The study of endophenotypes is an important strategy for understanding the biology of psychiatric disorders
(1) . A valid endophenotype represents a trait in the complex etiological pathway between susceptibility genes and clinical illness
(2) . An ideal endophenotype should be heritable, should remain stable over time, should be associated with the disorder, and should demonstrate cosegregation with the disorder in families
(1) . Many neurocognitive traits have been identified as possible endophenotypes for schizophrenia, by family studies that have shown a greater level of abnormality in the clinically unaffected relatives of individuals with schizophrenia than in normal comparison subjects
(3 –
9) . However, genetic and environmental influences are to some extent confounded in family studies. The classical twin design uses the difference in genetic sharing between monozygotic twins (100%) and dizygotic twins (50% on average) to untangle the effects of genes from the effects of family environment on any trait
(10 –
12) . The twin design, when augmented by sophisticated structural equation modeling techniques, is able to examine the genetic overlap between two traits, such as a disease and its putative endophenotype
(13 –
16) . Twin studies have been used to examine the relative magnitudes of genetic and environmental influences on brain volumes
(17 –
19) and neuropsychological measures
(20,
21) in schizophrenia.
Three event-related potentials have been proposed as candidate endophenotypes for schizophrenia: P300 (reduced amplitude and prolonged latency)
(3), P50 (reduction of inhibitory response)
(22), and mismatch negativity (reduced amplitude)
(23) . However, twin studies of event-related potentials in schizophrenia are rare. One study indicated that the P300 amplitude in both the affected and unaffected members of discordant monozygotic twin pairs was lower than in healthy comparison twins
(24) . The methods of this study did not allow estimation of either the heritability of the P300 amplitude or the extent of overlap between the genetic components for schizophrenia and for P300 amplitude.
To our knowledge, the present study is the first to use a twin design 1) to estimate the heritabilities for a range of event-related potential indices considered to be putative schizophrenia endophenotypes, 2) to quantify the strength of the relationship of each index with schizophrenia, and 3) to examine the genetic and environmental overlap with the illness. We measured mismatch negativity, P300, and P50 suppression in monozygotic twin pairs concordant and discordant for schizophrenia and in normal twin pairs. A subset of the monozygotic comparison twins were measured twice in order to assess the repeatability and the measurement error of the event-related potential indices.
Method
Participants
The study was approved by the U.K. Multi-Centre Research Ethics Committee. Probands were ascertained from U.K. national psychiatric services and the Maudsley Twin Study of Schizophrenia
(25) . Comparison twins were recruited from the Institute of Psychiatry Volunteer Twin Register and through advertisements. Written informed consent was obtained from all participants after a detailed description of the study aims and design.
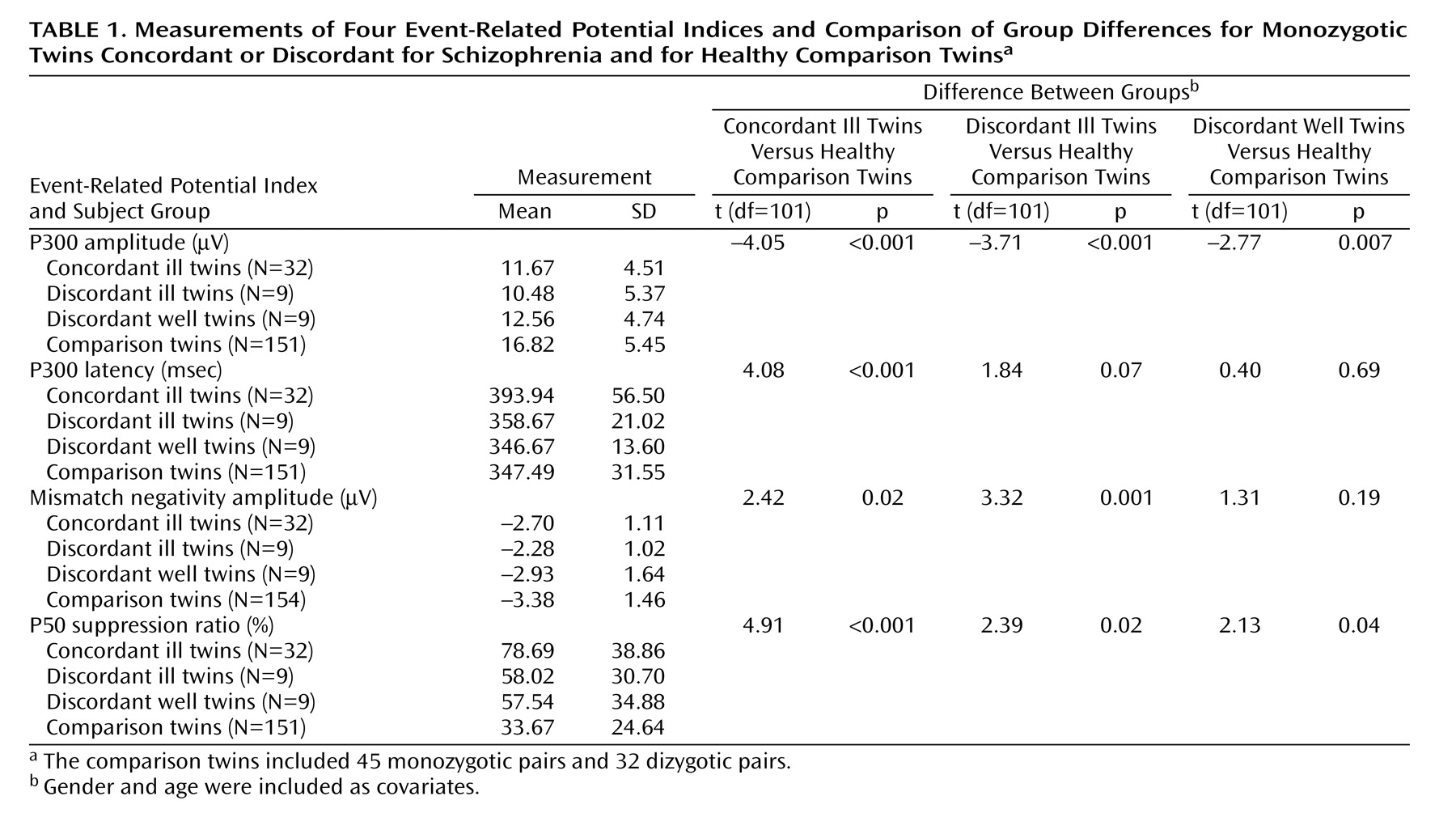
The study group consisted of 16 monozygotic twin pairs concordant for schizophrenia (mean age=41.5 years, range=23–64), nine monozygotic twin pairs discordant for schizophrenia (mean age=31.6, range=23–52), and 77 normal twin pairs: 45 monozygotic pairs (mean age=33.1, range=19–56) and 32 dizygotic pairs (mean age=40.2, range=20–58). Of those comparison twin pairs, 19 monozygotic pairs (eight male, 11 female; mean age=36.8 years) were tested on two occasions. The twin pairs concordant for schizophrenia had a higher male-to-female ratio (p<0.001, chi-square analysis), had lower parental socioeconomic status (F=4.06, df=2, 95, p=0.01), and smoked more cigarettes per day (F=7.61, df=3, 31, p<0.001) than the discordant and comparison twins, who did not differ from each other. The patients (both concordant and discordant twins) had received significantly less education than the comparison subjects (F=6.18, df=3, 206, p<0.05 in both cases).
