M ost studies of glucocorticoid responsiveness have used measures that reflect glucocorticoid actions in the periphery or challenge strategies that fail to distinguish peripheral from central effects.
1 Central glucocorticoid effects in posttraumatic stress disorder (PTSD) are suggested by the efficacy of cortisol administration in PTSD prophylaxis and treatment,
2 –
4 which may relate to alterations in cortisol, catecholamine, and corticotrophin releasing factor level in PTSD.
5 –
7 We recently reported that an intravenous bolus of 17.5 mg hydrocortisone sodium succinate (HCORT) in PTSD preferentially affected memory performance in combat veterans with PTSD, with different effects in younger versus older PTSD cohorts,
8 –
9 which may reflect age-related changes in brain glucocorticoid responsiveness.
10 –
12 The aim of our present study was to examine central effects of glucocorticoids on relative glucose metabolic rate (rGMR) in the hippocampus, dorsal amygdala, ventral amygdala, and anterior cingulate cortex (ACC). These regions have been implicated in PTSD pathophysiology
13 –
16 and are known to be regulated by glucocorticoids.
17 –
19 Studies examining activity of these regions of interest in response to presentation of traumatic or emotional stimuli have demonstrated alterations in metabolic activity of these structures as well as in their interconnections.
20 –
35 In particular, amygdala activation has most often been demonstrated to be increased in PTSD, while ACC activity is diminished.
20 –
24,
36 However these differences vary by psychological task, with ACC activation in PTSD greater than in comparison subjects during trauma script tasks.
37 Because the ACC normally inhibits amygdala activity in response to stress, the neuroimaging findings support an impaired regulation of the neural circuitry connecting the amygdala to the ACC, possibly observed even under baseline conditions.
