Suicide is a complex, heterogeneous phenotype as well as an intractable public health problem, with the overall annual suicide rate remaining stable over the past 60 years at around 10 to 12 per 100,000 (
1). The National Action Alliance for Suicide Prevention has set out to build a research agenda with the potential to reduce our national suicide rate by 20% within 5 years (
2). One strategy proposed is to identify and target subgroups at greatest risk. Heuristic models outlining the chain of events leading to suicide often include biological or genetic characteristics, early life events including trauma and other stressors, impulsive aggressive traits, psychopathology, inadequate social support, and access to lethal means (
3–
5).
A growing body of evidence suggests that suicide vulnerability may be due to epigenetic alterations in molecular pathways important for hypothalamic-pituitary-adrenal (HPA) axis function. For example, DNA methylation changes in the
NR3C1 gene, which encodes the glucocorticoid receptor, are altered by maternal behavior in rats (
6) and are higher than normal in the hippocampus of suicide decedents who experienced early life trauma (
7). The cumulative effect of these epigenetically mediated events is a reduction in glucocorticoid receptor levels, possibly leading to impaired responses to stressors. Suicidal individuals exhibit less ability to suppress cortisol after experimental administration of the synthetic glucocorticoid dexamethasone (
8), and the cortisol stress response has been identified as one of the most promising candidate suicide endophenotypes (
3). Other studies have provided evidence that first-degree relatives of suicide decedents fail to mount a proper HPA axis response to stress (
9). Such findings are consistent with the diathesis-stress or dual-risk hypothesis, whereby an underlying biological state moderates an aberrant response to stress (
10–
14). Identification of the underlying genetic and epigenetic factors influencing vulnerability to suicidal behaviors in the context of stressors is needed to maximize suicide prevention efforts.
The objective of this study was to use genome-wide screening techniques to identify novel epigenetic associations in postmortem brain tissue of suicide decedents, followed by replication and functional assessment of identified loci. A secondary objective was to assess the degree to which identified loci would be present in peripheral blood samples and to evaluate their biomarker efficacy in the context of stress and anxiety.
Discussion
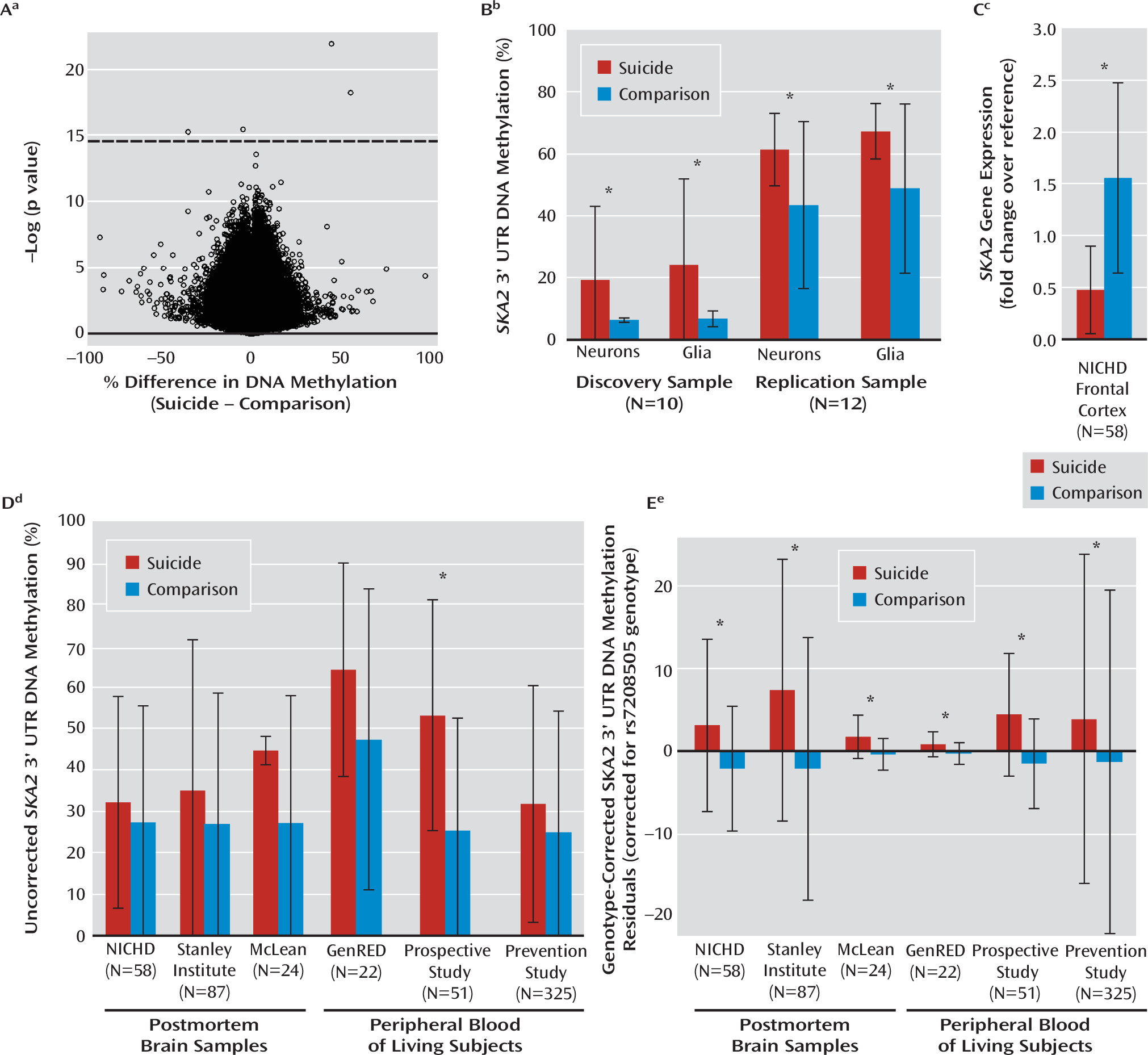
Using microarray technology to scan for epigenetic suicide associations, we identified a significant effect in a very small group of suicide decedents. The effect size of about 55% DNA methylation difference at SKA2 enabled this small study group to have adequate power to survive correction for multiple testing, which was driven by the underlying genetic status of the rs7208505 SNP, which abrogates the CpG dinucleotide. While microarray analysis was performed only in Caucasians, incorporation of both the genetic and epigenetic variation at this locus enabled replication across the entirety of the NICHD cohort, two additional postmortem brain cohorts, and three blood cohorts. Despite the consistency of the findings, the relatively small sizes of the studied cohorts suggest they represent promising but preliminary results warranting further study. The presented linear models suggested that DNA methylation and rs7208505 genotype may have opposing effects on suicidal behavior; however, as