All subjects underwent the same extensive clinical assessment. Diagnoses were based on all available clinical information, including structured clinical interviews using the Schedule for Affective Disorders and Schizophrenia—Lifetime Version (SADS-L) or the Structured Clinical Interview for DSM-IV (SCID). Additional clinical information was collected on the timing and nature of symptoms to make DSM-IV diagnoses. The exclusion criteria included a history of neurological disorder or head injury with loss of consciousness for more than 1 minute, hearing impairment, and current substance abuse or dependence. The probability that any of the discordant twins would become concordant in the future was low, as an average of 11.02 years (SD=6.49) had elapsed since the onset of the probands’ illness
(26) . The comparison subjects were free of a personal or family history, to second-degree relatives, of psychotic spectrum disorder.
All but two of the patients were taking antipsychotic medication at the time of assessment; 11 were also treated with antidepressant medication. The incidence of lifetime substance and alcohol problems was significantly higher in the concordant group (F=3.17, df=3, 204, p=0.01, logistic regression analysis) than in the comparison group. Three patients (two from the concordant group) and one nonschizophrenic co-twin had lifetime diagnoses of substance abuse (mainly for cannabis), while one nonschizophrenic co-twin had had cannabis abuse and alcohol dependence in the past. One patient from a discordant pair had a history of substance abuse and alcohol dependence. One patient from a concordant pair had a history of substance and alcohol abuse, while two patients from the concordant group had alcohol abuse histories. All subjects had been free of any substance misuse for at least 2 years at the time of testing. All patients were clinically stable at the time of assessment, with no recent changes to their medication. Over one-half (56%) of the unaffected co-twins had a history of major depression, and 15 of the comparison twins satisfied the DSM-IV criteria for a lifetime axis I disorder (mainly major depression). None was unwell at the time of assessment or was taking psychotropic medication. Zygosity was determined by using 12 highly polymorphic DNA markers and a standardized twin-likeness questionnaire.
Procedure and Tasks
Three separate recordings for P50, mismatch negativity, and P300 were carried out in a fixed order by means of methods described in detail elsewhere
(27) . Briefly, data were collected by using Neuroscan software (Scan 4.3, Compumedics, Hamburg, Germany). EEG data were recorded according to the 10/20 international system, referenced to the left ear. Eye movements were recorded from the outer canthus of each eye, above and below the left eye. Electrode impedances were below 6 kΩ. EEG activity was amplified 10,000 times with 0.03-Hz high-pass and 120-Hz low-pass filters and was digitized at 500 Hz. The subjects had last smoked no less than 40 minutes before data collection
(28) .
P300
P300 was assessed by using an auditory oddball task (400 binaural tones at 80 dB with a 20-msec duration); 20% were target stimuli (1500 Hz), and 80% were standard tones (1000 Hz). The participant pressed a button in response to a target tone. The EEG data were segmented into epochs (–100 to 800 msec), digitally filtered (0.15–40 Hz), low-pass filtered (8.5 Hz), and corrected for baseline values. Eye-blink artifacts were corrected by using regression-based weighting coefficients (29). Epochs were rejected if amplitudes exceeded 50 μV at the F7, F8, Fp1, or Fp2 site or if residual horizontal eye movements were present between –100 and 800 msec. Separate average waves for target and standard tones were calculated and were measured at the Pz site between 280 and 600 msec.