The amygdala projections leading to the ACC originate in the dorsal region of the amygdala, whereas the ventral amygdala, comprised of the basolateral complex, projects to orbitofrontal regions.
34 The basolateral complex has been specifically implicated in glucocorticoid enhancement of memory,
19 as shown on fMRI studies,
30,
34 justifying the separate examination of the dorsal and ventral regions in PTSD.
In this study, brain metabolism was examined in response to placebo and an intravenous bolus of HCORT, administered in a randomized, double-blind manner, and quantified using [
18 F]fluorodeoxyglucose (FDG) positron emission tomography (PET) neuroimaging. We hypothesized a greater response to the effects of HCORT in the PTSD+ group, consistent with enhanced responsiveness to glucocorticoids. In view of findings implicating laterality differences in PTSD, including reports of either greater group differences by hemisphere, or difference confined to one hemisphere,
20,
22 –
24 we further hypothesized that placebo rGMR might differ according to hemisphere based on group, and to the extent that this would be true, the effects of HCORT might also differ by hemisphere.
DISCUSSION
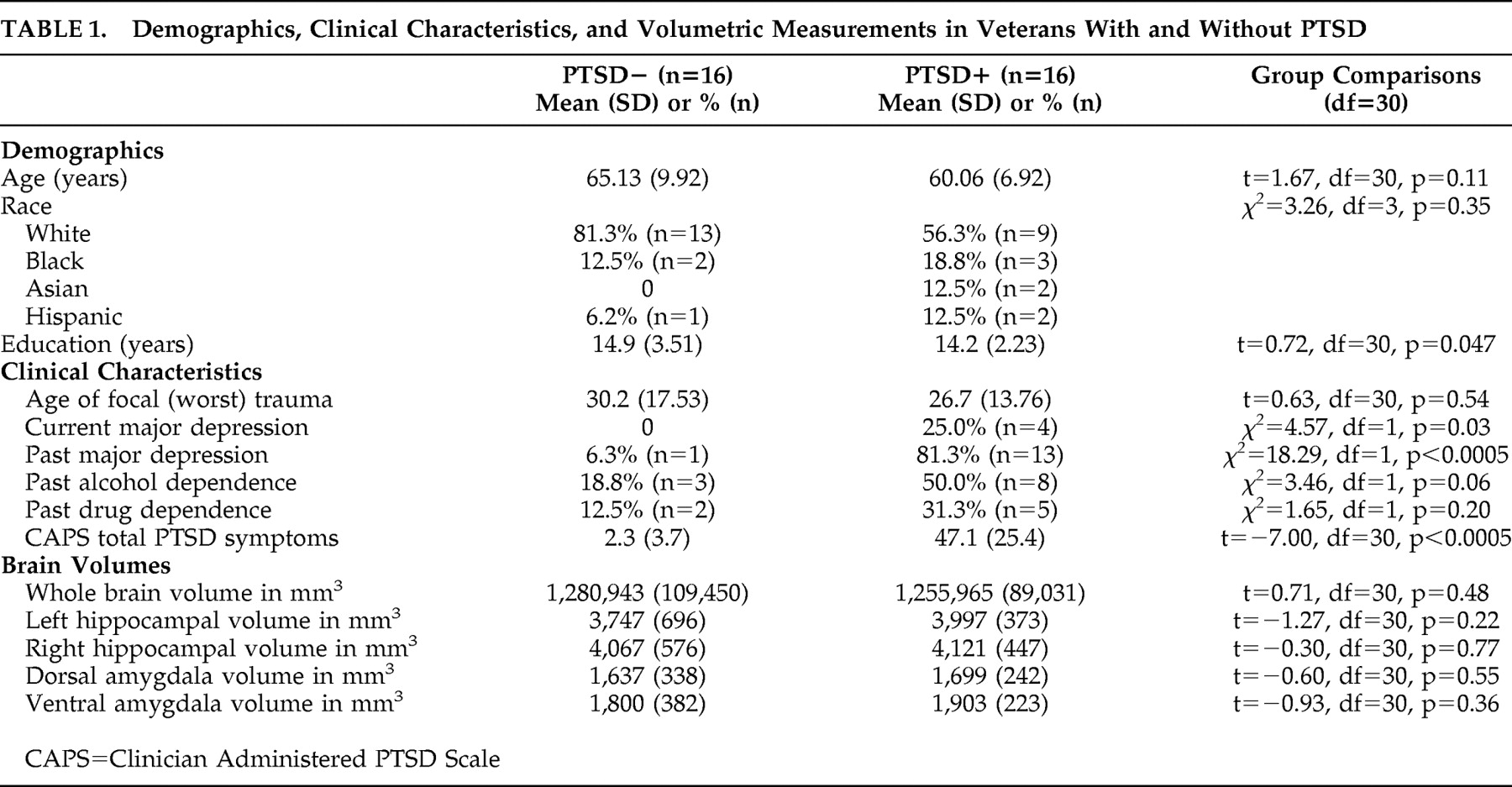
The results demonstrate regional differences in response to HCORT. The PTSD+ group showed a decrease in activity in the ACC with HCORT while the PTSD− group showed an increase, which reflects differences in central glucocorticoid responsiveness. The effect of HCORT was more prominent in the gray than white matter, consistent with a greater concentration of glucocorticoid responsiveness in gray than white matter in the human prefrontal cortex.
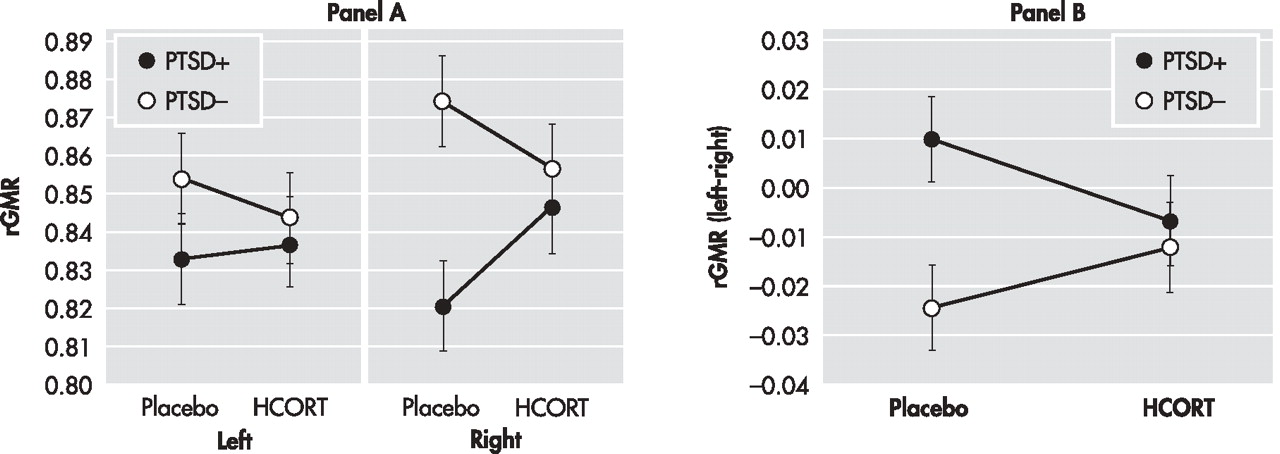
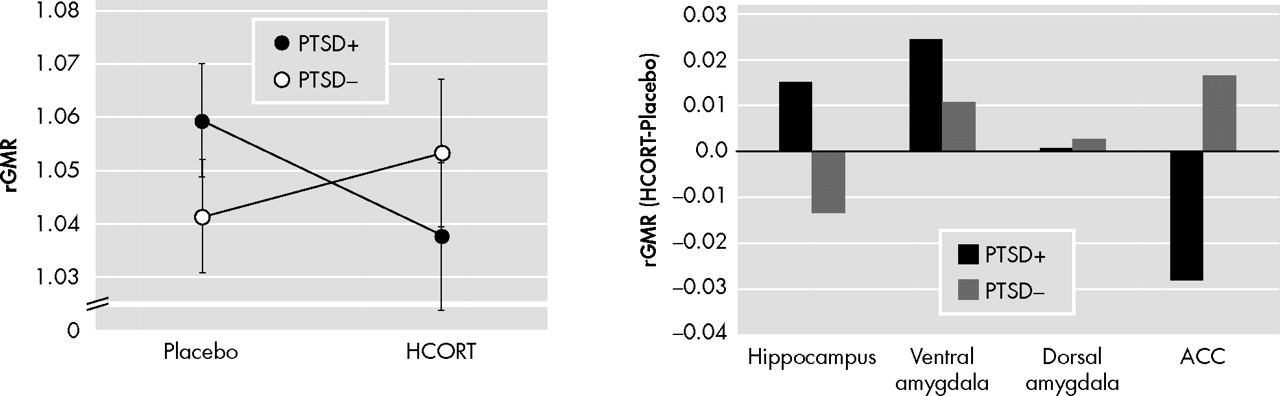