these metrics were highly correlated, the protective effects of rs7208505 may represent a statistical artifact. Analysis of genetic and epigenetic effects on suicidal behavior and gene expression separately indicated that DNA methylation alone may be the primary factor conferring risk. It is important that the overall proportion of DNA methylation at rs720505 increased significantly with each successive C-containing allele, suggesting that the underlying genetic architecture at rs7208505 may confer vulnerability by providing a genetic template for methylation changes to occur. This risk template would be expected to vary as a function of ethnicity, as allele frequencies for the C-containing allele of rs7208505 are reportedly much lower in African Americans (about 18%) than in other ethnicities (50%−60%). Cumulatively, numerous consistent associations were observed with suicidal ideation, suicide attempt, and suicide completion, independent of variation in ethnicity and psychiatric diagnosis, suggesting that variation in SKA2 may mediate risk for suicidal behaviors that progress from ideation to attempt to suicide.
SKA2 may influence suicidal phenotypes through its role in chaperoning the glucocorticoid receptor from the cytoplasm to the nucleus. Rice et al. (
21) demonstrated that knockdown of
SKA2 eliminated glucocorticoid receptor transactivation and response to dexamethasone treatment in vitro and that protein levels of SKA2 were decreased by glucocorticoid treatment, suggesting that
SKA2 gene expression may be a component of the glucocorticoid feedback inhibition system. In our data,
SKA2 genetic and epigenetic differences were associated with reduced suppression of salivary cortisol after waking in the GenRED cohort. As the blood was not drawn at the same time as the salivary cortisol sampling, the causative role of DNA methylation must be interpreted cautiously. While DNA methylation variation at rs7208505 might be important for suicidal ideation etiology, it remains possible that this variation is a reflection of cortisol variation.
In the proposed model,
SKA2 epigenetic and genetic variation represents an underlying state increasing suicide risk in the presence of a stressor.
SKA2 epigenetic and genetic variation interacted with stress and anxiety metrics to mediate suicidal ideation in the prospective cohort, while in the Prevention Research Center and GenRED offspring cohorts, the same model distinguished individuals with suicidal ideation who transitioned to suicide attempt. It is important to note that the suicidal ideation, suicide attempt, and suicide phenotypes are not interchangeable; however, in some individuals they represent progressive stages of suicidal behavior that share many etiological factors. The proportion of variance accounted for by our models was very high in some cohorts and leaves little room for the involvement of other factors. While our data suggest that
SKA2 may be etiologically relevant to glucocorticoid signaling, it is possible that the detected epigenetic variation at
SKA2 also represents a molecular record of suicide-dysregulated glucocorticoid load over time and thus may be reflective of other sources of etiologically relevant variation at other loci implicated in suicide. In a recent study, blood gene expression at
SAT1 prospectively predicted both suicidal ideation and suicide attempt (
23). Our supplemental analysis demonstrated an interaction between
SKA2 variation and DNA methylation at a CpG in the
SAT1 promoter located within a region enriched for glucocorticoid receptor binding.