Mismatch Negativity
Mismatch negativity was elicited by an auditory oddball-duration task using four blocks of 400 binaural 80-dB stimuli (interstimulus interval=0.3 sec) with 85% standards (25 msec, 1000 Hz, 5-msec rise/fall time) and 15% deviants (50 msec). The EEG data were segmented into epochs (–100 to 300 msec), filtered (0.1–30 Hz), and corrected for baseline values. Epochs were rejected if the amplitudes exceeded 100 μV in any channel. Eye-blink artifacts were corrected as already described. Mismatch negativity was extracted by subtracting the averaged waveforms for the standard stimuli from those for the deviant stimuli. The amplitude of the mismatch negativity was measured at Fz between 50 and 200 msec.
P50 Suppression
P50 waves were recorded with a conditioning-testing paradigm. Condition and test clicks were of 1-msec duration and separated by 500 msec. Intertrial intervals were 10 seconds. The participants were presented with four or five blocks of 30 pairs of conditioning and test clicks. The blocks were separated by 1-minute breaks. Stimulus intensity was adjusted individually to 43 dB above the hearing threshold. EEG signals were segmented into epochs (–100 to 400 msec), filtered (1-Hz high-pass filter), and corrected for baseline values. Epochs with activity exceeding 20 μV in the Cz or electro-oculography channel between 0 and 75 msec poststimulus were automatically rejected. Epochs were averaged separately for the condition and test waveforms, digitally filtered (10-Hz high-pass filter), and smoothed (by using a 7-point moving average applied twice). P50 event-related potentials are reported at the Cz site. For the conditioning response (C), the most prominent peak 40–75 msec poststimulus was selected as the P50 peak. The preceding negative trough was used to calculate the amplitude. For the test response (T), the positive peak with the latency closest to that of the conditioning P50 peak was selected, and its amplitude was determined as for the conditioning wave. The P50 suppression ratio was calculated as (T/C)×100.
Statistical Analyses
Comparison of means
For each event-related potential measure, we compared the means of the following groups: monozygotic twins concordant for schizophrenia, affected twins in monozygotic discordant pairs, unaffected twins in monozygotic discordant pairs, and healthy comparison twins. An observed impairment in the unaffected monozygotic discordant twins similar to that of their affected co-twins would suggest that the impairment in the event-related potential is caused by the genetic component of schizophrenia, whereas values intermediate between those of the patients and healthy comparison subjects would indicate that the deficit is caused by illness progression as well as the genetic predisposition to schizophrenia. The comparisons of mean values were analyzed by means of a regression command in STATA (Stata Corp., College Station, Tex.) that allows for nonindependent observations (e.g., twin pairs) by using a robust sandwich estimator to estimate standard errors. Gender and age were included as covariates.
Statistical modeling of the data
A more sophisticated approach to the analysis of twin data is the method of structural equation models, which aim to explain the pattern of correlations in the data by a linear model of relationships between latent and observed variables
[30] . For the present study, however, model fitting was complicated by a number of factors: 1) the multivariate nature of the data, involving schizophrenia and multiple event-related potential measures, 2) repeated measurements for the monozygotic comparison twin group, 3) the dichotomous nature of schizophrenia, 4) the uncertain ascertainment process for monozygotic twins concordant and discordant for schizophrenia, and 5) the possibility that the test for common environment is seriously underpowered (because of the absence of dizygotic twin pairs concordant and discordant for schizophrenia in the study design). The analytic strategies we used to overcome these difficulties are outlined as follows.
1. To accommodate the multivariate nature of the data, we used multivariate models that consider the patterns of covariances between multiple variables both within individuals and across twins
(31) . Also, we considered each of the event-related potential paradigms separately in the model-fitting analyses as we found no evidence of a genetic overlap among P300, P50 suppression, and mismatch negativity
(32) .
2. We dealt with the repeated measurements for the monozygotic comparison twin group by specifying additional observed variables in the covariance model for the comparison pairs that were retested.
3. To accommodate the dichotomous nature of schizophrenia, we used liability-threshold models for both schizophrenia and the event-related potential variables
(33) . For schizophrenia this model assumes that risk is normally distributed on a continuum and that the disorder occurs only when a certain threshold is exceeded
(16,
33) . Since Mx software
(30) does not allow simultaneous analyses of dichotomized and continuous data, the event-related potential variables were recoded into seven equal ordinal classes, which should capture most of the information in the continuous data.
