Nothing Tastes as Good as Skinny Feels: the Neurobiology of Anorexia Nervosa
Abstract
Introduction
Box 1. Example of a typical AN case
Temperament and personality in AN
Anxiety
Inhibitory self-control and reward
Set shifting
Harm avoidance
Perfectionism
Interoception and alexithymia
Appetite regulation in AN
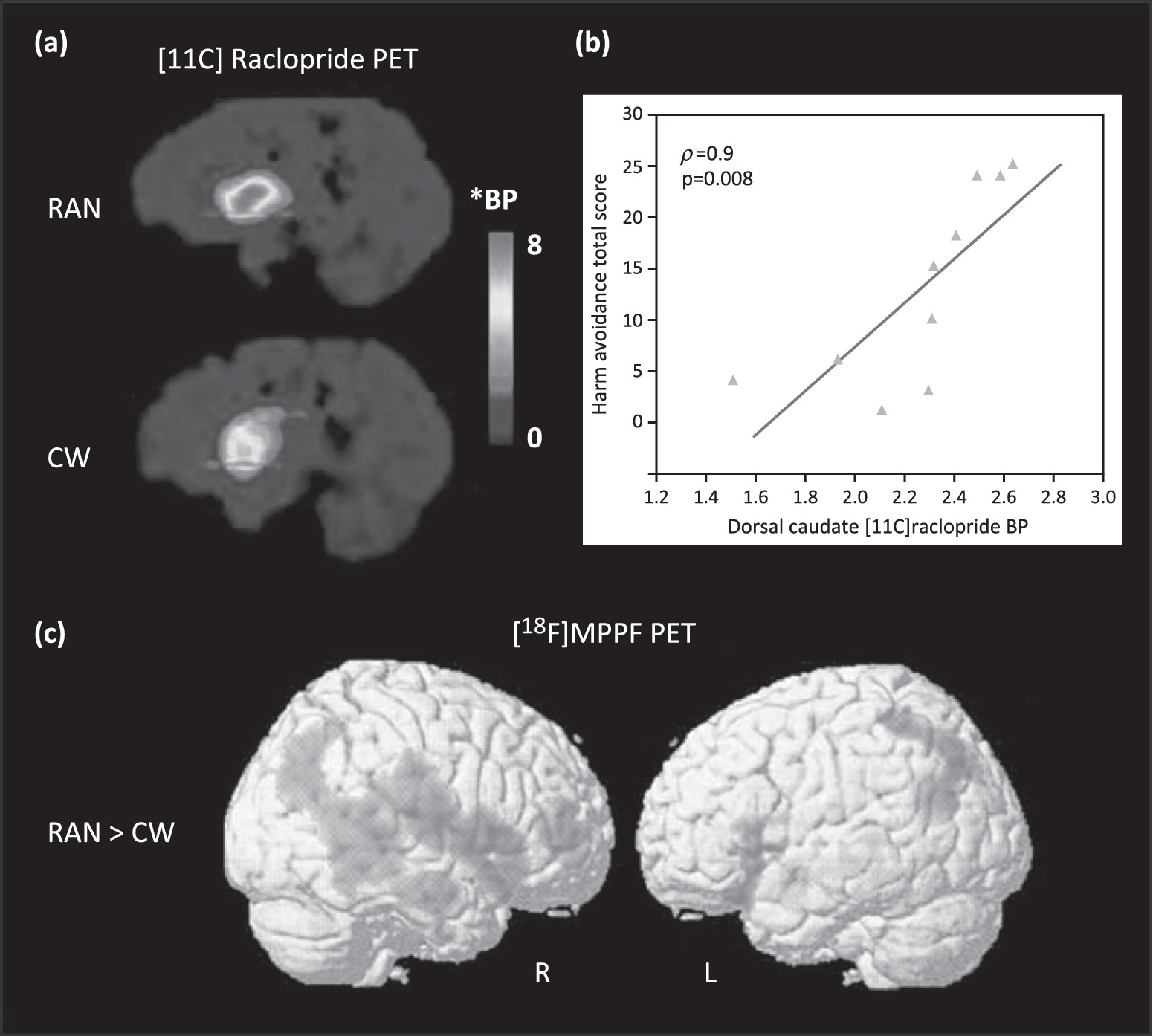
DA and 5-HT: relationship to HA
| Receptor/transporter | Imaging | III | Recovered | Regions | Direction of changeb | Refs |
|---|---|---|---|---|---|---|
| 5-HT2A | PET | – | AN | Mesial temporal (amygdala and hippocampus), pre- and subgenual cingulate, occipital/parietal cortical regions | – | [103] |
| – | AN-BN | Left subgenual cingulate, left parietal cortex, right occipital cortical regions | – | [104] | ||
| AN, AN-BN | – | – | = | [105] | ||
| SPECT | AN | – | Left frontal, bilateral parietal, occipital cortical regions | – | [106] | |
| 5-HT1A | PET | n/a | AN | _ | = | [107] |
| n/a | AN-BN | Cingulate, lateral and mesial temporal, lateral and medial orbital frontal, parietal, and prefrontal cortical regions; dorsal raphe | + | [107] | ||
| AN, AN-BN | – | Cingulate, lateral and mesial temporal, lateral and medial orbital frontal, parietal, and prefrontal cortical regions; dorsal raphe | + | [105] | ||