53 However, the pattern of decreased rGMR following HCORT was not recapitulated in the hippocampus and amygdala. Compared to the PTSD− group, the PTSD+ group responded to HCORT by increasing metabolism in the right hippocampus and the right ventral amygdala.
The effect of HCORT administration in the PTSD− group is consistent with findings in similarly aged healthy adults who also showed decreased hippocampal metabolism without effect in the prefrontal cortex following injection of an even larger dose of HCORT.
39 The negligible response to HCORT in the ACC in the PTSD− group is compatible with the demonstration of a relatively modest effect on rGMR in the ACC in combat comparison subjects following exposure to an olfactory stimulus designed to trigger traumatic memory compared to a neutral olfactory condition.
25 Yet, in the PTSD+ group, this exposure resulted in increased rGMR in the ACC compared to the neutral condition. Some recent studies have demonstrated increased activity in the medial prefrontal cortex/ACC in PTSD in response to a script-driven provocation paradigm,
26 and following nonconscious processing of fear.
27 However, the majority of reports demonstrate decreased activity in response to provocation,
20 –
23,
28 leading to the hypothesis that the medial prefrontal cortex is hyporesponsive in PTSD. Similarly, many studies in PTSD report increased brain activity in the amygdala in response to provocation,
13,
22 –
23,
25 as observed in the ventral amygdala in the current study in response to HCORT. Together, the findings of decreased activity in the medial prefrontal cortex/ACC and increased activity in the amygdala during various conditions in PTSD lead to the hypothesis that disruptions in the normal neurocircuitry of these regions manifest as a failed containment of amygdala activity by the medial prefrontal cortex/ACC.