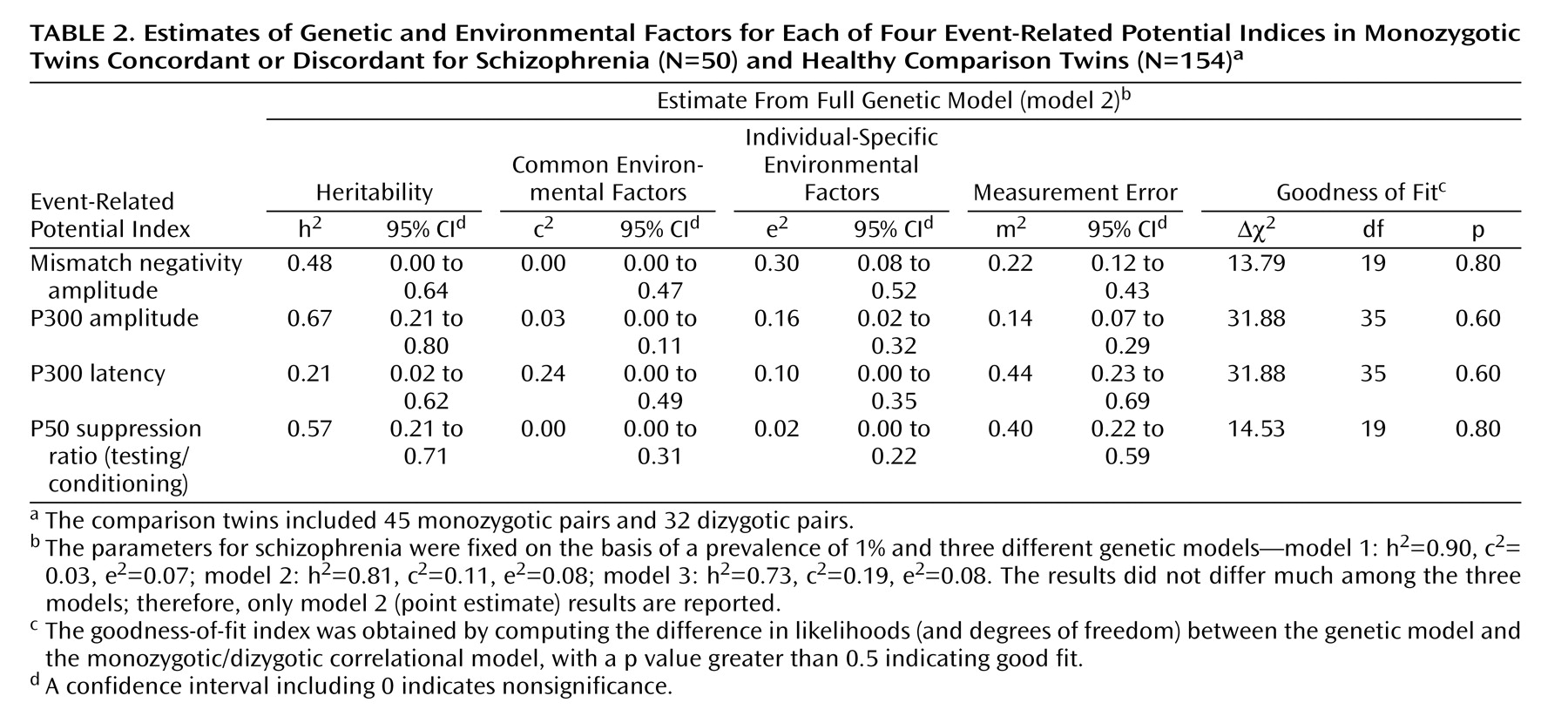
4. The uncertain ascertainment process for monozygotic twins concordant and discordant for schizophrenia was overcome by fixing the model parameters for schizophrenia to the population values, since the selection was based on schizophrenia and was blind to values for the event-related potentials
(16) . The model parameters for schizophrenia were fixed to three sets of values based on those from a meta-analysis report
(34) : the point estimate (model 2: h
2 =0.81, c
2 =0.11, e
2 =0.08), the lower 95% confidence limit (model 1: h
2 =0.73, c
2 =0.19, e
2 =0.08), and the upper 95% confidence limit (model 3: h
2 =0.90, c
2 =0.03, e
2 =0.07) (h
2, c
2, and e
2 represent estimates of heritability, common environmental factors, and individual-specific environmental factors, respectively). The prevalence rate of schizophrenia was fixed to 1%.
5. Because of the possibility that the test for common environment was underpowered, the shared genetic and common environmental effects in schizophrenia were tested separately by using two models: one fixing the correlational path for common environment (c′ 1 ) to zero (the genetics-only model) and one fixing the genetic correlational path (a′ 1 ) to zero (the model based on common environment only). Since these models are not nested, we used the Akaike information criterion to determine the best-fitting model.
