Schizophrenia has long been associated with abnormalities in information processing and attention mechanisms
(1,
2). Prepulse inhibition of the startle reflex, a cross-species phenomenon, provides a valuable opportunity to study such abnormalities
(3). Prepulse inhibition refers to a reduction in response to a strong startling stimulus, “pulse,” if this is preceded shortly by a prestimulus, “prepulse,” too weak to elicit a measurable startle response itself
(4,
5). The paradigms most often employed to demonstrate this effect use a strong noise burst as the pulse and a weak noise as the prepulse. The inhibitory mechanisms activated by the prepulse are thought to reduce the impact of the pulse, a powerful sensory stimulus, until the processing of the prepulse is complete and thus serve to prevent the organism from overload of information. In line with postulated deficiencies in early stages of information processing, reduced prepulse inhibition has repeatedly been demonstrated in people with schizophrenia as compared to healthy people
(6–
12).
Pharmacological agents that disrupt and enhance prepulse inhibition in experimental animals have psychotic and antipsychotic properties, respectively, in human beings. In the rat, prepulse inhibition is disrupted by dopamine agonists
(10–
12), an observation consistent with the dopamine hypothesis of schizophrenia
(13). Serotonin (5-HT) agonists and
N-methyl-
d-aspartate (NMDA) antagonists also disrupt prepulse inhibition
(10–
12); this finding is in line with the suggested involvement of these systems in the etiology and treatment of schizophrenia
(14). In addition, environmental manipulations, such as rearing in isolation, with possible relevance to neurodevelopmental models of schizophrenia
(15), are found to disrupt prepulse inhibition
(10–
12). The disruption of prepulse inhibition by dopamine agonists and social isolation is reversed by typical and atypical antipsychotics. However, atypical, but not typical, antipsychotics are able to reverse the disruption of prepulse inhibition by 5-HT agonists and NMDA antagonists
(11).
Although preclinical evidence points, as noted earlier in this article, to the differential effects of typical and atypical antipsychotics on prepulse inhibition, with a possible superiority of atypical drugs over typical ones, no published studies are available on the effects of typical and atypical antipsychotic drugs on prepulse inhibition in schizophrenic patients. A recent study
(16) reported that oral administration of the dopamine D
2 receptor agonist bromocriptine (1.25 mg) suppresses prepulse inhibition in normal male subjects, compared to placebo, and that haloperidol (3 mg) antagonizes the suppression by bromocriptine but reduces prepulse inhibition when administered on its own. We
(17) also observed that oral haloperidol (5 mg) disrupted prepulse inhibition in normal male smokers. These findings suggest that both hypo- and hyperdopaminergic states may reduce prepulse inhibition in normal human beings. However, the cognitive effects of antipsychotics are known to differ in schizophrenic patients and normal volunteers
(18). Given that prepulse inhibition deficits were noted in clinically stable, medicated (presumably with typical antipsychotics) schizophrenic patients in a number of previous studies (6–9), it seems likely that typical medication leads to, at best, a partial rather than a full restoration of underlying deficits in postulated prepulse inhibition circuitry with inputs from dopaminergic as well as from nondopaminergic systems
(11).
We therefore measured prepulse inhibition of the acoustic startle response in two groups of schizophrenic patients—those receiving clozapine and those receiving a range of typical antipsychotics—and compared them with a group of healthy subjects. Given the superiority of clozapine over typical antipsychotics in reversing prepulse inhibition deficits in the rat, we predicted greater prepulse inhibition with clozapine-treated patients than with those receiving typical antipsychotics.
