brain pH | 6.6 | 0.3 | 6.4 | 0.3 | 0.002 | 6.7 | 0.3 | 6.4 | 0.3 | 0.004 |
| Storage time (months) | 148.8 | 34.8 | 148.3 | 25.8 | 0.952 | 149.0 | 35.7 | 148.3 | 25.8 | 0.941 |
| N | % | N | % | p | N | % | N | % | p | |
| Sexb | 0.053 | 0.019 | ||||||||
| Male | 47 | 74.6 | 10 | 50.0 | 19 | 86.4 | 10 | 50.0 | ||
| Female | 16 | 25.4 | 10 | 50.0 | 3 | 13.6 | 10 | 50.0 | ||
| Race | 0.099 | |||||||||
| White | 40 | 71.4 | 15 | 83.3 | 0.463 | 10 | 52.6 | 15 | 83.3 | |
| Black | 11 | 19.6 | 3 | 16.7 | 7 | 36.8 | 3 | 16.7 | ||
| Other | 5 | 9.0 | 0 | 0.0 | 2 | 10.6 | 0 | 0.0 | ||
| Medication use | ||||||||||
| Any | 17 | 89.5 | 16 | 88.9 | 1.000 | |||||
| Antipsychotic | 15 | 78.9 | 14 | 77.8 | 0.931 | |||||
| Anticonvulsant | 3 | 15.8 | 7 | 38.9 | 0.151 | |||||
| Antidepressant | 8 | 42.1 | 8 | 44.4 | 1.000 | |||||
| Benzodiazepine | 3 | 15.8 | 4 | 22.2 | 0.693 | |||||
| Other | 12 | 63.2 | 12 | 66.7 | 1.000 | |||||
a
Clusters were determined using hierarchical analysis.
b
Larger Ns were available for data that were available from all three data sets.
Effect of Antipsychotic Medication on Protein Insolubility and Ubiquitination in Rat Brain
An obvious distinction between patient and control brains is premortem exposure to antipsychotic medication. To determine whether chronic antipsychotic treatment itself can lead to the protein abnormalities detected in a subset of brains from individuals with schizophrenia, we treated rats for 4.5 months with haloperidol (1.5 mg/kg per day), risperidone (6 mg/kg per day), or water alone in their drinking water (dosages were based on previous work showing levels needed to produce vacuous chewing movements in rats). Using the same sarkosyl fractionation protocol applied to human brains, we found no evidence of medication-induced changes in protein insolubility or ubiquitination (see Figure S3 in the online supplement ), suggesting that the protein abnormalities detected in a subset of human brains were not a consequence of antipsychotic treatment.
Similar Molecular Signature of Insoluble Proteins in Patients With Increased Protein Insolubility and Ubiquitination
To investigate the potential impact of protein insolubility on cellular function, we performed mass spectrometry for the full-scale study to identify the specific proteins contained in the insoluble fraction from each of the brains in the University of Pittsburgh Brain Bank sample. A total of 182,913 peptide spectral matches were obtained in this analysis, corresponding to 6,958 unique peptides that were condensed to 994 unique proteins. Approximately seven unique peptides were found for each protein identified. (A complete list of the proteins and their quantification values is provided in Table S6 in the online supplement .) Of these, 602 proteins contained quantification measurements derived from at least two unique peptides for further analysis. A principal component analysis (Figure 4) demonstrated that the brains with high protein insolubility and ubiquitination separated from the group without these protein abnormalities. Specifically, brains S10, S12, S13, S14, S15, S16, S17, and S18 demonstrated tight clustering, and brains S11 and S19, while less clearly so, appeared to differentiate from the brains of most other patients and control subjects.
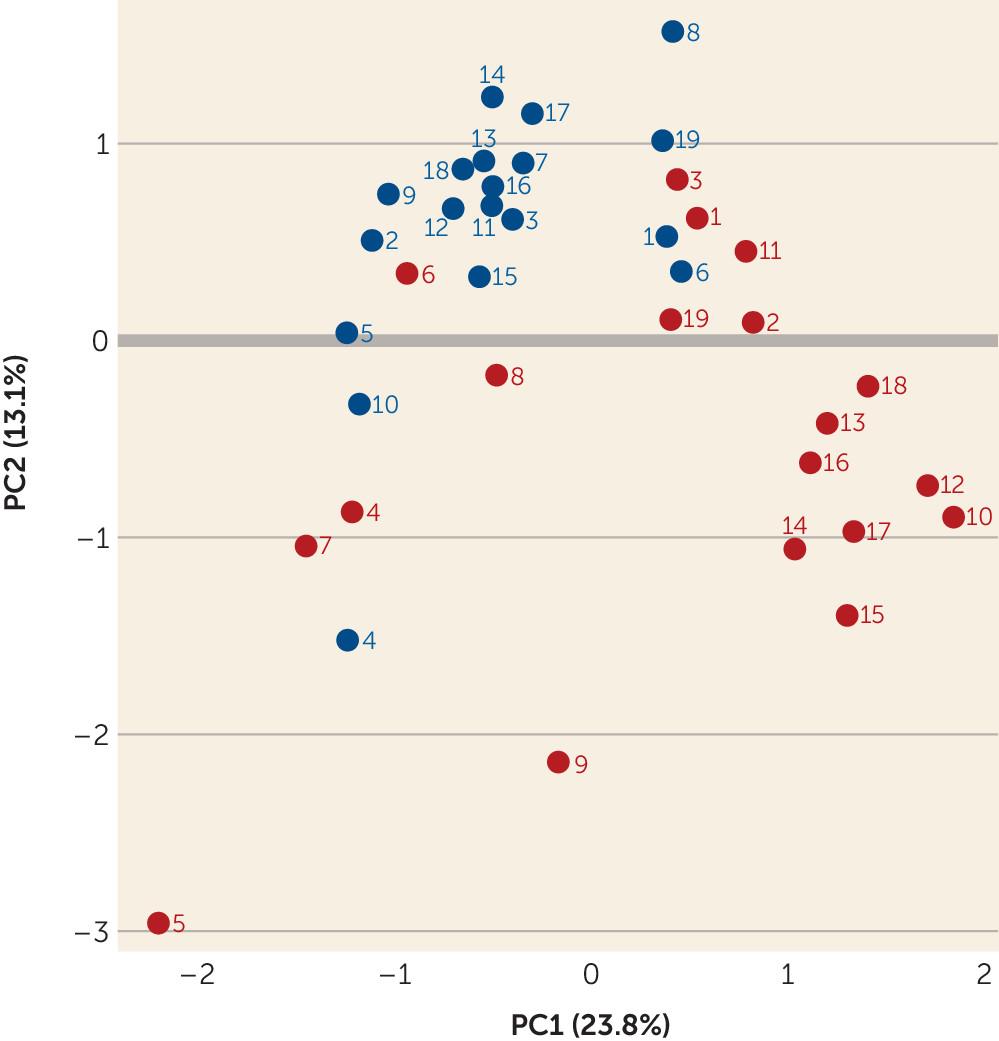
FIGURE 4. Similar molecular signature of insoluble proteins for schizophrenia brains with increased protein insolubility and ubiquitinationa
a A principal component analysis plot differentiates schizophrenia brains with high insolubility and high ubiquitination from patient and control brains without increases in these markers of protein abnormality.
