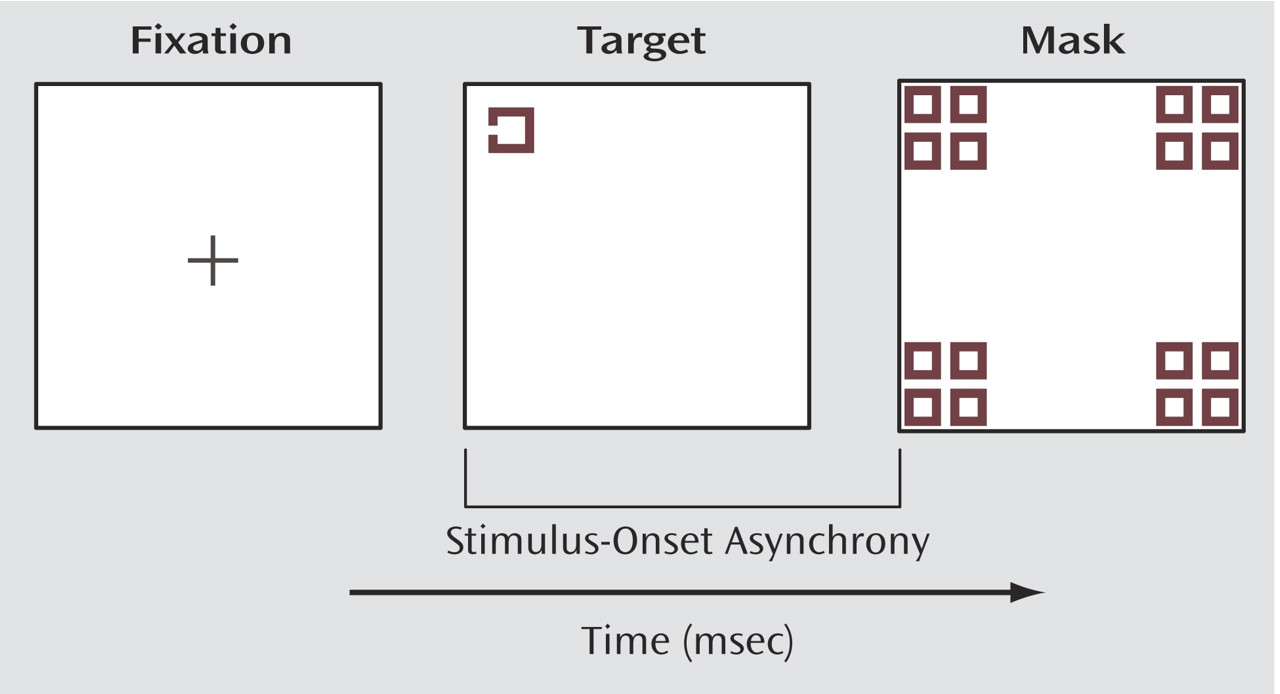
The purpose of the current study was to expand upon our preliminary findings of gamma abnormalities during masking in schizophrenia and examine in greater detail how gamma activity is influenced by the strength of the masking effect. The backward-masking task we employed in this study is an appropriate way to study gamma activity because participants must discriminate the form of a target from the mask and process contour information of that target. Perception of visual form presumably depends upon integrating basic visual elements (e.g., line or edge orientation) to construct higher-order conjunctions (e.g., angles or vertices), and finally, to form a coherent representation of an object. It is believed that a key role of gamma activity is to achieve this kind of binding together of visual features. In this study, we examined the effects of masking interval, unmasked versus masked trials, correct versus incorrect trials, and brain region on gamma activity in schizophrenia patients and healthy comparison subjects.