SKA2-mediated failure to suppress normal stress response may play a role in
SAT1 gene expression variation and could contribute to the transition from suicidal ideation to suicide attempt. Cumulatively, our data are consistent with findings from an epidemiological study of 108,664 individuals in 21 countries that found disorders characterized by anxiety and poor impulse control predicted the transition from suicidal ideation to suicide attempt (
24).
One caveat with these analyses is that different metrics of suicidal ideation, suicide attempt, stress, and anxiety were available across the studied cohorts. In the Prevention Research Center and prospective study groups, anxiety was measured by response to a single question, while the Screen for Child Anxiety Related Disorders (SCARED) was used for the GenRED offspring. In the prospective cohort, we showed that anxiety, as measured by the Edinburgh Postnatal Depression Scale, was highly associated with perceived stress, which in another study was correlated with salivary cortisol level (
25). Thus, it is possible that the observed interactions of
SKA2 with anxiety across cohorts are a reflection of underlying differences in stress and HPA axis response in anxious individuals. Despite these limitations, the ability of the linear model generated from the Prevention Research Center cohort to accurately predict both suicidal ideation and suicide attempt in the prospective and GenRED cohorts suggests that consistency was captured by these diverse metrics.
The lower
SKA2 gene expression in NICHD brains from suicide decedents was associated primarily with isolated neuronal nuclei, suggesting the epigenetic dysregulation may be confined to neurons. The postmortem brain data were generated in the prefrontal cortex, a brain region with inhibitory connections to the HPA axis (
26,
27) and responsible for decision making, inhibition of negative thoughts, and impulsivity (
28,
29). Low glucocorticoid receptor transactivation is consistent with current models of suicide diathesis, as experimentally reduced glucocorticoid receptor gene expression in rodents mimics suicidal human characteristics (
8), producing increases in corticosterone and helplessness in response to stress (
30).
The influence of
SKA2 3′ UTR epigenetic variation on gene expression appeared to be mediated by interaction with epigenetic variation within the gene promoter and proximal to intronic miR-301a, which has previously been shown to be low in the postmortem prefrontal cortex of individuals who die by suicide (
31). Critically, miR-301a modulates
SKA2 gene expression in A549 cell models by indirectly inhibiting CREB binding to the
SKA2 promoter (
32), while the promoter CpGs shown to correlate with rs7208505 DNA methylation directly flank this CREB binding site. As would be expected given the model, we observed a significant effect of the interaction of miR-301a and promoter CpG variation on
SKA2 expression. The observed correlations of
SKA2 3′ UTR DNA methylation with other CpGs across the gene could result from common epigenetic reprogramming effects of glucocorticoid receptor binding, as the regions demonstrating correlations were located within glucocorticoid receptor immunoprecipitation peaks identified from Encyclopedia of DNA Elements (ENCODE) data. It is also possible that miR-301a proximal genetic variation in linkage disequilibrium with rs7208505 serves to alter glucocorticoid receptor recruitment, subsequently reprogramming DNA methylation in the region, as discussed in the online supplementary material. The miR-301a is an intronic microRNA and requires mRNA transcription of
SKA2 to be generated by Drosha, an RNase III enzyme (
33). DNA methylation upstream of miR-301a may therefore result in cotranscriptional slowing and allow for spliceosomal interaction as occurs with inclusion of methylated alternative exons in alternatively spliced genes (
34). Elevated neuronal but not glial DNA methylation levels proximal to miR-301a suggest a possible functionally different effect of miR-301a in these two cell types. It is important to note that while epigenetic variation proximal to the miR-301a and CREB binding site was associated with
SKA2 gene expression, it was not associated with suicidal phenotype.