Biological Processes and Pathways Identified From the Insoluble Proteins
We next used Gene Ontology (GO) enrichment analysis to analyze all of the proteins that were significantly different between the patients with high insolubility and ubiquitination compared with all other patients and control subjects to identify biological processes associated with the high protein insolubility phenotype. GO analysis identified axon target recognition as the biological process of highest significance (see Figure S4A in the online supplement ). We then performed Ingenuity Pathway Analysis (IPA), which provides expert manual curation of pathways and functions to enable the identification of statistical enrichment for a list of proteins in the curated pathways and functions. IPA identified neurological disease as a top pathway for diseases and disorders, cell assembly and organization as a top pathway for molecular and cellular function, and nervous system development and function as a top pathway for physiological system development and function (see Figure S4B in the online supplement ).
We also identified proteins enriched in several stages of the ubiquitination process in the insoluble fraction from brains with high insolubility compared with those with low insolubility, consistent with our detection of increased ubiquitination in the insoluble fraction (see Table S7 in the online supplement ).
Potential Cell Type Specificity
To determine whether the insoluble proteins are expressed by specific cell types, we utilized single cell RNA-seq expression data from brain cell type groups (from the Darmanis et al. data set) (31) and neuron subtype groups (from the Lake et al. data set) (32). We found that the insoluble proteins were enriched in neurons (false discovery rate <0.001) in the Darmanis et al. data set (see Figure S5A and Table S8 in the online supplement ). Analysis with the Lake et al. data set suggested that enrichment occurs in specific neuronal subtypes. Enrichment was significant (false discovery rate <0.05) for one inhibitory subtype (In6) and four excitatory subtypes (Ex1, Ex3, Ex4, and Ex5) (see Figure S5B and Table S8 in the online supplement ). From subtype characterization performed by Lake et al., In6 neurons are parvalbumin GABAergic neurons, Ex1 are cortical projection neurons in layers 2/3, Ex3 are granule neurons in layer 4, Ex4 are subcortical projection neurons in layers 4–6, and Ex5 are subcortical projection neurons in layers 5–6. We recognize that studying proteins through their RNA expression has its limitations. Further analysis should be performed when a brain cell type expression database is available for protein expression.
Discussion
In this study, we identified a subset of brains from schizophrenia patients with increased protein insolubility and increased protein ubiquitination in cortical tissue. When the insoluble fractions were analyzed by mass spectrometry, we found proteins that were present in all brains but found in greater abundances in the brains with high insolubility and ubiquitination. We also found unique proteins that were detected only in brains with high insolubility and ubiquitination. The insoluble proteins are disproportionally reflective of pathways related to nervous system development and axon target recognition, including axonogenesis, neurogenesis, and neuron projection.
Given the novelty of our findings, we used a tiered strategy of examining a small number of brains from the Harvard Brain Tissue Resource Center to explore our hypothesis, then retesting the hypothesis in a larger sample of brains from the University of Pittsburgh Brain Bank, and replicating the findings in a third sample of brains from University of Texas Southwestern Medical Center. Each sample yielded the same result, as did analysis of the combined samples, increasing our confidence that increased protein insolubility and ubiquitination are not artifacts related to the ascertainment, processing, or storage of postmortem brains in a particular brain bank. Furthermore, the available tissue from each brain bank was from a different cortical region, yet results were similar, consistent with the idea that aspects of schizophrenia pathophysiology have pan-cortical involvement (39–43). In addition, we used three different but related markers to explore protein abnormalities: insolubility, ubiquitin, and peptide abundance. As a result, we detected increased levels of these markers in a cluster of the same brains. We posit that this cluster identifies a subset of patients with a common pathophysiology.
Analysis of autopsy brains presents a number of challenges. Brain pH may reflect postmortem changes and storage conditions, and it could influence protein properties. We detected lower pH in the brains with protein abnormalities in the University of Pittsburgh Brain Bank sample and the combined sample from three brain banks, but not in the University of Texas Southwestern Medical Center sample analyzed alone. However, some of the brains with increased protein insolubility and ubiquitination were within the same pH range as brains from schizophrenia patients and control subjects without protein abnormalities. In addition, RIN values, which may be a better proxy for brain quality (13), did not differ significantly in any of our group comparisons. Interestingly, a difference of 0.2 to 0.4 pH units is frequently detected between schizophrenia and control brains (44–47). In addition, a magnetic resonance spectroscopy study showed that schizophrenia patients have significantly lower intracellular brain pH compared to control subjects (48). While the reason for this difference is not fully established, it may be related to premortem alterations in metabolic pathways and lactic acid (46, 49–53). In addition, rodent models indicate that oxidative stress can lower brain pH (54–56). Importantly, both processes are stress cascades implicated in schizophrenia; it is therefore possible that the processes leading to lower pH in schizophrenia brains is a primary intrinsic pathogenic process related to protein insolubility.
In general, alterations of protein conformation and solubility have been implicated as part of the primary pathogenic process (rather than a consequence of medication) of many disorders, including cancer, cardiac and pulmonary diseases, muscle diseases, and neurodegenerative disorders (6). However, antipsychotics and other medications influence various biological processes in the brain, potentially including protein processing. To address the concern that antipsychotics may influence protein solubility and ubiquitination, we chronically treated rats with either haloperidol or risperidone, representing two different classes of antipsychotics. Neither treatment led to protein insolubility or ubiquitination, suggesting that exposure to antipsychotics did not generate the protein abnormalities that we detected in human brains. However, this experiment was done without the extended PMI found in the human cases. In our data sets, there was no significant difference in exposure to antipsychotics, antidepressants, or benzodiazepines between brains with and without increased protein insolubility or ubiquitination. Exposure to anticonvulsants was significantly associated with increased protein insolubility and ubiquitination in the University of Pittsburgh Brain Bank samples, but not in the University of Texas Southwestern samples. However, there was no overall significant difference in anticonvulsant exposure when samples from all brain banks were combined. Anticonvulsants, and in particular valproic acid, may have neuroprotective properties in neurodegenerative disorders characterized by protein insolubility (57) and therefore likely do not promote protein misfolding and insolubility. Together, these data suggest that protein insolubility is intrinsic to the mechanism of a subset of mental illness and is not related to medication effects.