METHOD
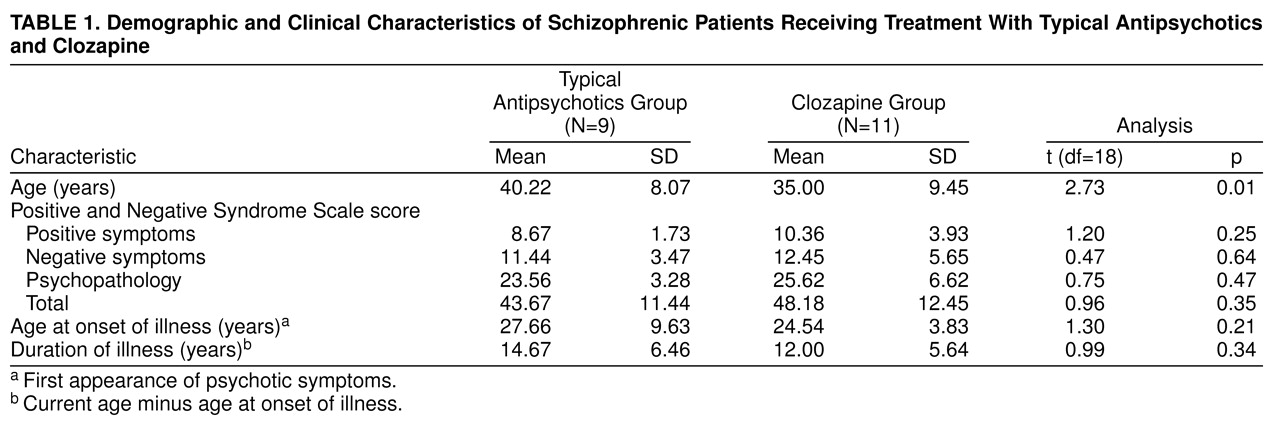
Twenty-two male patients (age range=20–65 years) with a DSM-IV diagnosis of schizophrenia or schizoaffective disorder were recruited through the inpatient and outpatient services at the Maudsley Hospital, London. Of these, 11 patients were receiving typical antipsychotics and 11, clozapine for a minimum of 6 weeks. Because of unusable eye-blink data, two patients receiving typical antipsychotics were excluded from the final analysis, thus reducing the size of this group to nine subjects. Symptoms were rated with the Positive and Negative Syndrome Scale
(19).
Table 1 presents demographic and clinical characteristics of the patients. Eleven normal volunteers (all men, age range=20–50 years; mean age=28.36, SD=6.72) were recruited through advertisement in the local newspaper and were screened for thyroid dysfunction, heart disease, hypo- and hypertension, a history of mental illness, anorexia, violent or rapid mood changes, drug abuse (ascertained by urine toxicology screen) and alcohol abuse, regular medical prescriptions, and presence of psychosis in their first-degree relatives before they were accepted as subjects. All subjects were screened for intact auditory abilities through use of an audiometer at 40 dB [A] (1000 Hz).
A commercial human startle response monitoring system (San Diego Instruments) was used to generate and deliver the startle stimuli and to record and score the electromyographic (EMG) activity for 250 msec starting from the onset of the acoustic startle stimulus. Auditory stimuli were presented to subjects binaurally through headphones. EMG recordings were taken while subjects were sitting comfortably in a moderately lit, soundproof laboratory.
The eye-blink component of the startle response was indexed by recording EMG activity of the orbicularis oculi muscle directly beneath the right eye, by positioning two miniature silver/silver chloride electrodes filled with electrolyte paste, following standard procedures
(17,
20–
22). The startle system recorded EMG activity for 250 msec (sample interval=1 msec) from the onset of the startle stimulus. Recorded EMG activity was band-pass filtered, as recommended by San Diego Instruments. A 50-Hz filter was used to eliminate the 50-Hz interference. EMG data were scored off-line, by the analytic program of this system, for response amplitude (in arbitrary analog-to-digit units) and latencies to response onset and peak (in milliseconds). Scoring criteria were identical to those reported in our previous studies
(20–
22).
The session began with a 5-minute acclimatization period consisting of 70-dB [A] continuous white noise. The pulse-alone (non-prepulse) stimulus was a 40-msec presentation of 115-dB [A] white noise, and the prepulse stimulus was a 20-msec presentation of 85-dB [A] white noise, both over 70-dB [A] continuous background noise. The pulse-alone stimulus was presented 30, 60, and 120 msec after the onset of the prepulse stimulus. Each lead interval was presented 12 times; probe without prepulse was also presented 12 times (excluding the first initial trial). There were 61 startle stimuli in all, with a mean intertrial interval of 15 seconds (range=9–23 seconds). The session lasted approximately 20 minutes.
All subjects gave written informed consent after the aim and the procedure of the study were explained to them. They were told that the purpose of the experiment was to measure their reactivity to loud noises and were told, “You are going to hear a number of auditory clicks, some of which may make you blink. Please keep your eyes open during this experiment, which will last about 20 minutes.”