Discussion
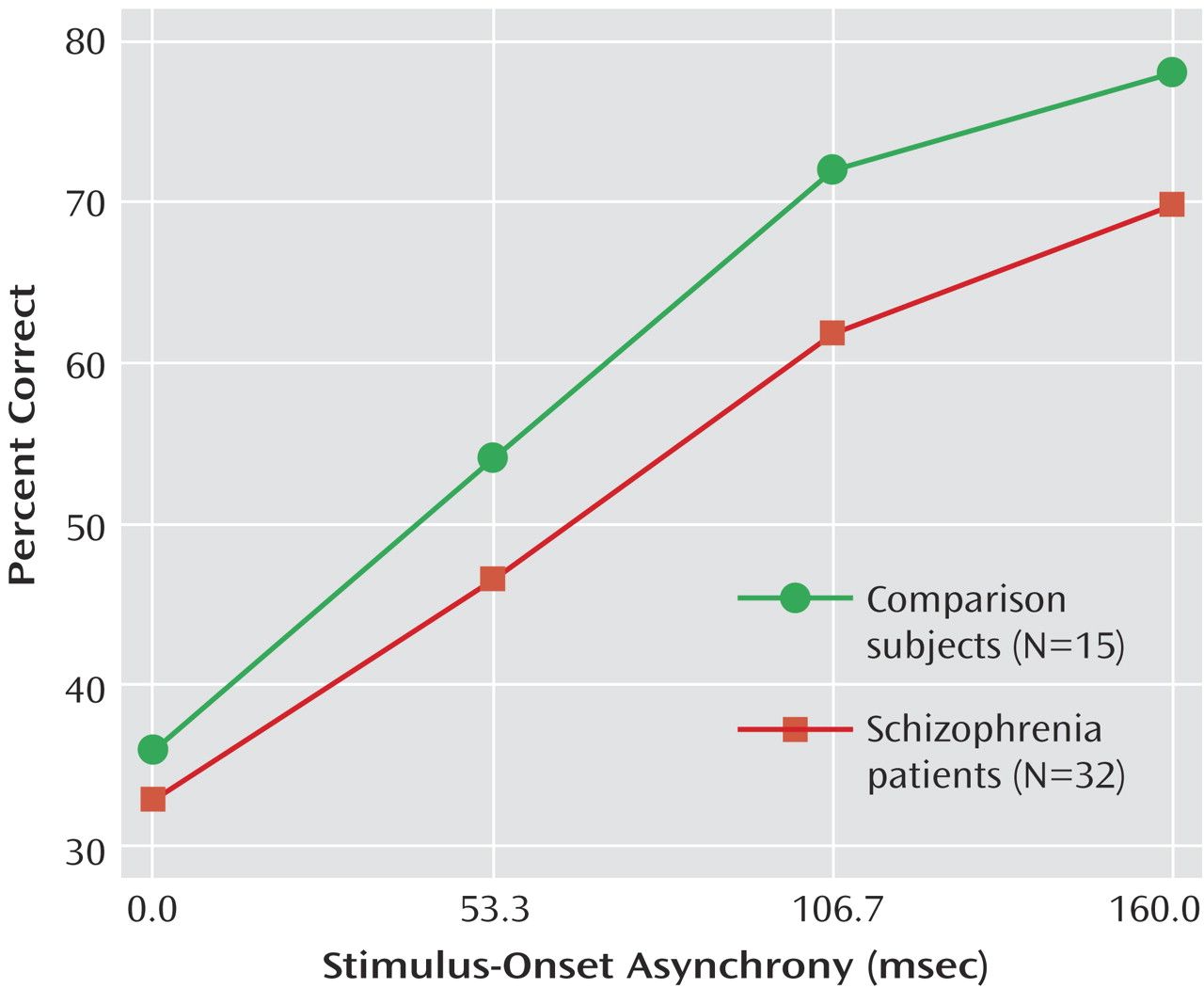
The results of the present study provide further support for an event-related abnormality in gamma activity in schizophrenia patients. Group differences were seen both in the amount and distribution of gamma activity during the backward-masking procedure. These results, moreover, are specific to targets presented in the presence of a mask. The lack of a group difference in gamma activity to unmasked stimuli suggests comparable processing of unambiguous visual stimuli in patients and comparison subjects.