Since the brains in all three of our samples were obtained from coroner’s offices, there was limited availability of detailed social, medical, and psychiatric history to link abnormal protein solubility to a specific clinical phenotype, familial factor, or environmental exposure. However, there was a significant sex difference, with protein insolubility and ubiquitination more frequently found in brains from female patients in the combined sample. It is unknown whether this finding can be replicated in other brain collections and what the potential biological implications are. Furthermore, limitations in sample size precluded an analysis of the correlation of protein abnormalities with substance use or suicide; however, these phenomena were not prevalent in our samples and were nearly evenly distributed in brains with and without protein abnormalities. We suspect that cigarette smoking, whether through nicotine or other factors, is unlikely to contribute to individual variation in protein insolubility, as smoking was common to both patients with and without protein abnormalities, but we cannot exclude the relevance of variation in tobacco exposure with regard to protein abnormalities. Future analyses of brain samples with more clinical history would help clarify these issues.
Our findings are consistent with proposed pathogenic mechanisms of schizophrenia. While schizophrenia is not associated with cortical neuron loss (58), sequestration of proteins into insoluble forms could lead to disruption of critical pathways, resulting in neuronal dysfunction. Interestingly, while protein misfolding is often associated with neuronal death in neurodegenerative disorders, our pathway analysis implicated a neurodevelopmental role for protein insolubility in schizophrenia, consistent with the neurodevelopmental hypothesis of this disease (59–61). Enrichment of insoluble proteins in axon target recognition and neuronal projection pathways may be consistent with the related hypothesis that schizophrenia is a consequence of aberrant development of neuronal projections and connectivity (39, 62, 63).
A role for protein insolubility in schizophrenia has previously been suggested by studies of rare genetic variants associated with this disease (7, 8). DISC1, a protein originally associated with schizophrenia and affective disorders through a chromosomal translocation in a single large family, was identified in the insoluble fraction in a subset of brains from individuals with depression, bipolar affective disorder, and schizophrenia (64). Also, in a rat model, DISC1-containing protein aggregates disrupted dopamine homeostasis (65). A mutation in the neuronal transcription factor NPAS3, linked to schizophrenia in a small family (66), leads to the aggregation of NPAS3 and interferes with its transcriptional functioning (8). Other genetic factors associated with schizophrenia have been linked to similar protein abnormalities, including dysbindin-1, CRMP1, and TRIOBP-1 (67–70). Furthermore, environmental insults related to oxidative stress or altered immune signaling are linked with neuropsychiatric disorders and have also been associated with protein misfolding, such as endoplasmic reticulum stress and heat shock factors (71, 72).
Overall, our results provide strong support for the hypothesis that protein insolubility occurs in a subset of schizophrenia patients. Based on the function of the detected proteins, we suggest that the insolubility is related to disease pathogenesis. Future experiments could refine the regional and cell-type specificity of the phenomenon and elucidate the possible relationship between protein insolubility and other psychiatric disorders (e.g., bipolar disorder and major depressive disorder). Further exploration of the molecular mechanism underlying the protein insolubility in subsets of patients with schizophrenia or other disorders could lead to a better understanding of the pathways, circuitry, and symptoms seen in patients with mental illness, and could lead to improved nosology and novel therapeutic targets.
Footnotes
The content of this article is solely the responsibility of the authors and does not necessarily represent the official views of the U.S. Department of Veterans Affairs or the U.S. government.
Dr. MacDonald received support from NIMH grant K01 MH107756. Dr. Margolis received support from the ABCD Charitable Trust and grant support from Teva and NIH. Dr. Tamminga received support from Sunovion and travel funds from Autiphony Therapeutics, and has served as a consultant for Astellas, Eli Lilly, Kaye Scholer LLC for Pfizer, and Lundbeck. Dr. Sweet received support from NIMH grant R01 MH071533. Dr. Sawa received support from NIMH (grants MH-094268, MH-092443 Silvio O. Conte Center, MH-105660, and MH-107730), as well as support from the Stanley Foundation and the S&R Foundation.
Supplementary Material
File (appi.ajp.2019.18070864.ds001_tables_figures.pdf)
- View/Download
- 1.43 MB
File (appi.ajp.2019.18070864.ds002_tables6.xlsx)
- Download
- 279.81 KB
References
1.
Perälä J, Suvisaari J, Saarni SI, et al: Lifetime prevalence of psychotic and bipolar I disorders in a general population. Arch Gen Psychiatry 2007; 64:19–28
2.
Schizophrenia Working Group of the Psychiatric Genomics Consortium: Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014; 511:421–427
3.
Insel T, Cuthbert B, Garvey M, et al: Research Domain Criteria (RDoC): toward a new classification framework for research on mental disorders. Am J Psychiatry 2010; 167:748–751
4.
Clementz BA, Sweeney JA, Hamm JP, et al: Identification of distinct psychosis biotypes using brain-based biomarkers. Am J Psychiatry 2016; 173:373–384
5.
Arnedo J, Svrakic DM, Del Val C, et al: Uncovering the hidden risk architecture of the schizophrenias: confirmation in three independent genome-wide association studies. Am J Psychiatry 2015; 172:139–153
6.
Stefani M: Protein misfolding and aggregation: new examples in medicine and biology of the dark side of the protein world. Biochim Biophys Acta 2004; 1739:5–25
7.
Leliveld SR, Bader V, Hendriks P, et al: Insolubility of Disrupted-in-Schizophrenia 1 disrupts oligomer-dependent interactions with nuclear distribution element 1 and is associated with sporadic mental disease. J Neurosci 2008; 28:3839–3845
8.
Nucifora LG, Wu YC, Lee BJ, et al: A mutation in NPAS3 that segregates with schizophrenia in a small family leads to protein aggregation. Mol Neuropsychiatry 2016; 2:133–144
9.