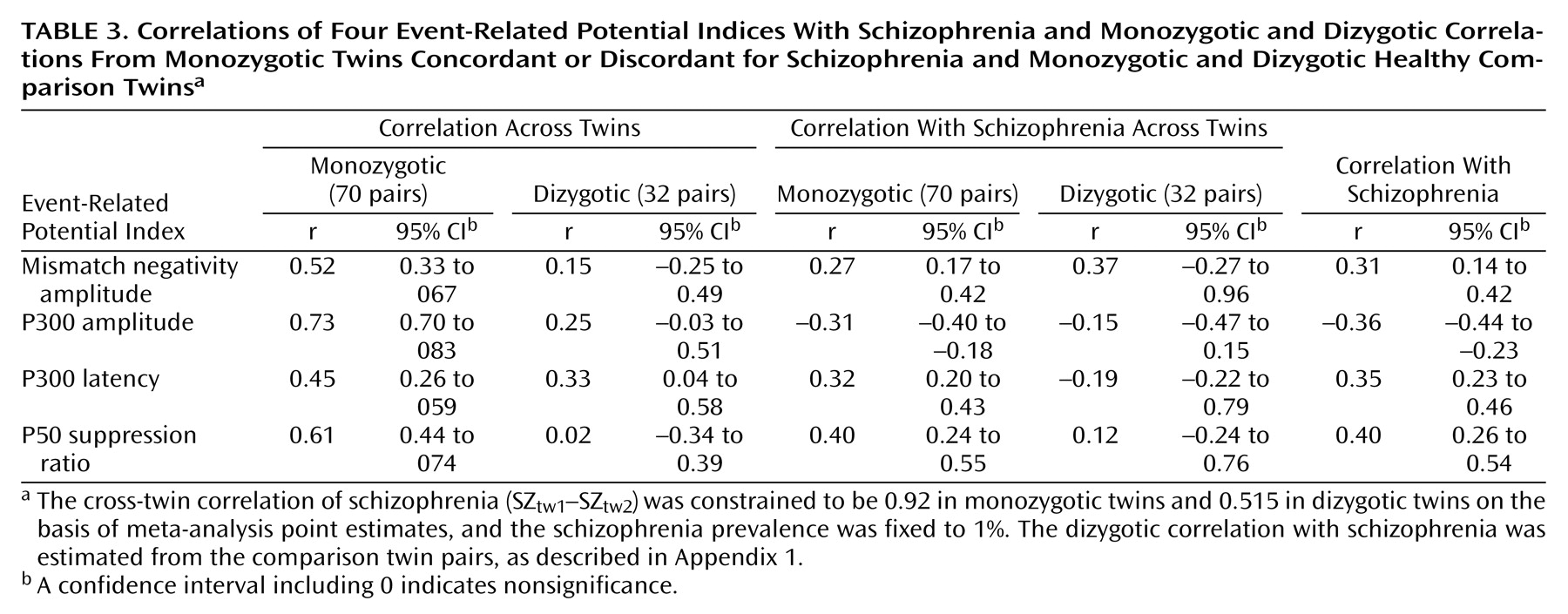
Twin correlations
Twin correlations between the event-related potential measures and schizophrenia were estimated by fitting a correlation model to the corresponding observed raw data for monozygotic and dizygotic twins.
The correlation model for each combination of schizophrenia status and event-related potential was constrained to produce 1) the same cross-trait correlation for all individuals, 2) the same monozygotic cross-twin within-trait correlations regardless of occasion, 3) the same monozygotic cross-twin cross-trait correlations regardless of twin order and occasion, and 4) the same dizygotic cross-twin cross-trait correlations regardless of twin order. In addition, for schizophrenia, the monozygotic (MZ) and dizygotic (DZ) cross-twin correlations were fixed according to the point estimates from the meta-analysis
(34) (i.e., r
MZ =0.92 and r
DZ =0.515) and the threshold to a prevalence of 1%. The dizygotic correlation with schizophrenia was estimated from the comparison twin pairs (as described in
Appendix 1 ).
Genetic model fitting
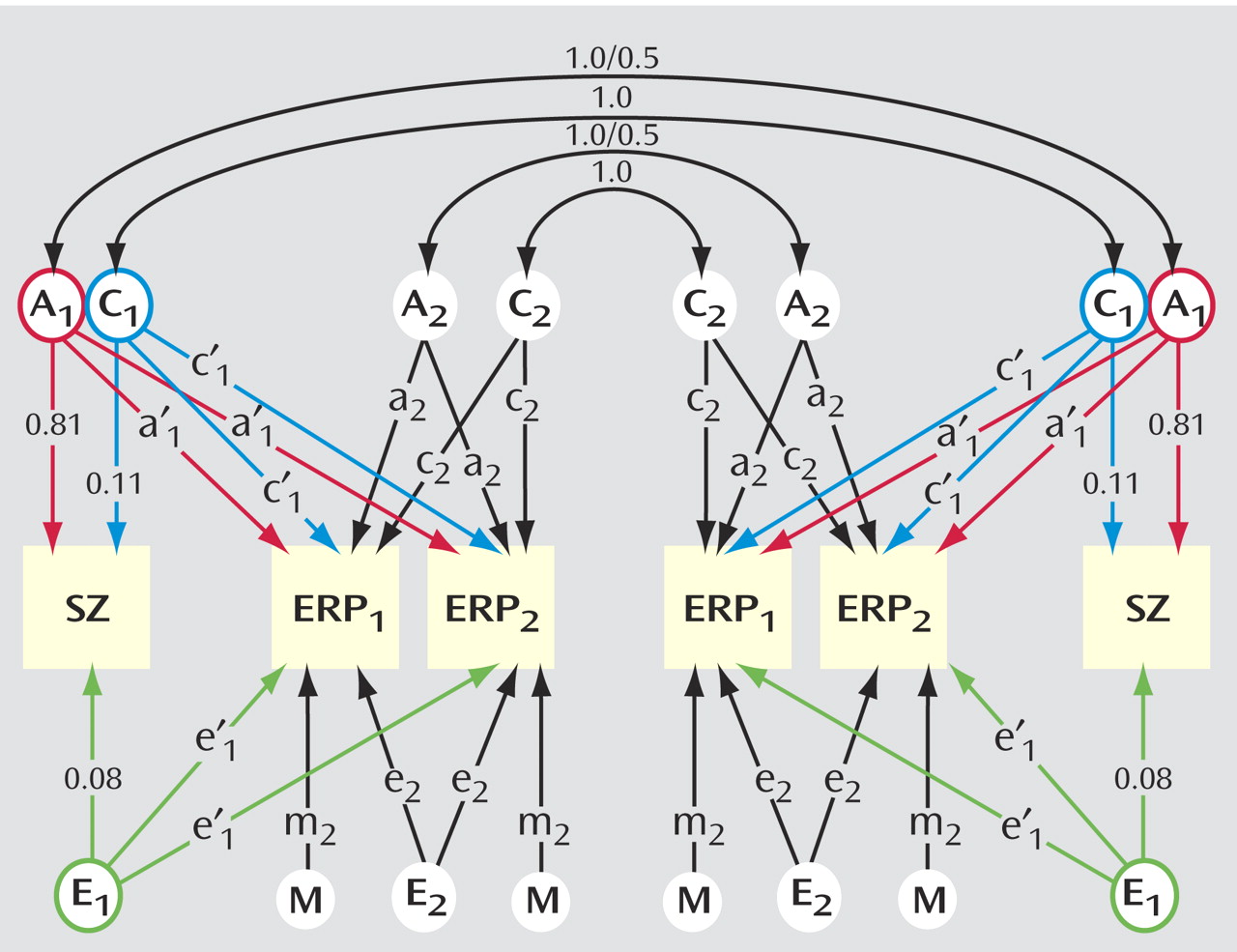
Genetic model fitting was applied to estimate, for each event-related potential variable, 1) heritability, 2) genetic and environmental correlations with schizophrenia, and 3) measurement error (due to the repeated measurements of the monozygotic comparison group). The full model for monozygotic pairs (used for P50 and mismatch negativity) is illustrated in