All analyses were performed by SPSS (Windows, Version 6.0). Prepulse inhibition was computed as the percentage reduction of the amplitude over pulse-alone (non-prepulse) trials, i.e., prepulse inhibition=(a–b)/a × 100, where a=amplitude over pulse-alone trials and b=amplitude over prepulse trials. Such a procedure is required to correct for the influence of individual differences in startle amplitude
(23). First, the two patient groups were compared through use of between-subjects t tests for the clinical and demographic variables reported in
Table 1. Next, the effects of diagnosis (patients versus comparison subjects) and medication type (clozapine versus typical antipsychotics) on prepulse inhibition of the startle response were examined by a three- (group: patients receiving clozapine, patients receiving typical antipsychotics, comparison subjects) by-three (trial type: prepulse trials with prepulse-to-pulse intervals of 30-msec, 60-msec, and 120-msec) multivariate analysis of variance (MANOVA; Wilks’s F) with repeated measures on trial type. Further MANOVAs on prepulse inhibition scores were conducted to compare the two patient groups (separately) with comparison subjects.
DISCUSSION
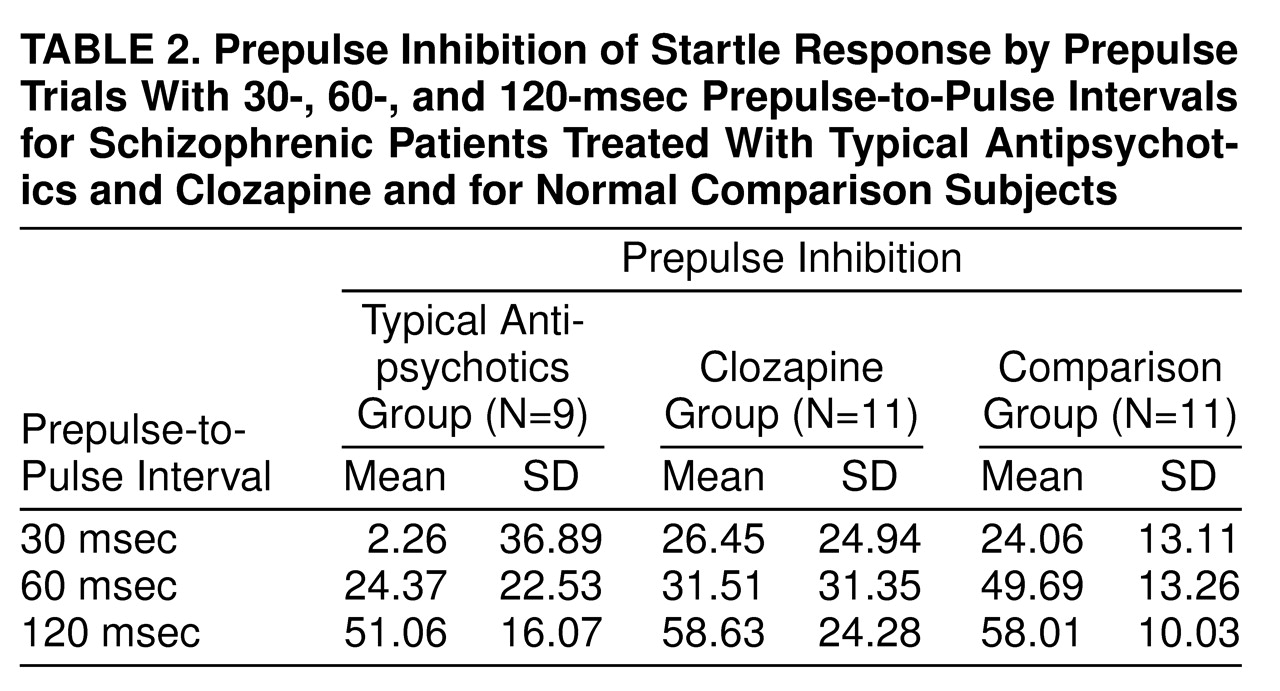
We found less prepulse inhibition in patients receiving typical antipsychotics than in comparison subjects at 30-msec and 60-msec prepulse-to-pulse lead intervals; however, they showed prepulse inhibition comparable to that of comparison subjects at the 120-msec lead interval. The finding of normal prepulse inhibition in patients receiving typical antipsychotics with 120-msec prepulse trials lends support to numerous previous findings showing positive effects of typical neuroleptic medication on controlled attention
(25). Prepulse inhibition, at least in part, is thought to reflect automatic (preattentive, mainly in prepulse inhibition with prepulses delivered at 60 msec or shorter prepulse-to-pulse intervals) processing, but it can be modulated to some degree by voluntary attention (involving conscious awareness) in human beings
(26). Active attention to the prepulses, especially at long lead intervals (>100 msec), produces an increase in prepulse inhibition
(26,
27). We used a neutral task (subjects were not instructed to pay attention to or ignore the prepulses) in order to facilitate the comparison of resulting data with those obtained from animal pharmacological studies; however, prepulses with a 120-msec prepulse-to-pulse interval are likely to have involved conscious awareness and active processing of the prepulse (although not intention, which would be required if subjects were to ignore some and pay attention to other prepulses) and thus differed from the prepulses with shorter prepulse-to-pulse intervals.