Rubio MD, Wood K, Haroutunian V, et al: Dysfunction of the ubiquitin proteasome and ubiquitin-like systems in schizophrenia. Neuropsychopharmacology 2013; 38:1910–1920
10.
Scott MR, Rubio MD, Haroutunian V, et al: Protein expression of proteasome subunits in elderly patients with schizophrenia. Neuropsychopharmacology 2016; 41:896–905
11.
Nucifora FC Jr, Nucifora LG, Ng CH, et al: Ubiqutination via K27 and K29 chains signals aggregation and neuronal protection of LRRK2 by WSB1. Nat Commun 2016; 7:11792
12.
Deo AJ, Cahill ME, Li S, et al: Increased expression of Kalirin-9 in the auditory cortex of schizophrenia subjects: its role in dendritic pathology. Neurobiol Dis 2012; 45:796–803
13.
Stan AD, Ghose S, Gao XM, et al: Human postmortem tissue: what quality markers matter? Brain Res 2006; 1123:1–11
14.
Camchong J, Dyckman KA, Chapman CE, et al: Basal ganglia-thalamocortical circuitry disruptions in schizophrenia during delayed response tasks. Biol Psychiatry 2006; 60:235–241
15.
Glahn DC, Ragland JD, Abramoff A, et al: Beyond hypofrontality: a quantitative meta-analysis of functional neuroimaging studies of working memory in schizophrenia. Hum Brain Mapp 2005; 25:60–69
16.
Goldman-Rakic PS: Working memory dysfunction in schizophrenia. J Neuropsychiatry Clin Neurosci 1994; 6:348–357
17.
Javitt DC, Sweet RA: Auditory dysfunction in schizophrenia: integrating clinical and basic features. Nat Rev Neurosci 2015; 16:535–550
18.
Manoach DS, Press DZ, Thangaraj V, et al: Schizophrenic subjects activate dorsolateral prefrontal cortex during a working memory task, as measured by fMRI. Biol Psychiatry 1999; 45:1128–1137
19.
Rajarethinam RP, DeQuardo JR, Nalepa R, et al: Superior temporal gyrus in schizophrenia: a volumetric magnetic resonance imaging study. Schizophr Res 2000; 41:303–312
20.
Seidman LJ, Thermenos HW, Poldrack RA, et al: Altered brain activation in dorsolateral prefrontal cortex in adolescents and young adults at genetic risk for schizophrenia: an fMRI study of working memory. Schizophr Res 2006; 85:58–72
21.
Sun J, Maller JJ, Guo L, et al: Superior temporal gyrus volume change in schizophrenia: a review on region of interest volumetric studies. Brain Res Brain Res Rev 2009; 61:14–32
22.
Weinberger DR: Schizophrenia and the frontal lobe. Trends Neurosci 1988; 11:367–370
23.
Wojtalik JA, Smith MJ, Keshavan MS, et al: A systematic and meta-analytic review of neural correlates of functional outcome in schizophrenia. Schizophr Bull 2017; 43:1329–1347
24.
Gao XM, Cooper T, Suckow RF, et al: Multidose risperidone treatment evaluated in a rodent model of tardive dyskinesia. Neuropsychopharmacology 2006; 31:1864–1868
25.
Erde J, Loo RR, Loo JA: Enhanced FASP (eFASP) to increase proteome coverage and sample recovery for quantitative proteomic experiments. J Proteome Res 2014; 13:1885–1895
26.
Paweletz CP, Wiener MC, Bondarenko AY, et al: Application of an end-to-end biomarker discovery platform to identify target engagement markers in cerebrospinal fluid by high resolution differential mass spectrometry. J Proteome Res 2010; 9:1392–1401
27.
Shen X, Shen S, Li J, et al: An IonStar experimental strategy for MS1 ion current-based quantification using ultrahigh-field orbitrap: reproducible, in-depth, and accurate protein measurement in large cohorts. J Proteome Res 2017; 16:2445–2456
28.
The M, Tasnim A, Käll L: How to talk about protein-level false discovery rates in shotgun proteomics. Proteomics 2016; 16:2461–2469
29.
Käll L, Canterbury JD, Weston J, et al: Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat Methods 2007; 4:923–925
30.
Oberg AL, Vitek O: Statistical design of quantitative mass spectrometry-based proteomic experiments. J Proteome Res 2009; 8:2144–2156
31.
Darmanis S, Sloan SA, Zhang Y, et al: A survey of human brain transcriptome diversity at the single cell level. Proc Natl Acad Sci USA 2015; 112:7285–7290
32.
Lake BB, Ai R, Kaeser GE, et al: Neuronal subtypes and diversity revealed by single-nucleus RNA sequencing of the human brain. Science 2016; 352:1586–1590
33.
Diner I, Hales CM, Bishof I, et al: Aggregation properties of the small nuclear ribonucleoprotein U1-70K in Alzheimer disease. J Biol Chem 2014; 289:35296–35313
34.
Diner I, Nguyen T, Seyfried NT: Enrichment of detergent-insoluble protein aggregates from human postmortem brain. J Vis Exp 2017 (doi: 10.3791/55835)
35.
Ferrer I, Santpere G, Arzberger T, et al: Brain protein preservation largely depends on the postmortem storage temperature: implications for study of proteins in human neurologic diseases and management of brain banks: a BrainNet Europe study. J Neuropathol Exp Neurol 2007; 66:35–46
36.
Halliday GM, Song YJ, Lepar G, et al: Pick bodies in a family with presenilin-1 Alzheimer’s disease. Ann Neurol 2005; 57:139–143
37.
Ikeuchi T, Kakita A, Shiga A, et al: Patients homozygous and heterozygous for SNCA duplication in a family with parkinsonism and dementia. Arch Neurol 2008; 65:514–519
38.
Tremblay C, François A, Delay C, et al: Association of neuropathological markers in the parietal cortex with antemortem cognitive function in persons with mild cognitive impairment and Alzheimer disease. J Neuropathol Exp Neurol (Epub ahead of print, January 31, 2017)
39.
Owen MJ, Sawa A, Mortensen PB: Schizophrenia. Lancet 2016; 388:86–97
40.
Lewis DA: Cortical circuit dysfunction and cognitive deficits in schizophrenia: implications for preemptive interventions. Eur J Neurosci 2012; 35:1871–1878
41.
Linden DE: The challenges and promise of neuroimaging in psychiatry. Neuron 2012; 73:8–22
42.