39In the present study, stronger effects were detected in the ACC than hippocampus or amygdala, possibly attributed to a greater concentration of glucocorticoid responsiveness in the prefrontal cortex.
54 Interestingly, regardless of the extent or direction of group differences in rGMR on the placebo day, the effect of HCORT, in all cases, was to reduce the differences observed on the placebo day. In the hippocampus and amygdala, this was manifest by decreasing group differences in hemispheric laterality, whereas in the ACC this was demonstrated by a reduction in rGMR in the PTSD+ group.
The current findings showing hemispheric differences may reflect the fact that the right hemisphere operates with a greater range of activity—thus, the addition of HCORT would be greater due to increased variation. However, differences in hemispheric laterality may underlie differences observed regarding right and left hemispheric differences in provocation-related activation patterns, in association with ACC activity, in the previously published literature. To date, almost every study examining the hippocampus, amygdala, or ACC has found effects to be restricted to one hemisphere, or directionally different in the two hemispheres.
17,
19,
21,
22 That there is an association between brain laterality and PTSD risk has recently been confirmed in a large epidemiologic study demonstrating mixed lateral preferences for handedness was associated with PTSD.
55 Differences in handedness do not explain the current findings because only one subject in the sample was left handed. However, the original hypothesis regarding brain laterality emphasized that those with a reduced cerebral lateralization may be less able to detect and/or engage in corrective cognitive processing of threatening stimuli.
56 In the current study, there was less laterality in hippocampal and amygdala rGMR in PTSD on the placebo day, and HCORT reduced these group differences by increasing lateralization in the PTSD+ group and decreasing it in the PTSD− group. Resting data with