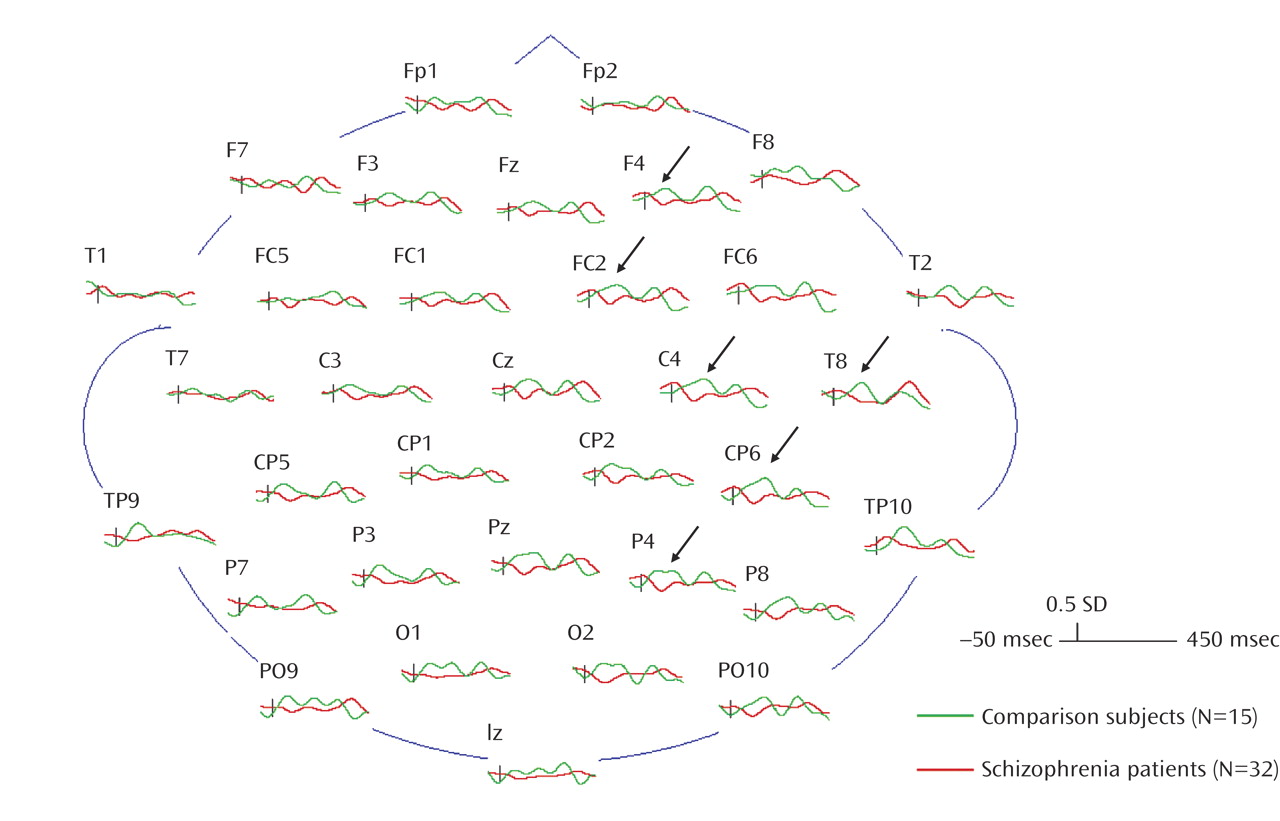
The finding of a group-by-hemisphere difference in gamma activity in masked trials is consistent with previous findings of a deficit in the right hemisphere in schizophrenia for processing visual information (e.g., references
20–22). For example, Nuechterlein and colleagues
(23) scanned subjects with positron emission tomography (PET) during a perceptually demanding continuous performance test and found that the schizophrenia patients did not use the right hemisphere preferentially to process ambiguous visual stimuli as normal subjects did
(24). With simple dot-pattern stimuli, Heckers et al.
(22) found that normal comparison subjects showed significant decreases in PET activity in the right hemisphere from the first viewing to the last viewing of the dot pattern, suggesting that habituation of activity in the right hemisphere is indicative of processing simple visual stimuli. The schizophrenia patients showed a reversed asymmetry, suggesting that their left hemisphere was overactive in processing visual stimuli. On the basis of the gamma activity findings of the current study, it appears that schizophrenia patients lack the normal right hemisphere lateralization to process ambiguous visual information and that this deficit extends to the backward-masking task.
The finding of a significant interaction between group, stimulus-onset asynchrony, and accuracy (correct versus incorrect) was due to a tendency for the comparison subjects to produce greater gamma activity to incorrect trials at a stimulus-onset asynchrony of 53 msec, whereas the schizophrenia patients showed the reverse pattern. This pattern occurred at only one stimulus-onset asynchrony and was not predicted. We do not have an interpretation at this time but will see if the pattern replicates itself.