Haijma SV, Van Haren N, Cahn W, et al: Brain volumes in schizophrenia: a meta-analysis in over 18 000 subjects. Schizophr Bull 2013; 39:1129–1138
43.
Wheeler AL, Voineskos AN: A review of structural neuroimaging in schizophrenia: from connectivity to connectomics. Front Hum Neurosci 2014; 8:653
44.
Eastwood SL, Harrison PJ: Decreased expression of vesicular glutamate transporter 1 and complexin II mRNAs in schizophrenia: further evidence for a synaptic pathology affecting glutamate neurons. Schizophr Res 2005; 73:159–172
45.
Lipska BK, Deep-Soboslay A, Weickert CS, et al: Critical factors in gene expression in postmortem human brain: focus on studies in schizophrenia. Biol Psychiatry 2006; 60:650–658
46.
Prabakaran S, Swatton JE, Ryan MM, et al: Mitochondrial dysfunction in schizophrenia: evidence for compromised brain metabolism and oxidative stress. Mol Psychiatry 2004; 9:684–697
47.
Torrey EF, Barci BM, Webster MJ, et al: Neurochemical markers for schizophrenia, bipolar disorder, and major depression in postmortem brains. Biol Psychiatry 2005; 57:252–260
48.
Du F, Cooper AJ, Thida T, et al: In vivo evidence for cerebral bioenergetic abnormalities in schizophrenia measured using 31P magnetization transfer spectroscopy. JAMA Psychiatry 2014; 71:19–27
49.
Holmes E, Tsang TM, Huang JT, et al: Metabolic profiling of CSF: evidence that early intervention may impact on disease progression and outcome in schizophrenia. PLoS Med 2006; 3:e327
50.
Huang JT, Leweke FM, Tsang TM, et al: CSF metabolic and proteomic profiles in patients prodromal for psychosis. PLoS One 2007; 2:e756
51.
Altar CA, Jurata LW, Charles V, et al: Deficient hippocampal neuron expression of proteasome, ubiquitin, and mitochondrial genes in multiple schizophrenia cohorts. Biol Psychiatry 2005; 58:85–96
52.
Prabakaran S, Wengenroth M, Lockstone HE, et al: 2-D DIGE analysis of liver and red blood cells provides further evidence for oxidative stress in schizophrenia. J Proteome Res 2007; 6:141–149
53.
Michaelis T, Helms G, Frahm J: Metabolic alterations in brain autopsies: proton NMR identification of free glycerol. NMR Biomed 1996; 9:121–124
54.
Sipos H, Törocsik B, Tretter L, et al: Impaired regulation of pH homeostasis by oxidative stress in rat brain capillary endothelial cells. Cell Mol Neurobiol 2005; 25:141–151
55.
Tsai KL, Wang SM, Chen CC, et al: Mechanism of oxidative stress-induced intracellular acidosis in rat cerebellar astrocytes and C6 glioma cells. J Physiol 1997; 502:161–174
56.
Mulkey DK, Henderson RA 3rd, Ritucci NA, et al: Oxidative stress decreases pHi and Na(+)/H(+) exchange and increases excitability of solitary complex neurons from rat brain slices. Am J Physiol Cell Physiol 2004; 286:C940–C951
57.
Chiu CT, Wang Z, Hunsberger JG, et al: Therapeutic potential of mood stabilizers lithium and valproic acid: beyond bipolar disorder. Pharmacol Rev 2013; 65:105–142
58.
Dorph-Petersen KA, Lewis DA: Postmortem structural studies of the thalamus in schizophrenia. Schizophr Res 2017; 180:28–35
59.
Weinberger DR: Implications of normal brain development for the pathogenesis of schizophrenia. Arch Gen Psychiatry 1987; 44:660–669
60.
Birnbaum R, Weinberger DR: Genetic insights into the neurodevelopmental origins of schizophrenia. Nat Rev Neurosci 2017; 18:727–740
61.
Ursini G, Punzi G, Chen Q, et al: Convergence of placenta biology and genetic risk for schizophrenia. Nat Med 2018; 24:792–801
62.
Garrity AG, Pearlson GD, McKiernan K, et al: Aberrant “default mode” functional connectivity in schizophrenia. Am J Psychiatry 2007; 164:450–457
63.
Skudlarski P, Jagannathan K, Anderson K, et al: Brain connectivity is not only lower but different in schizophrenia: a combined anatomical and functional approach. Biol Psychiatry 2010; 68:61–69
64.
St Clair D, Blackwood D, Muir W, et al: Association within a family of a balanced autosomal translocation with major mental illness. Lancet 1990; 336:13–16
65.
Trossbach SV, Bader V, Hecher L, et al: Misassembly of full-length Disrupted-in-Schizophrenia 1 protein is linked to altered dopamine homeostasis and behavioral deficits. Mol Psychiatry 2016; 21:1561–1572
66.
Yu L, Arbez N, Nucifora LG, et al: A mutation in NPAS3 segregates with mental illness in a small family. Mol Psychiatry 2014; 19:7–8
67.
Bradshaw NJ, Korth C: Protein misassembly and aggregation as potential convergence points for non-genetic causes of chronic mental illness. Mol Psychiatry (Epub ahead of print, August 8, 2018)
68.
Ottis P, Bader V, Trossbach SV, et al: Convergence of two independent mental disease genes on the protein level: recruitment of dysbindin to cell-invasive disrupted-in-schizophrenia 1 aggresomes. Biol Psychiatry 2011; 70:604–610
69.
Bader V, Tomppo L, Trossbach SV, et al: Proteomic, genomic, and translational approaches identify CRMP1 for a role in schizophrenia and its underlying traits. Hum Mol Genet 2012; 21:4406–4418
70.
Bradshaw NJ, Bader V, Prikulis I, et al: Aggregation of the protein TRIOBP-1 and its potential relevance to schizophrenia. PLoS One 2014; 9:e111196
71.
Saxena S, Caroni P: Selective neuronal vulnerability in neurodegenerative diseases: from stressor thresholds to degeneration. Neuron 2011; 71:35–48
72.