Figure 1 . There are three common factors—additive genetic (A
1 ), shared environment (C
1 ), and individual-specific environment (E
1 )—influencing both schizophrenia (paths a
1, c
1, e
1 ) and the event-related potential (paths a′
1, c′
1, e′
1 ), whereas measurement error is among the four specific factors (A
2, C
2, E
2, and M
2 ) that influence the event-related potential (paths a
2, c
2, e
2, m
2 ). The parameters were constrained to be equal across measurement occasions. This standard multivariate genetic model partitions the phenotypic correlation between schizophrenia and the event-related potential into components related to genetic influences, common environment, and individual-specific environment, by considering monozygotic and dizygotic correlations across traits and twins
(36), that is, the correlation between one twin’s liability to schizophrenia and the co-twin’s value for the event-related potential. For example, significantly greater cross-trait cross-twin correlations in monozygotic than in dizygotic pairs would suggest a genetic contribution to the correlation between schizophrenia and the event-related potential. The parameter estimates from the model can be used to derive the correlation between the genetic factors of schizophrenia and the event-related potential (R
g ) and can be used similarly for the correlations of common environmental (R
c ) and individual-specific environmental (R
e ) factors.
The liability-threshold model for schizophrenia and the P300 measure was analyzed in a similar way, except that the model included both the amplitude and latency.
Models were fitted directly to the raw data. In this case, a goodness-of-fit index (chi-square value) was obtained by computing the difference in likelihoods (and degrees of freedom) between the genetic models and the correlational model. Submodels of the full ACEM model were evaluated by comparing the difference in chi-square values relative to the difference in degrees of freedom, according to the principles of parsimony, operationalized by the significance of the difference in chi-square.
Discussion
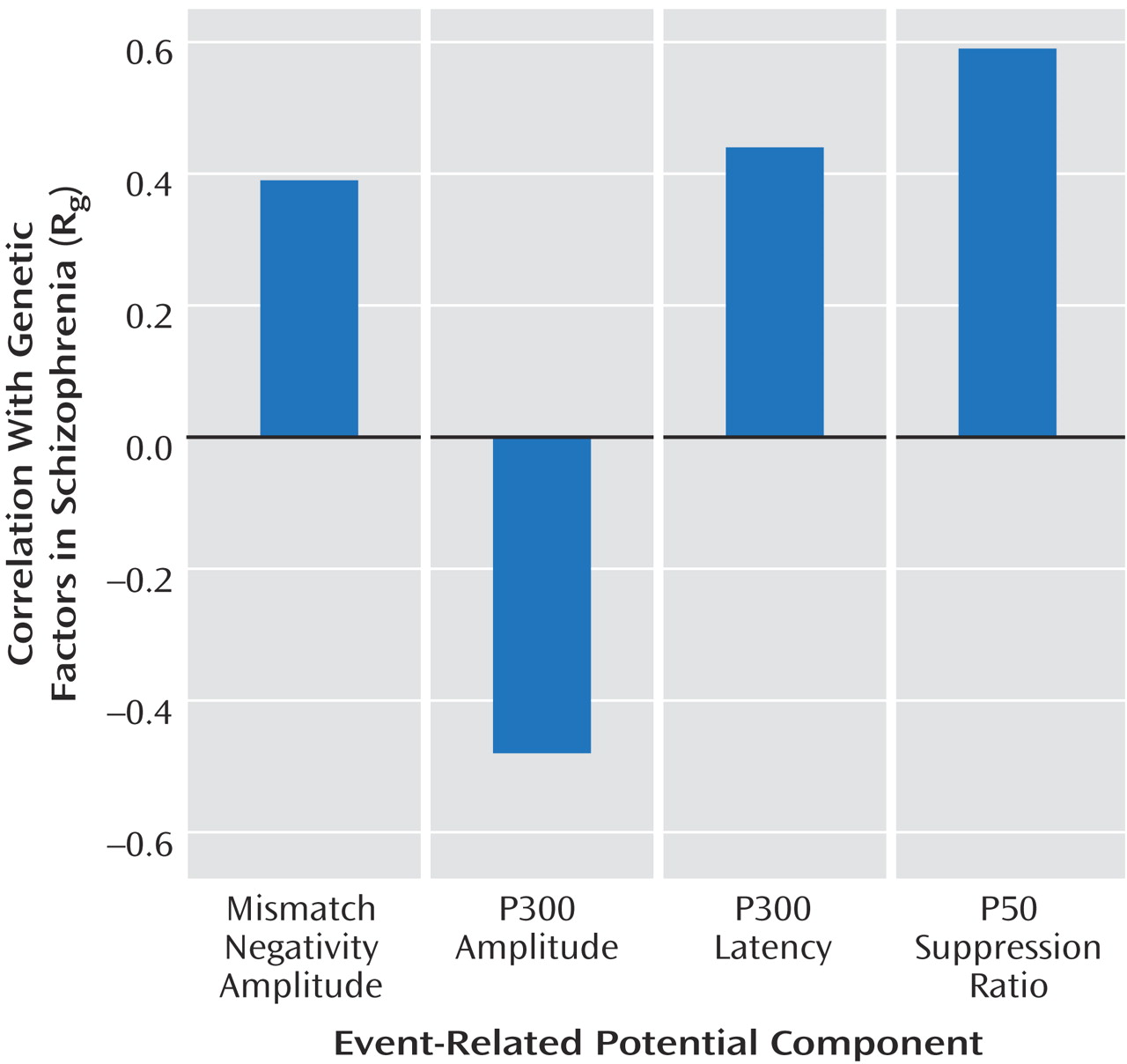
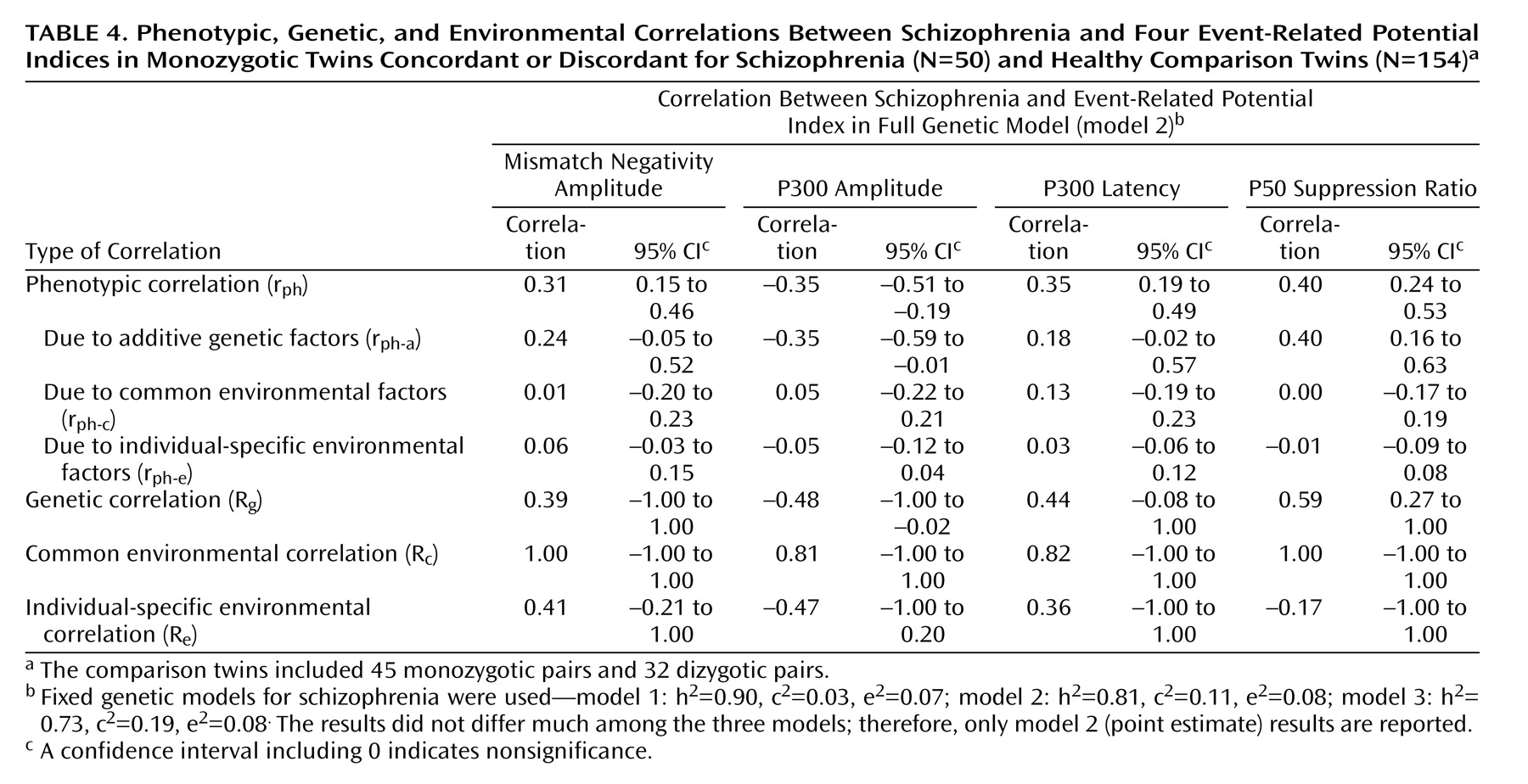
We detected significant phenotypic correlations between schizophrenia and each of the event-related potential measures across all three schizophrenia models. Genetic factors were the main source of the phenotypic correlations. The genetic correlations (R g ) were highest for the P50 suppression ratio, followed by the P300 amplitude and latency, and lowest for the mismatch negativity amplitude. Environmental correlations (R c and R e ) for each variable were nonsignificant.