Clozapine-treated patients showed normal levels of prepulse inhibition at all prepulse intervals. This finding suggests that in schizophrenic patients, clozapine normalizes information processing functions, as assessed by prepulse inhibition of the startle response. These findings have significance for clinical and functional outcome of schizophrenia given that 1) poor cognitive functioning is related to poor functional outcome in schizophrenia
(28), and 2) impaired prepulse inhibition is positively associated with various cognitive deficits. Impaired prepulse inhibition in schizophrenia predicts poor responses on the Ego Impairment Index human experience variable, a measure of thought disorder
(29), and correlates positively with poor performance on Wisconsin Card Sort Test
(30,
31) and with distractibility on the Continuous Performance Test
(32). Impaired inhibitory processes underlying diminished prepulse inhibition in schizophrenia
(6,
11) are conceptualized to reflect an overload of sensory information, which in turn leads to cognitive fragmentation frequently seen in schizophrenia
(33). An improvement in cognitive functions with clozapine treatment would thus be expected to result in better functional outcome in schizophrenia. In line with our findings, a positive effect of clozapine has also been noted in a number of cognitive domains
(25), especially attention and verbal fluency in both treatment-resistant and non-treatment-resistant schizophrenia
(34).
The exact mechanism responsible for the superiority of clozapine over typical antipsychotics, particularly at short prepulse intervals, cannot be specified at present. This effect may be due to clozapine’s effects on prefrontal regions of the brain, especially (but not exclusively) 5-HT
2 antagonism or its limbic selectivity, or due to its actions on a range of neuroreceptors that is broader than the range affected by typical antipsychotics
(35,
36); the last possibility seems more likely given the involvement of multiple receptors in the regulation of prepulse inhibition in the rat
(11,
12). Clozapine also reduces ad lib smoking in schizophrenic patients
(37), and smoking itself is known to enhance prepulse inhibition of the acoustic startle response in normal smokers
(22). However, the mechanism responsible for the positive actions of nicotine on prepulse inhibition in the rat (38) and in human beings
(20,
21) is not yet understood. It is possible that clozapine’s effect on prepulse inhibition is mediated by the same mechanism that underlies its effect on smoking.
Although much remains to be learned about the processes underlying, and the pharmacology of, prepulse inhibition in human beings, our findings suggest that antipsychotic drugs with different pharmacological profiles would produce differential effects on automatic and controlled components of information processing, as assessed by prepulse inhibition of the startle response. It is possible that impairments in prepulse inhibition at short and long intervals reflect dysfunctions at different stages or processes of information processing and at different levels of the neurophysiological startle gating (prepulse inhibition) circuitry
(26). There are indications of such effects in both experimental animals and human beings.
Prepulse inhibition in rats is thought to be controlled by the limbic and mesolimbic-cortico-pallido-thalamic circuitry
(12). The drugs primarily acting at different neural structures in this circuitry are found to produce different patterns of prepulse inhibition disruption (or enhancement) in the rat
(39–
41). For example, apomorphine, at doses that have no effect on control animals, disrupts prepulse inhibition in rats with supersensitive nucleus accumbens dopamine receptors following intra-accumbal infusion of 6-hydroxydopamine. This effect is particularly strong for prepulse inhibition at 60-msec prepulse-to-pulse intervals, although it is also observed with prepulses delivered at 120-msec and 480-msec intervals
(40). Apomorphine’s disruptive effect on prepulse inhibition is seen in rats with 6-hydroxydopamine lesions in the substantia nigra, but most evident in this case, at prepulses delivered at 120-msec interval, with no effect at the 60-msec interval
(34). The NMDA antagonist ketamine not only disrupts prepulse inhibition at 60–500-msec prepulse-to-pulse intervals but also produces significant prepulse facilitation at the 30-msec prepulse-to-pulse interval; prepulses with a 30-msec prepulse-to-pulse interval have no effect in no-drug control conditions
(41).