Akerfelt M, Morimoto RI, Sistonen L: Heat shock factors: integrators of cell stress, development and lifespan. Nat Rev Mol Cell Biol 2010; 11:545–555
Information & Authors
Information
Published In


History
Received: 23 July 2018
Revision received: 4 December 2018
Revision received: 6 January 2019
Accepted: 19 February 2019
Published online: 6 May 2019
Published in print: September 01, 2019
Keywords
Authors
Funding Information
ABCD Charitable Trust:
National Institute of Mental Health10.13039/100000025: K01 MH 107756, MH-094268, MH-092443 Silvio O. Conte center, MH-10, R01, MH071533
Brain and Behavior Research Foundation10.13039/100000874: Young Investigator Award
Supported by the Brain and Behavior Research Foundation (Young Investigator Award 2017 to Dr. L.G. Nucifora; Young Investigator Award 2013 to Dr. F.C. Nucifora).Metrics & Citations
Metrics
Citations
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
For more information or tips please see 'Downloading to a citation manager' in the Help menu.
View Options
View options
PDF/EPUB
View PDF/EPUBFigures
FIGURE 1. Increased ratio of insoluble protein and ubiquitination to total protein in a subset of schizophrenia brains compared with control brains from the full-scale study (University of Pittsburgh Brain Bank)a
a Cold sarkosyl fractionation was used to obtain insoluble fractions from postmortem brains of 19 schizophrenia patients and 19 control subjects from the University of Pittsburgh Brain Bank. In panel A, when insolubility and ubiquitination measurements were jointly analyzed, a subset of patient brains separated from other patient and control brains. In panel B, hierarchical clustering analysis determined that 10 of the schizophrenia brains (brains S10–S19, cluster 2) clustered separately from all other patient and control brains. The y-axis corresponds to dissimilarity. In panel C, schizophrenia brains in cluster 2 have significantly more protein insolubility than the patient and control brains in cluster 1 (p≤0.001). In panel D, the schizophrenia brains in cluster 2 were also significantly different from the patient and control brains in cluster 1 based on the amount of ubiquitination in the insoluble fraction (p≤0.001). Blue represents control brains with low levels of the markers of protein abnormality (insolubility and ubiquitination), pink indicates schizophrenia brains with low levels of these markers, and red indicates schizophrenia brains with high levels of these markers, based on hierarchical cluster analysis. Brain S1 was excluded from the positive cluster by hierarchical cluster analysis because it had a high level of protein insolubility but a low level of ubiquitination. In panel E, the ubiquitination was strikingly higher in the subset of schizophrenia brains with increased markers (cluster 2) than in the brains of the other patients or control subjects (cluster 1).
FIGURE 2. Increased ratio of insoluble protein and ubiquitination to total protein in a subset of schizophrenia brains compared with control brains from the replication study (University of Texas Southwestern Medical Center)a
a Cold sarkosyl fractionation was used to obtain insoluble fractions from postmortem brains of 18 schizophrenia patients and 18 control subjects from the University of Texas Southwestern Medical Center (UTSW). In panel A, when insolubility and ubiquitination measurements were jointly analyzed, a subset of schizophrenia brains separated from all other patient and control brains. In panel B, hierarchical cluster analysis determined that eight of the schizophrenia brains (brains S30–S37, cluster 2) clustered separately from all other patient and control brains. The y-axis corresponds to dissimilarity. In panel C, schizophrenia brains in cluster 2 have significantly more protein insolubility than the patient and control brains in cluster 1 (p<0.001). In panel D, the schizophrenia brains in cluster 2 were also significantly different than the patient and control brains in cluster 1 based on the amount of ubiquitination in the insoluble fraction (p<0.001). Blue represents control brains with low levels of the markers of protein abnormality (insolubility and ubiquitination), pink indicates schizophrenia brains with low levels of these markers, and red indicates schizophrenia brains with high levels of these markers based on hierarchical clustering analysis. Hierarchical clustering analysis excluded brains S23 and S24 because they had high levels of protein insolubility but low levels of ubiquitination, and brain S25 because it had a high level of ubiquitination but a low level of protein insolubility. In panel E, the ubiquitination was strikingly higher in the subset of schizophrenia brains with increased markers (cluster 2) than in the brains of the other patients or control subjects (cluster 1).
FIGURE 3. Increased protein insolubility and ubiquitination in a subset of schizophrenia brains compared with control brains in the combined sample of three brain banksa
a Data obtained from all 42 schizophrenia brains and 41 control brains were compared together. In panel A, hierarchical cluster analysis on all patient and control brains is similar to the separate analyses of the Harvard Brain Tissue Resource Center, University of Pittsburgh Brain Bank, and University of Texas Southwestern Medical Center samples. The same patient brains that clustered in the samples analyzed separately formed a larger cluster in the combined analysis, except that brain S1 became part of the positive cluster and brain S33 dropped out. The y-axis corresponds to dissimilarity. In panel B, protein insolubility was greater in brains from cluster 2 than in brains from cluster 1 (p>0.001). In panel C, ubiquitination in brains from cluster 2 was greater than in brains from cluster 1 (p<0.001).
FIGURE 4. Similar molecular signature of insoluble proteins for schizophrenia brains with increased protein insolubility and ubiquitinationa
a A principal component analysis plot differentiates schizophrenia brains with high insolubility and high ubiquitination from patient and control brains without increases in these markers of protein abnormality.
Tables
Media
References
References
1.
Perälä J, Suvisaari J, Saarni SI, et al: Lifetime prevalence of psychotic and bipolar I disorders in a general population. Arch Gen Psychiatry 2007; 64:19–28
2.
Schizophrenia Working Group of the Psychiatric Genomics Consortium: Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014; 511:421–427
3.
Insel T, Cuthbert B, Garvey M, et al: Research Domain Criteria (RDoC): toward a new classification framework for research on mental disorders. Am J Psychiatry 2010; 167:748–751
4.
Clementz BA, Sweeney JA, Hamm JP, et al: Identification of distinct psychosis biotypes using brain-based biomarkers. Am J Psychiatry 2016; 173:373–384
5.
Arnedo J, Svrakic DM, Del Val C, et al: Uncovering the hidden risk architecture of the schizophrenias: confirmation in three independent genome-wide association studies. Am J Psychiatry 2015; 172:139–153
6.
Stefani M: Protein misfolding and aggregation: new examples in medicine and biology of the dark side of the protein world. Biochim Biophys Acta 2004; 1739:5–25
7.
Leliveld SR, Bader V, Hendriks P, et al: Insolubility of Disrupted-in-Schizophrenia 1 disrupts oligomer-dependent interactions with nuclear distribution element 1 and is associated with sporadic mental disease. J Neurosci 2008; 28:3839–3845
8.