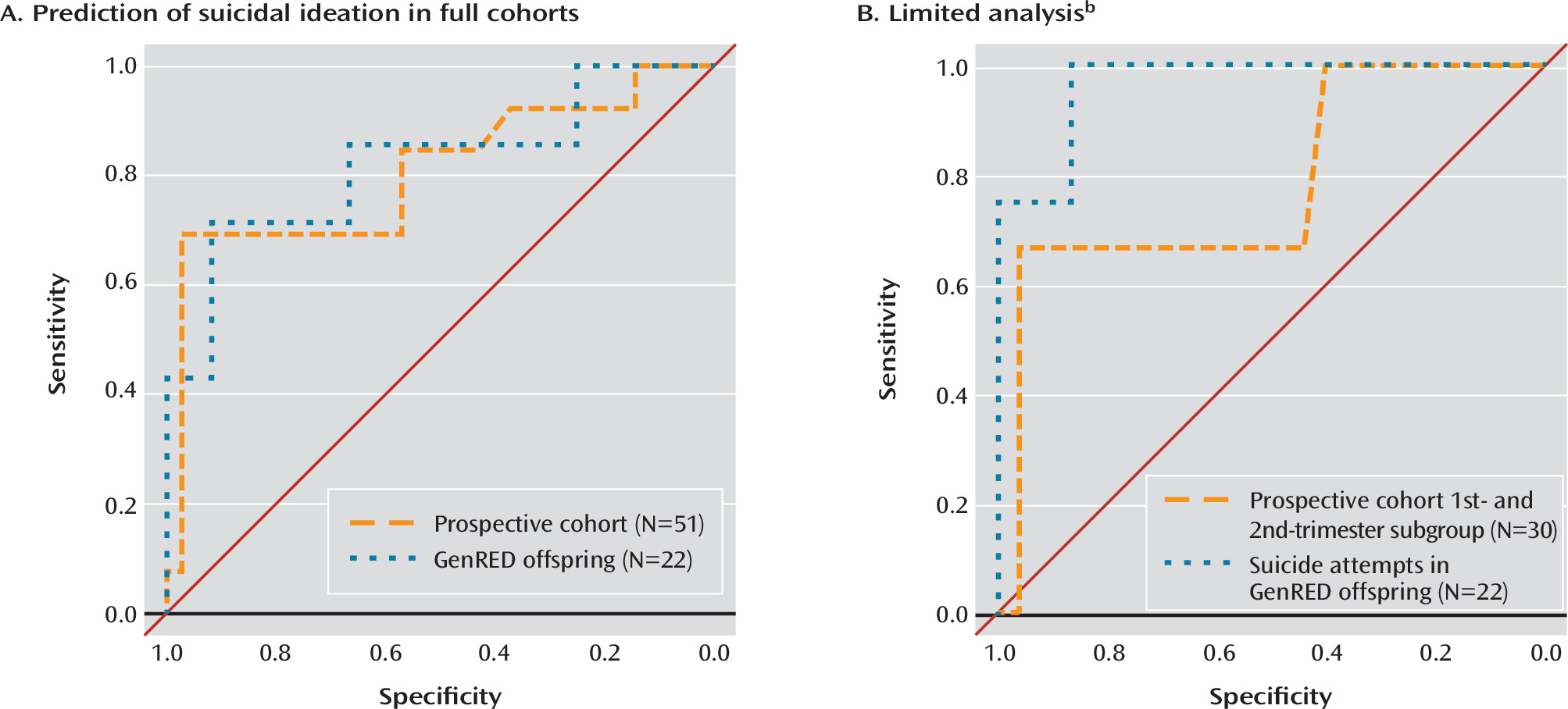
While a growing number of studies are investigating epigenetic alterations in suicide (
23,
35–
40), few studies have identified biomarkers with high prediction accuracy. To our knowledge, the biomarker identified in this study represents the first genetic and epigenetic biomarker capable of predicting suicidal ideation and suicide attempt in a prospective manner with over 80% accuracy from blood. The model performed remarkably well at predicting suicide attempt in the GenRED cohort; however, with only four attempters, this result should be interpreted with caution. While the Prevention Research Center cohort contained many more suicide attempt cases, we did not test prediction in this cohort as the time between suicide attempt and blood draw was greater than 10 years on average. Accumulating epigenetic change due to stochastic drift, substance use, and errors in retrospective reporting would call into question the reliability of the prediction. However, this highlights the fact that the cause versus effect of prediction accuracy in the GenRED offspring cohort must also be interpreted with care as the blood was taken after the suicide attempt. Nevertheless, our data demonstrate similar accuracies when predicting suicidal ideation in a prospective manner, suggesting that
SKA2 epigenetic and genetic variation may represent a trait influencing underlying suicide risk when interacting with stress. Cumulatively, the clinical implications of this finding are that early screening of those at risk for suicidal ideation and suicide attempt may be possible, allowing for the identification of individuals at risk, proactive treatment, and stress and anxiety reduction. The potential efficacy of this biomarker is relevant to numerous populations, for example, the military, where the identification of an underlying vulnerability may identify individuals at risk for developing suicidal behaviors when exposed to the stress of war-time situations. Future studies should be carried out to further evaluate the prospective efficacy of this finding in additional populations.