The findings in the present study add to a growing literature demonstrating that schizophrenia patients exhibit reduced gamma activity to simple stimuli. Previous studies with auditory stimulation have shown that schizophrenia patients are less able to engage or maintain gamma activity
(11,
12). For example, Kwon et al.
(11) found that schizophrenia patients failed to show EEG synchronization to 40-Hz steady-state auditory trains. In a separate study, Clementz et al.
(12) found that gamma-band (e.g., 30–50 Hz) abnormalities in the auditory P50 suppression response were better than the broadband P50 response (e.g., 10–50 Hz) at detecting differences between patients and comparison subjects. Although these studies used a different modality than we did, they similarly examined time-locked, or evoked, gamma responses. The results across these studies suggest that schizophrenia patients may have abnormalities in establishing or maintaining gamma activity to simple auditory and visual stimuli. These abnormalities, in turn, may account for some of the perceptual deficits in schizophrenia.
Gamma oscillations at the neuronal level are thought to be modulated by γ-aminobutyric acid (GABA)-ergic interneuronal circuits (e.g., references
25,
26), as well as
N-methyl-
d-aspartic acid (NMDA)-modulated pyramidal cells (e.g., references
27,
28). Schizophrenia has been associated with abnormalities in the GABA and NMDA systems, which may explain the gamma abnormalities that have been observed. It is possible that such abnormalities in gamma synchronization could lead, through a cascade of processes, to mistakes in perceptual processing and misinterpretation of ambiguous stimuli.
Our study had a few limitations. The stimuli we used were very small (0.23° of the visual angle), which probably reduced the amount of gamma activity seen. Recent research has shown that the amount of gamma to visual stimuli increases with increasing size of the stimuli
(29). For the current study, we intentionally selected stimuli that were identical to those used in our behavioral studies
(30). These stimuli need to be relatively small and faint to establish each subject’s perceptual threshold.
Another limitation is that the study has a limited number of stimulus-onset asynchronies. Although this was intentional so as to make the length of the testing session tolerable to the participant, it had the disadvantage of making it more difficult to detect any stimulus-onset asynchrony-dependent, event-related gamma activity. Future studies that use a wider range of stimulus-onset asynchronies may be in a better position to detect stimulus-onset asynchrony-dependent gamma activity. On the basis of the results from the unmasked trials, we expect that gamma activity would be normal in schizophrenia patients at the longest stimulus-onset asynchronies when performance approaches unmasked levels.