Nucifora LG, Wu YC, Lee BJ, et al: A mutation in NPAS3 that segregates with schizophrenia in a small family leads to protein aggregation. Mol Neuropsychiatry 2016; 2:133–144
9.
Rubio MD, Wood K, Haroutunian V, et al: Dysfunction of the ubiquitin proteasome and ubiquitin-like systems in schizophrenia. Neuropsychopharmacology 2013; 38:1910–1920
10.
Scott MR, Rubio MD, Haroutunian V, et al: Protein expression of proteasome subunits in elderly patients with schizophrenia. Neuropsychopharmacology 2016; 41:896–905
11.
Nucifora FC Jr, Nucifora LG, Ng CH, et al: Ubiqutination via K27 and K29 chains signals aggregation and neuronal protection of LRRK2 by WSB1. Nat Commun 2016; 7:11792
12.
Deo AJ, Cahill ME, Li S, et al: Increased expression of Kalirin-9 in the auditory cortex of schizophrenia subjects: its role in dendritic pathology. Neurobiol Dis 2012; 45:796–803
13.
Stan AD, Ghose S, Gao XM, et al: Human postmortem tissue: what quality markers matter? Brain Res 2006; 1123:1–11
14.
Camchong J, Dyckman KA, Chapman CE, et al: Basal ganglia-thalamocortical circuitry disruptions in schizophrenia during delayed response tasks. Biol Psychiatry 2006; 60:235–241
15.
Glahn DC, Ragland JD, Abramoff A, et al: Beyond hypofrontality: a quantitative meta-analysis of functional neuroimaging studies of working memory in schizophrenia. Hum Brain Mapp 2005; 25:60–69
16.
Goldman-Rakic PS: Working memory dysfunction in schizophrenia. J Neuropsychiatry Clin Neurosci 1994; 6:348–357
17.
Javitt DC, Sweet RA: Auditory dysfunction in schizophrenia: integrating clinical and basic features. Nat Rev Neurosci 2015; 16:535–550
18.
Manoach DS, Press DZ, Thangaraj V, et al: Schizophrenic subjects activate dorsolateral prefrontal cortex during a working memory task, as measured by fMRI. Biol Psychiatry 1999; 45:1128–1137
19.
Rajarethinam RP, DeQuardo JR, Nalepa R, et al: Superior temporal gyrus in schizophrenia: a volumetric magnetic resonance imaging study. Schizophr Res 2000; 41:303–312
20.
Seidman LJ, Thermenos HW, Poldrack RA, et al: Altered brain activation in dorsolateral prefrontal cortex in adolescents and young adults at genetic risk for schizophrenia: an fMRI study of working memory. Schizophr Res 2006; 85:58–72
21.
Sun J, Maller JJ, Guo L, et al: Superior temporal gyrus volume change in schizophrenia: a review on region of interest volumetric studies. Brain Res Brain Res Rev 2009; 61:14–32
22.
Weinberger DR: Schizophrenia and the frontal lobe. Trends Neurosci 1988; 11:367–370
23.
Wojtalik JA, Smith MJ, Keshavan MS, et al: A systematic and meta-analytic review of neural correlates of functional outcome in schizophrenia. Schizophr Bull 2017; 43:1329–1347
24.
Gao XM, Cooper T, Suckow RF, et al: Multidose risperidone treatment evaluated in a rodent model of tardive dyskinesia. Neuropsychopharmacology 2006; 31:1864–1868
25.
Erde J, Loo RR, Loo JA: Enhanced FASP (eFASP) to increase proteome coverage and sample recovery for quantitative proteomic experiments. J Proteome Res 2014; 13:1885–1895
26.
Paweletz CP, Wiener MC, Bondarenko AY, et al: Application of an end-to-end biomarker discovery platform to identify target engagement markers in cerebrospinal fluid by high resolution differential mass spectrometry. J Proteome Res 2010; 9:1392–1401
27.
Shen X, Shen S, Li J, et al: An IonStar experimental strategy for MS1 ion current-based quantification using ultrahigh-field orbitrap: reproducible, in-depth, and accurate protein measurement in large cohorts. J Proteome Res 2017; 16:2445–2456
28.
The M, Tasnim A, Käll L: How to talk about protein-level false discovery rates in shotgun proteomics. Proteomics 2016; 16:2461–2469
29.
Käll L, Canterbury JD, Weston J, et al: Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat Methods 2007; 4:923–925
30.
Oberg AL, Vitek O: Statistical design of quantitative mass spectrometry-based proteomic experiments. J Proteome Res 2009; 8:2144–2156
31.
Darmanis S, Sloan SA, Zhang Y, et al: A survey of human brain transcriptome diversity at the single cell level. Proc Natl Acad Sci USA 2015; 112:7285–7290
32.
Lake BB, Ai R, Kaeser GE, et al: Neuronal subtypes and diversity revealed by single-nucleus RNA sequencing of the human brain. Science 2016; 352:1586–1590
33.
Diner I, Hales CM, Bishof I, et al: Aggregation properties of the small nuclear ribonucleoprotein U1-70K in Alzheimer disease. J Biol Chem 2014; 289:35296–35313
34.
Diner I, Nguyen T, Seyfried NT: Enrichment of detergent-insoluble protein aggregates from human postmortem brain. J Vis Exp 2017 (doi: 10.3791/55835)
35.
Ferrer I, Santpere G, Arzberger T, et al: Brain protein preservation largely depends on the postmortem storage temperature: implications for study of proteins in human neurologic diseases and management of brain banks: a BrainNet Europe study. J Neuropathol Exp Neurol 2007; 66:35–46
36.
Halliday GM, Song YJ, Lepar G, et al: Pick bodies in a family with presenilin-1 Alzheimer’s disease. Ann Neurol 2005; 57:139–143
37.
Ikeuchi T, Kakita A, Shiga A, et al: Patients homozygous and heterozygous for SNCA duplication in a family with parkinsonism and dementia. Arch Neurol 2008; 65:514–519
38.
Tremblay C, François A, Delay C, et al: Association of neuropathological markers in the parietal cortex with antemortem cognitive function in persons with mild cognitive impairment and Alzheimer disease. J Neuropathol Exp Neurol (Epub ahead of print, January 31, 2017)
39.
Owen MJ, Sawa A, Mortensen PB: Schizophrenia. Lancet 2016; 388:86–97
40.
Lewis DA: Cortical circuit dysfunction and cognitive deficits in schizophrenia: implications for preemptive interventions. Eur J Neurosci 2012; 35:1871–1878
41.