| – | AN | Right superior temporal, inferior frontal gyrus; parietal operculum, temporoparietal junction; left amygdala, parahippocampal gyrus, left temporal pole | + | [108] | ||
| AN | n/a | Right superior temporal, inferior frontal gyrus; parietal operculum, temporoparietal junction; left amygdala, parahippocampal gyrus, left temporal pole | + | [108] | ||
| 5-HTT | PET | – | AN | – | = | [57] |
| – | AN-BN | – | = | [57] |
| Receptor/transporter | III | Recovered | HA correlation | Regions | Refs |
|---|---|---|---|---|---|
| 5-HT2A | – | AN | No | – | [103] |
| – | AN-BN | Positive | Subgenual cingulate, mesial temporal | [104] | |
| AN, AN-BN | Positive | Supragenual cingulate, frontal, parietal | [105] | ||
| 5-HT1A | – | AN | Positive | Subgenual cingulate, mesial temporal | [107] |
| – | AN-BN | No | – | [107] | |
| DA D2/D3 | – | AN, AN-BN | Positive | Dorsal caudate, dorsal putamen | [18] |
| – | AN, AN-BN, BN | Positive | Dorsal caudate, dorsal putamen | [47] |
Alterations in the dopaminergic system
Alterations in the serotonergic system
Relationship between 5-HT, DA, and HA
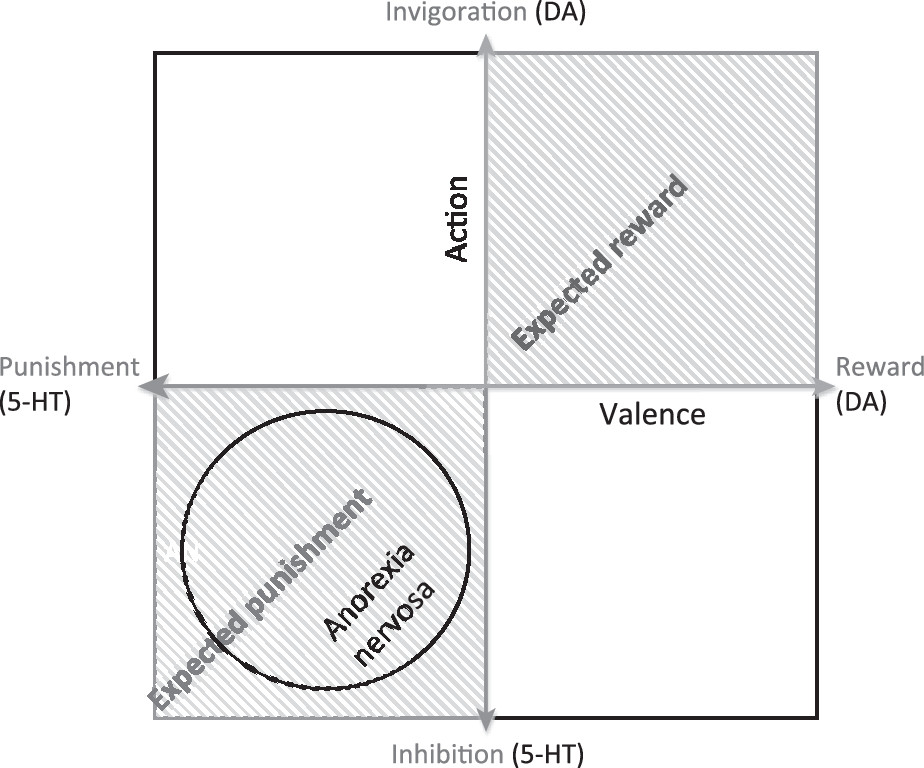
DA, anxiety, and restricted eating
Competition between DA and 5-HT
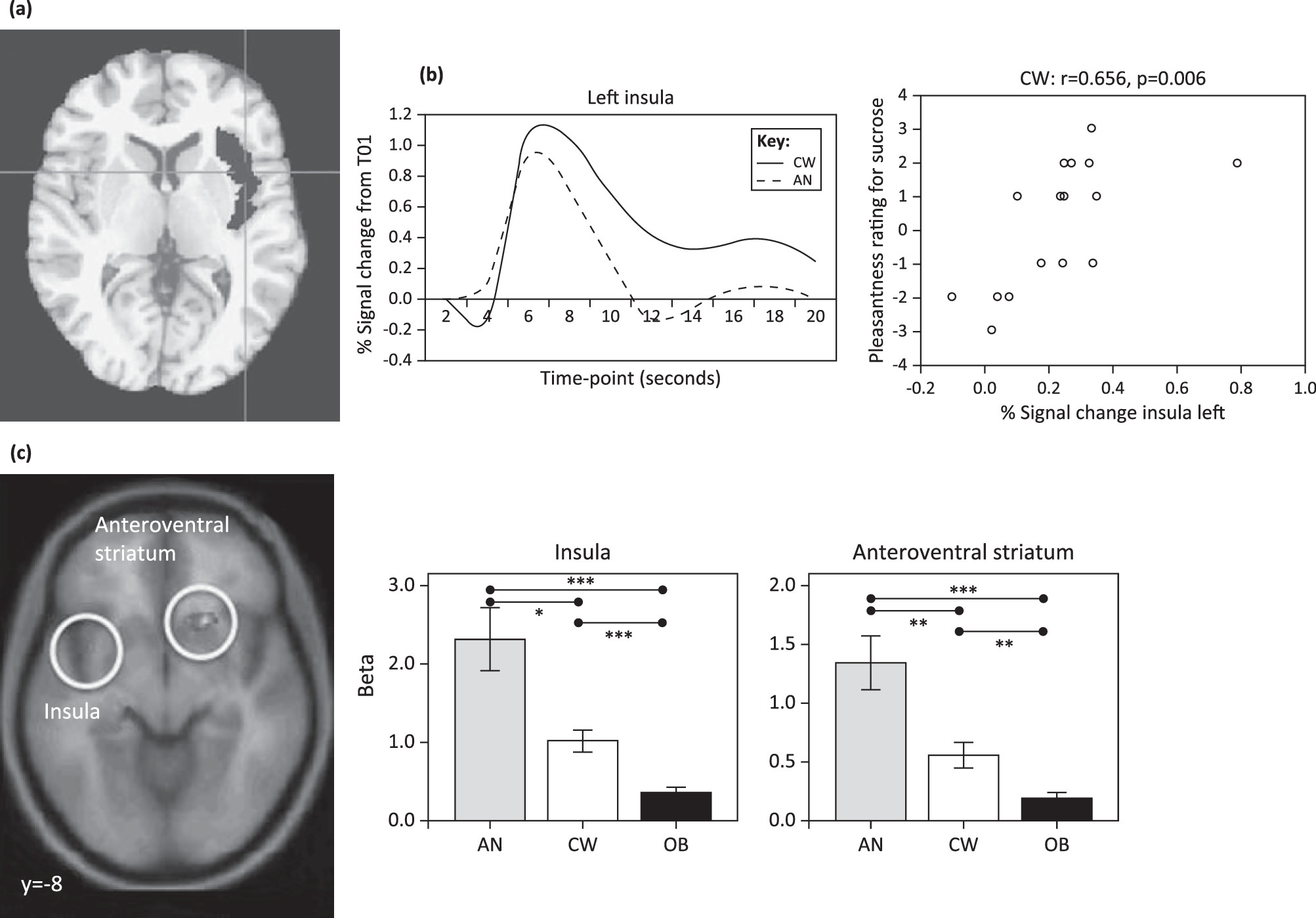
fMRI studies of appetite: relationship to food taste
fMRI studies of reward and executive control
Concluding remarks
Box 2. Outstanding questions
References
Information & Authors
Information
Published In


History
Authors
Metrics & Citations
Metrics
Citations
Export Citations
If you have the appropriate software installed, you can download article citation data to the citation manager of your choice. Simply select your manager software from the list below and click Download.
For more information or tips please see 'Downloading to a citation manager' in the Help menu.
View Options
View options
PDF/EPUB
View PDF/EPUBGet Access
Login options
Already a subscriber? Access your subscription through your login credentials or your institution for full access to this article.
Personal login Institutional Login Open Athens loginNot a subscriber?
PsychiatryOnline subscription options offer access to the DSM-5-TR® library, books, journals, CME, and patient resources. This all-in-one virtual library provides psychiatrists and mental health professionals with key resources for diagnosis, treatment, research, and professional development.
Need more help? PsychiatryOnline Customer Service may be reached by emailing [email protected] or by calling 800-368-5777 (in the U.S.) or 703-907-7322 (outside the U.S.).