In addition to schizophrenia, deficient prepulse inhibition is observed in a number of psychiatric disorders that are characterized by abnormalities in limbic and mesolimbic-cortico-striato-pallido-thalamic circuitry and exhibit deficient gating of cognitive, sensory, or motor information, such as Huntington’s disease
(42), obsessive-compulsive disorder
(43), attention deficit disorder
(44), and Tourette’s syndrome
(45,
46). However, the pattern of disruption is not uniform across these disorders. Prepulse inhibition deficits have most thoroughly been investigated in schizophrenia, and disruptions have been found across a range of prepulse intensities and intervals in this disease. Given that schizophrenic patients are known to have abnormalities at various levels of prepulse inhibition circuitry, these findings do not appear surprising. Patients with Huntington’s disease show profound disruption of acoustic prepulse inhibition across 30-msec, 60-msec, and 120-msec prepulse-to-pulse intervals
(42). The pattern of disruption in these patients is remarkably different from that in schizophrenic patients. Although schizophrenic patients show less prepulse inhibition than control subjects, they show an increase in prepulse inhibition with 30-msec to 120-msec prepulse-to-pulse intervals, with the appearance of significant prepulse inhibition at the 120-msec prepulse-to-pulse interval. Consistent with the possibility of different, but overlapping, pharmacological and neural correlates of prepulse inhibition with different prepulse-to-pulse intervals, our recent neuroimaging study
(47) revealed different patterns of brain activation during production of prepulse inhibition with 60-msec and 120-msec prepulse trials in normal human volunteers, with significant greater prefrontal activation during 60-msec prepulse condition, as compared to the 120-msec prepulse condition. The 120-msec prepulse condition elicited significantly greater activation than the 60-msec prepulse condition in the striatum (caudate nucleus), an observation that may help to explain the previous finding of a profound loss of prepulse inhibition with the 120-msec prepulse-to-pulse interval in patients with Huntington’s disease, since they are known to have substantial damage to the corpus striatum
(42). Taken together, these observations suggest that parametric manipulations may be critical in determining, with the prepulse inhibition model, the effects of pharmacological agents on information processing functions.
In conclusion, our findings suggest that clozapine is superior to typical antipsychotics in normalizing cognitive deficits in schizophrenia, at least as assessed by prepulse inhibition of the acoustic startle response. The findings also suggest that for several investigations of schizophrenic patients treated with atypical antipsychotics, the findings of normal or minimally impaired attentional and information processing functions may be attributable to the cognitive enhancing characteristics of atypical antipsychotics. The present findings support the view that prepulse inhibition is a useful animal model for future investigations of novel potential antipsychotic agents for the treatment of schizophrenia
(11) but also indicate that antipsychotics with different pharmacological profiles may produce differential prepulse inhibition profiles.
There are some limitations to this study. First, this study, as a first step, used a between-subjects design. However, this should not be considered a serious limitation because all subjects were of the same sex and had been free of substance abuse for at least 6 weeks before their participation in the study, and the two patient groups did not differ significantly in symptoms or duration of illness. Furthermore, no relationships were found between these variables and prepulse inhibition levels in general, although the small study group did not allow a meaningful evaluation of effects of symptoms. Clozapine-treated patients were younger than patients receiving typical antipsychotics, but age of subjects is known to have no relationship to prepulse inhibition
(24). The finding of normal prepulse inhibition in clozapine-treated patients is thus best explained in terms of pharmacological effects of clozapine. Another limitation of the present study is the lack of data for drug-free patients, which makes it difficult to sustain the conclusion that clozapine normalized rather than reversed deficits caused by typical antipsychotics. However, previous studies by other researchers
(48) and our preliminary data (N=2, with the paradigm reported in this study) in schizophrenic patients show that prepulse inhibition deficits are even greater in unmedicated patients than in patients receiving typical antipsychotics and that treatment with antidopaminergic drugs improves prepulse inhibition in unmedicated patients
(48,
49). Furthermore, prepulse inhibition deficits have also been noted in patients with schizotypal personality disorder, a schizophrenia-related population, the majority of whom were free from antipsychotics at the time of testing (only two of 16 patients were receiving low doses of antipsychotics)
(50). Taken together, these findings support the conclusion that clozapine is more effective than typical antipsychotics in normalizing prepulse inhibition deficits in schizophrenia.
Further clinical studies using a longitudinal, within-subjects design are required to follow up the effects of clozapine reported here and to delineate the effects of typical and a range of newer atypical antipsychotics on information processing functions in schizophrenia. Such effects can reliably, objectively, and repeatedly be measured over time in treatment-responsive and treatment-resistant schizophrenic patients and, given the similarities between the human and animal prepulse inhibition, can easily be compared with those observed in experimental animals.