18 FDG-PET can assess trait-like laterality differences present without a specific psychological stimulus. Blood-oxygen-level-dependent (BOLD) activation studies are only able to interpret laterality (or hemispheric) differences in connectivity between PTSD and comparison groups in response to stimulation. Yet, numerous such observations have been made, supporting the importance of laterality differences in brain function in PTSD.
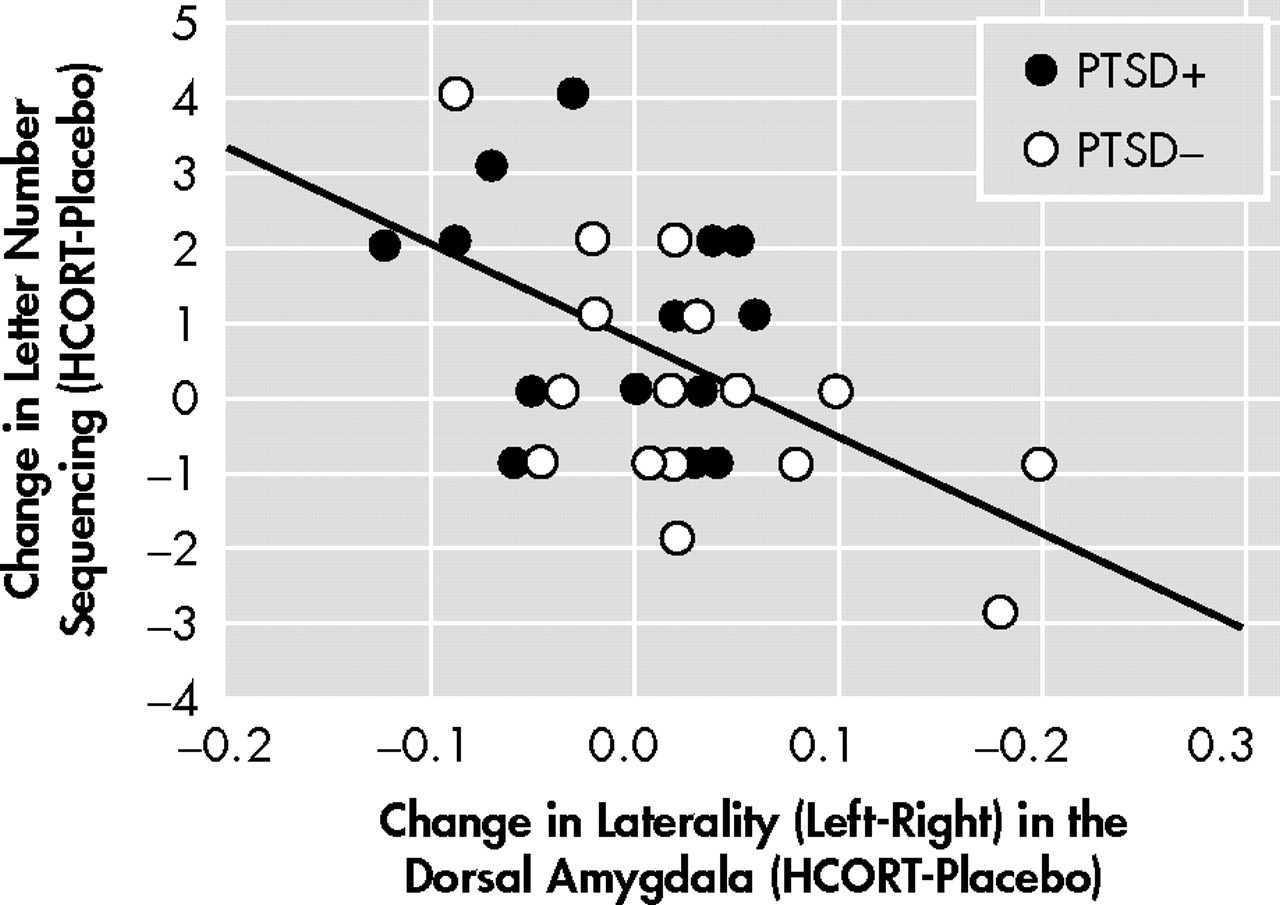
Interestingly, the only region to demonstrate an activation-related association with improvement in working memory was the dorsal amygdala. Though group differences for this region did not approach statistical significance, the correlation between brain activity in this region and memory performance is compatible with results of animal studies demonstrating that glucocorticoid induced effects on memory are enabled by noradrenergic activation of the basolateral amygdala.
57The findings add to the literature not only in their demonstration of group differences in the effects of HCORT, but also in their description of group differences in rGMR under resting conditions without pharmacological challenge. The differences on the placebo day, in particular, provide a context for interpreting data in response to provocation. Previously reported group differences in the direction of amygdala and/or ACC activity in response to provocation may be misleading without consideration of baseline activation. Had our study simply examined metabolic effects of HCORT administration on PTSD+ compared to PTSD− we would have likely concluded that effects were minimal, if not absent.
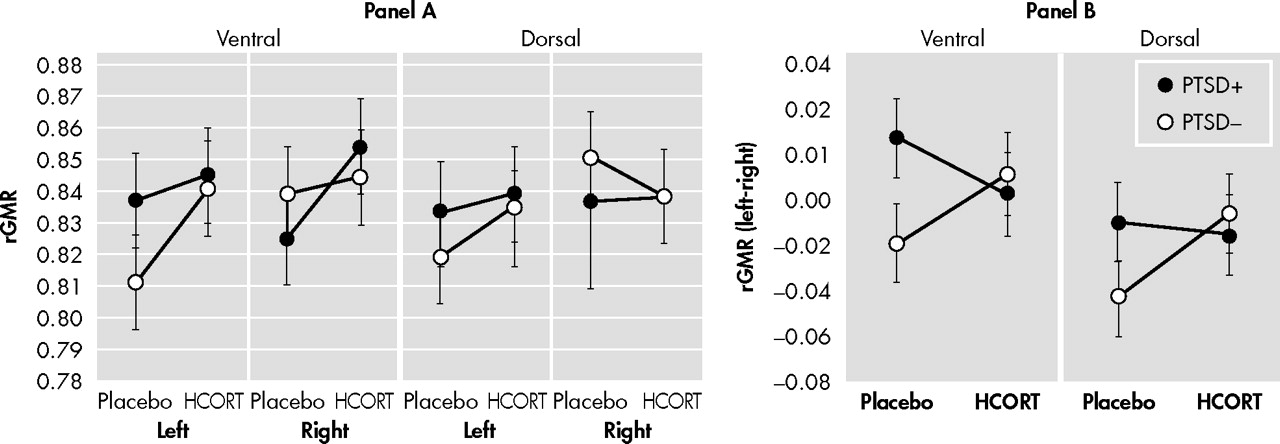
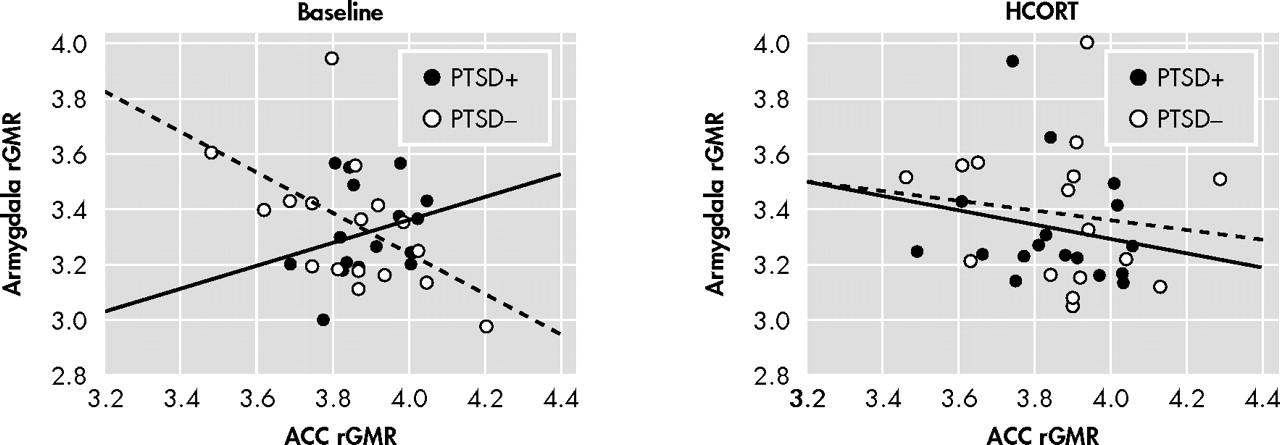
The negative correlation between ACC and amygdala metabolism observed on the placebo day in the PTSD− group implies that this neural circuit may regulate amygdala activity even under ambient conditions.
58 –
60 In the present study, this relationship was absent, and even in the opposite direction (although nonsignificant), in subjects with PTSD. Furthermore, in PTSD the rGMR in the ACC under placebo conditions was increased relative to comparison subjects. It is difficult to interpret the effect of this activity on the amygdala since right and left hemispheres showed different responses in PTSD. In that the PTSD+ group