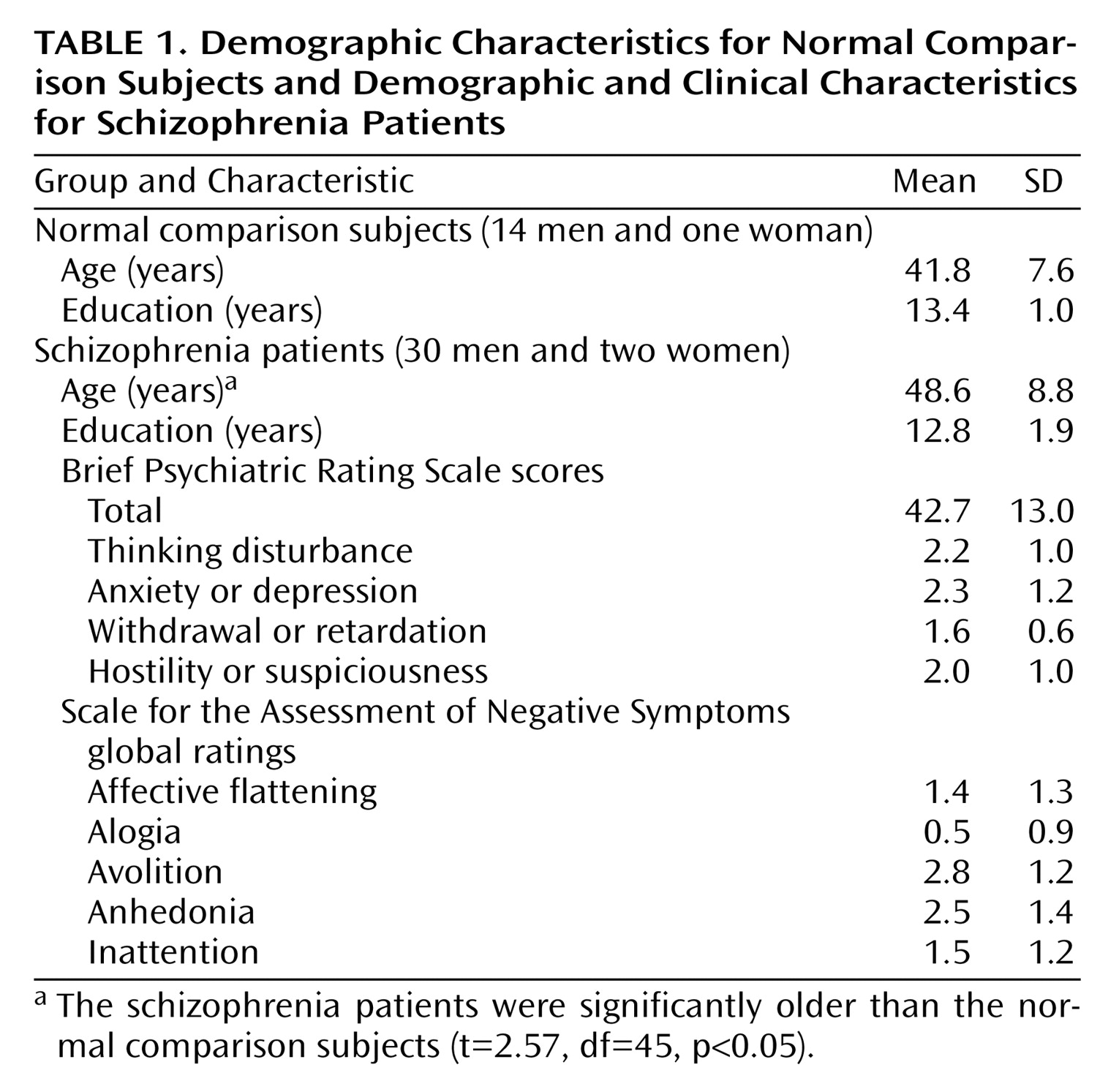
The patients and comparison subjects differed in age. However, as we noted, covarying age did not change any of the significant results. Age also did not correlate with any gamma measures of interest (e.g., overall gamma activity, gamma to correct versus incorrect trials, etc.), further reducing the possibility that our results are due to age differences. Age effects on performance were minimized by matching all subjects for unmasked target discrimination with a psychophysical staircase method.
Although visual-masking procedures appear to be excellent probes for aberrant gamma activity in schizophrenia, this study does not clarify whether abnormal gamma activity is the main cause of masking deficits in schizophrenia. Evidence has accumulated showing that schizophrenia patients have abnormal transient activity
(7,
31,
32), and this abnormality may partially account for visual-masking deficits. However, gamma-range activity in backward-masking tasks is considered to be a function of sustained channels
(8,
9). Therefore, although the differences in gamma activity in the current study are provocative, the role that abnormal gamma activity plays in masking performance deficits in schizophrenia patients remains unclear.
Finally, our study examined only evoked—but not induced—gamma activity. Evoked gamma activity is phase-locked to the onset of a stimulus and can be detected by averaging EEG responses. This is indicative of early sensory registration or processing of stimuli
(33). Induced gamma activity is a non-phase-locked activity, is not seen with traditional averaging techniques, and is indicative of perceptual organization or feature binding
(33). Because backward masking interrupts the very early stages of target processing, we were primarily interested in the early evoked gamma response (50–200 msec). However, a close examination of induced gamma activity in future studies could provide further insight into the gamma deficits seen in schizophrenia patients.
In conclusion, the current study extends the findings of abnormal gamma activity in schizophrenia patients during a backward-masking task
(10). The current study employed a masking paradigm to elicit gamma activity. Decreased gamma activity was seen in schizophrenia patients only during the masking task, consistent with theories that abnormalities of generating or maintaining gamma activity can contribute to perceptual aberrations in schizophrenia
(34). We further found that these deficits are especially pronounced in the right hemisphere, an area that is preferentially responsible for processing ambiguous visuospatial information. Although the current study has several limitations, these results continue to demonstrate the usefulness of using backward-masking procedures to explore underlying neurophysiological abnormalities in schizophrenia.