Our study, including schizophrenic twins, gave estimates of the heritability of these event-related potential indices that are comparable to those previously reported for healthy twins
(27,
35) . Also, the present findings are consistent with the results of meta-analyses of P300, P50, and mismatch negativity that support an association between schizophrenia and deficits in these measures
(3,
22,
37) .
The results of the present study suggest that the P50 ratio is an attractive event-related potential endophenotype for schizophrenia for several reasons: 1) it has little environmental variance, except measurement error, 2) it has the highest phenotypic correlation with schizophrenia, 3) almost all of the phenotypic correlation with schizophrenia is explained by shared genetic factors, and 4) its genetic correlation with schizophrenia was robust across the three sets of schizophrenia models.
P50 suppression provides a measure of sensory inhibition in the brain and reflects the individual’s ability to filter out repetitive stimuli in order to minimize information overload. Impaired P50 inhibitory responses have been observed in schizophrenic patients and their unaffected relatives and have been linked to the alpha-7 nicotinic receptor gene (CHRNA7)
(38) . Our results provide supporting evidence that there is a genetic relationship between schizophrenia and P50 suppression.
The P300 explores selective and sustained attention as well as working memory. The amplitude of the wave is thought to be proportional to the amount of attentional resources devoted to the task
(39) . Its latency reflects stimuli classification speed and reaction time
(40) . Our results also support the use of P300 amplitude as an endophenotype. The phenotypic correlation between P300 amplitude and schizophrenia was –0.35, of which 75% was explained by genetic effects. Also, a high heritability estimate and low measurement error are favorable features for a good endophenotype. However, for the latency, although a substantial phenotypic correlation was found between latency and schizophrenia (0.35), the heritability estimate was relatively low, and the significance of the genetic correlation with schizophrenia depended on using a model with the highest heritability estimate for schizophrenia.
In response to a repetitive auditory stimulus, the brain automatically develops an accurate neuronal trace or “echoic memory” that represents the physical features of the stimulus. When a new input does not match the trace, the mismatch generator process is activated
(41) . The mismatch negativity may form part of an alert that serves as a survival mechanism by detecting unusual and possibly dangerous events in the environment
(42) .
Duration mismatch negativity amplitude appears to be a weak endophenotype for schizophrenia. The phenotypic correlation between mismatch negativity and schizophrenia is weak, and a significant genetic correlation (R
g =0.38) could be detected only when the highest schizophrenia heritability estimate was applied. In addition, the significant environmental component also weakens its usefulness as a putative endophenotype. Mismatch negativity is reported to be specifically impaired in chronically ill schizophrenic patients but is normal in first-episode psychotic patients
(43) . Thus, mismatch negativity may not be as robust as the other two measures. It remains to be determined whether the deficits in mismatch negativity are environmental or secondary to the disease process or, instead, reflect a late genetically determined abnormality.
Contrasting the results from the comparisons of means and the genetic model fitting reveals two interesting points: 1) interpretation based only on the results of comparing mean values might lead one to conclude that the P300 amplitude is a better endophenotype for schizophrenia than the P50 ratio, and 2) few or no genetic effects were found in the comparison of mean values for P300 latency and mismatch negativity amplitude, whereas genetic effects were picked up from using genetic model fitting. These somewhat disparate results between the two approaches may have occurred because comparisons of means do not utilize all available information. In contrast, statistical modeling uses all the available information to address directly the parameters of interest in a likelihood framework
(10,
44) . Nevertheless, it is reassuring when the results of the two approaches are largely consistent with each other.
There is some evidence that typical and atypical antipsychotic medications affect P300
(45) but not mismatch negativity
(46) in schizophrenic patients; Clozapine but not other atypical or typical antipsychotics may normalize P50 suppression deficits in patients
(47) . In our study group, 10 patients were taking clozapine (seven from concordant twin pairs, three from discordant pairs), and their averaged suppression ratio (mean=59.46, SD=24.60) was not significantly different from that for the rest of the patients (p=0.28, two-tailed t test). In addition, there was no difference in event-related potential responses between the probands and their unaffected, unmedicated co-twins, suggesting that clozapine was unlikely to contribute to the differences observed in our data.
Our findings are subject to several limitations. First, the study group was small. Our power analyses showed that with this group we could detect only a genetic correlation between schizophrenia and an event-related potential of 0.4 when the heritability of the event-related potential was 50%. Second, the study design was underpowered in detecting common environmental effects, partly because of the absence of dizygotic twins with schizophrenia, but any common environmental effects cannot be greater than the common environmental effects for schizophrenia, which are small. Third, the measurement of each event-related potential was based on one recording location. Therefore, it was not possible to examine topographical differences between patients and comparison subjects.
The strengths of this study include the use of a model-fitting method to quantify the genetic overlaps with schizophrenia and allow us to compare the magnitudes of these overlaps across a range of event-related potential indices.
The presence of substantial genetic influences on schizophrenia and event-related potentials suggests that research into the neurochemical mechanisms of abnormalities in event-related potentials may illuminate the pathophysiology of schizophrenia. In addition, if such abnormalities in unaffected relatives represent the expression of genes relevant to the etiology of schizophrenia, then including these phenotypes in genetic analyses could improve the power to detect susceptibility genes through either linkage or association
(48) .