on the placebo day showed greater activity of the amygdala in the left, but lower activity in the right compared to the PTSD− group, the placebo ACC/amygdala relationship can either be interpreted as inverse or positive depending on the hemisphere. In response to a single dose of HCORT, the associations between ACC and amygdala metabolism were no longer discrepant between the PTSD+ and PTSD− groups, nor were differences in these associations present in the two hemispheres. It is particularly noteworthy that the effect of HCORT resulted in similar associations between amygdala and ACC activity in both groups on the active treatment day since the effect of HCORT in PTSD+ subjects was to restore a normal inverse association, while PTSD− subjects experienced a disruption of this normal neural network. A limitation of the study is that without a healthy comparison group not exposed to combat it is unclear whether the PTSD− group also represents a group with altered glucocorticoid function. Additionally, as we only studied male veterans, these results may not be generalizable.
Regarding possible treatment implications, the beneficial effects of glucocorticoids have been investigated and continue to be of interest, particularly in PTSD prophylaxis.
3,
61 The current observations are consistent with the beneficial effects noted on memory observed in this older cohort. However, in a younger cohort HCORT resulted in impairment of working memory,
8 consistent with a larger literature in healthy subjects.
62 –
63 Regionally specific differential responses to glucocorticoids have been noted according to age,
64 –
69 and their relevance should be fully explored since it may be that therapeutic approaches that are optimal at some phases of illness are contraindicated in others based on age, chronicity, or other factors contributing to dynamic changes in pathophysiology and brain function. Little is known about the efficacy of standard treatments of PTSD in the elderly. However the current discrepancies between aging PTSD and normal aging profiles suggests that we cannot extrapolate on the basis of the literature on normal aging to what might be true as persons with PTSD advance in age.