Linden DE: The challenges and promise of neuroimaging in psychiatry. Neuron 2012; 73:8–22
42.
Haijma SV, Van Haren N, Cahn W, et al: Brain volumes in schizophrenia: a meta-analysis in over 18 000 subjects. Schizophr Bull 2013; 39:1129–1138
43.
Wheeler AL, Voineskos AN: A review of structural neuroimaging in schizophrenia: from connectivity to connectomics. Front Hum Neurosci 2014; 8:653
44.
Eastwood SL, Harrison PJ: Decreased expression of vesicular glutamate transporter 1 and complexin II mRNAs in schizophrenia: further evidence for a synaptic pathology affecting glutamate neurons. Schizophr Res 2005; 73:159–172
45.
Lipska BK, Deep-Soboslay A, Weickert CS, et al: Critical factors in gene expression in postmortem human brain: focus on studies in schizophrenia. Biol Psychiatry 2006; 60:650–658
46.
Prabakaran S, Swatton JE, Ryan MM, et al: Mitochondrial dysfunction in schizophrenia: evidence for compromised brain metabolism and oxidative stress. Mol Psychiatry 2004; 9:684–697
47.
Torrey EF, Barci BM, Webster MJ, et al: Neurochemical markers for schizophrenia, bipolar disorder, and major depression in postmortem brains. Biol Psychiatry 2005; 57:252–260
48.
Du F, Cooper AJ, Thida T, et al: In vivo evidence for cerebral bioenergetic abnormalities in schizophrenia measured using 31P magnetization transfer spectroscopy. JAMA Psychiatry 2014; 71:19–27
49.
Holmes E, Tsang TM, Huang JT, et al: Metabolic profiling of CSF: evidence that early intervention may impact on disease progression and outcome in schizophrenia. PLoS Med 2006; 3:e327
50.
Huang JT, Leweke FM, Tsang TM, et al: CSF metabolic and proteomic profiles in patients prodromal for psychosis. PLoS One 2007; 2:e756
51.
Altar CA, Jurata LW, Charles V, et al: Deficient hippocampal neuron expression of proteasome, ubiquitin, and mitochondrial genes in multiple schizophrenia cohorts. Biol Psychiatry 2005; 58:85–96
52.
Prabakaran S, Wengenroth M, Lockstone HE, et al: 2-D DIGE analysis of liver and red blood cells provides further evidence for oxidative stress in schizophrenia. J Proteome Res 2007; 6:141–149
53.
Michaelis T, Helms G, Frahm J: Metabolic alterations in brain autopsies: proton NMR identification of free glycerol. NMR Biomed 1996; 9:121–124
54.
Sipos H, Törocsik B, Tretter L, et al: Impaired regulation of pH homeostasis by oxidative stress in rat brain capillary endothelial cells. Cell Mol Neurobiol 2005; 25:141–151
55.
Tsai KL, Wang SM, Chen CC, et al: Mechanism of oxidative stress-induced intracellular acidosis in rat cerebellar astrocytes and C6 glioma cells. J Physiol 1997; 502:161–174
56.
Mulkey DK, Henderson RA 3rd, Ritucci NA, et al: Oxidative stress decreases pHi and Na(+)/H(+) exchange and increases excitability of solitary complex neurons from rat brain slices. Am J Physiol Cell Physiol 2004; 286:C940–C951
57.
Chiu CT, Wang Z, Hunsberger JG, et al: Therapeutic potential of mood stabilizers lithium and valproic acid: beyond bipolar disorder. Pharmacol Rev 2013; 65:105–142
58.
Dorph-Petersen KA, Lewis DA: Postmortem structural studies of the thalamus in schizophrenia. Schizophr Res 2017; 180:28–35
59.
Weinberger DR: Implications of normal brain development for the pathogenesis of schizophrenia. Arch Gen Psychiatry 1987; 44:660–669
60.
Birnbaum R, Weinberger DR: Genetic insights into the neurodevelopmental origins of schizophrenia. Nat Rev Neurosci 2017; 18:727–740
61.
Ursini G, Punzi G, Chen Q, et al: Convergence of placenta biology and genetic risk for schizophrenia. Nat Med 2018; 24:792–801
62.
Garrity AG, Pearlson GD, McKiernan K, et al: Aberrant “default mode” functional connectivity in schizophrenia. Am J Psychiatry 2007; 164:450–457
63.
Skudlarski P, Jagannathan K, Anderson K, et al: Brain connectivity is not only lower but different in schizophrenia: a combined anatomical and functional approach. Biol Psychiatry 2010; 68:61–69
64.
St Clair D, Blackwood D, Muir W, et al: Association within a family of a balanced autosomal translocation with major mental illness. Lancet 1990; 336:13–16
65.
Trossbach SV, Bader V, Hecher L, et al: Misassembly of full-length Disrupted-in-Schizophrenia 1 protein is linked to altered dopamine homeostasis and behavioral deficits. Mol Psychiatry 2016; 21:1561–1572
66.
Yu L, Arbez N, Nucifora LG, et al: A mutation in NPAS3 segregates with mental illness in a small family. Mol Psychiatry 2014; 19:7–8
67.
Bradshaw NJ, Korth C: Protein misassembly and aggregation as potential convergence points for non-genetic causes of chronic mental illness. Mol Psychiatry (Epub ahead of print, August 8, 2018)
68.
Ottis P, Bader V, Trossbach SV, et al: Convergence of two independent mental disease genes on the protein level: recruitment of dysbindin to cell-invasive disrupted-in-schizophrenia 1 aggresomes. Biol Psychiatry 2011; 70:604–610
69.
Bader V, Tomppo L, Trossbach SV, et al: Proteomic, genomic, and translational approaches identify CRMP1 for a role in schizophrenia and its underlying traits. Hum Mol Genet 2012; 21:4406–4418
70.
Bradshaw NJ, Bader V, Prikulis I, et al: Aggregation of the protein TRIOBP-1 and its potential relevance to schizophrenia. PLoS One 2014; 9:e111196
71.
Saxena S, Caroni P: Selective neuronal vulnerability in neurodegenerative diseases: from stressor thresholds to degeneration. Neuron 2011; 71:35–48
72.
Akerfelt M, Morimoto RI, Sistonen L: Heat shock factors: integrators of cell stress, development and lifespan. Nat Rev Mol Cell Biol 2010; 11:545–555