Anorexia nervosa has been shown to run in families
(1), and one twin study, using unsystematic ascertainment, has shown substantially higher concordance rates in monozygotic than in dizygotic twin pairs
(2). We previously reported on the only population-based twin sample examined for anorexia nervosa
(3). Although we could demonstrate significant familial aggregation, the narrow criteria utilized produced a sample too small for accurate statistical modeling.
Both clinical and population-based studies of women have consistently revealed an increased association between major depression and anorexia nervosa. In population-based studies, lifetime major depression has been reported in about 50% of the women with anorexia nervosa
(4). In addition, the rates of major depression are elevated in the first-degree relatives of women with anorexia nervosa
(1). The origin of the comorbidity of anorexia nervosa and major depression has been debated in the literature
(5). Some authors contend that anorexia nervosa and major depression share a common etiology
(6), and others suggest that the liability for anorexia nervosa is distinct from that operating in the transmission of affective disorders
(7).
In this report, using a broader phenotypic definition of anorexia nervosa than we used previously
(3), we attempt to illuminate the most accurate means of determining individual variation in anorexia nervosa and to clarify the model for the comorbidity of lifetime anorexia nervosa and major depression.
METHOD
Our data are from a population-based longitudinal study of Caucasian female twins drawn from the Virginia Twin Registry, a population described by us previously
(3). Data used in the present study are from the first interview wave (1987–1989), which consisted of interviews with 2,163 women, including 1,030 complete female twin pairs of known zygosity (597 monozygotic and 433 dizygotic). The mean age of the twins was 29.3 years (SD=7.7). Written informed consent was obtained before face-to-face interviews, and verbal assent was given before telephone interviews. Zygosity was determined blindly by standard questions and photographs; recent revalidation of zygosity assignment by means of highly polymorphic polymerase chain reactions showed an error rate of 4.5%.
Interviewers were blind to any information about the co-twins. To maximize the statistical power in this study of a disorder with a low prevalence, a broad definition of lifetime anorexia nervosa was adopted. A previous examination of this population suggested an etiologic continuity between narrowly and broadly defined anorexic symptoms
(3). Individuals were considered to have lifetime anorexia nervosa if one or more of the following definitions were true: 1) meets strict DSM-III-R criteria, 2) meets DSM-III-R criteria minus criterion D (amenorrhea), and 3) meets DSM-III-R criteria minus criterion C (feeling fat when emaciated). Lifetime major depression was also diagnosed according to DSM-III-R criteria. Both disorders were assessed by using the Structured Clinical Interview for DSM-III-R
(8).
We used the Mx statistical program (9) to perform bivariate Cholesky decompositions (10) with the input data (in the form of tetrachoric correlation matrices) and the associated asymptotic weighted least-squares matrices. In the bivariate twin model, the correlation between two phenotypes can be divided into the correlations due to genetic, common environmental, and unique environmental effects.
RESULTS
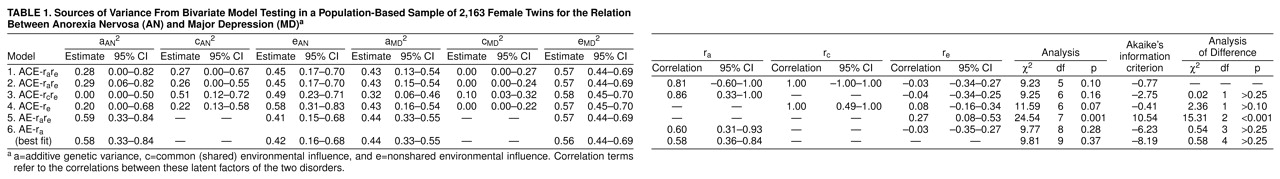
A total of 77 women (3.6%) met our expanded criteria for anorexia nervosa—11 (0.5%), 29 (1.3%), and 37 (1.7%) for each of our three definitions, respectively. Of these, 18 monozygotic and 20 dizygotic female co-twins (N=38, 49.4%) also had a lifetime diagnosis of major depression. Four monozygotic and two dizygotic twin pairs were concordant for anorexia nervosa; 86 monozygotic and 60 dizygotic pairs were concordant for major depression.
In addition to testing the full model in the bivariate model fitting, we tested 10 nested submodels, five of which are shown in
table 1. Of the five additional submodels tested (not shown), none provided a better fit for the data. The best-fitting model, as determined by Akaike’s information criterion values, was model 6, which included genetic and unique environmental effects for the development of both anorexia nervosa and major depression and for the genetic correlation between the two disorders (0.58). This model indicated that the proportion of shared genetic variance between anorexia nervosa and major depression is 34% (95% confidence interval [CI]=13%–71%).
DISCUSSION
The rarity of anorexia nervosa has previously precluded biometrical model fitting in twin studies
(3). By expanding the definition of anorexia nervosa—which is supported by the observation that anorexia nervosa and anorexia nervosa-like syndromes appear to lie on a continuum
(3,
7)—and by conducting bivariate twin analyses on a disorder with a higher prevalence in the total population (major depression) than that of anorexia nervosa, we were able to derive useful estimates of the genetic and environmental contributions to the liability for anorexia nervosa.
The best-fitting model for the expanded phenotype of anorexia nervosa suggests substantial heritability (58%). Noteworthy, however, are the wide 95% CIs indicating that our estimate is imprecise. This suggests that we consider equally the full and best-fitting models in interpreting the results. While the best-fitting model suggests that the environmental contribution to the syndrome of anorexia nervosa consists of nonshared environment only, the full model suggests that shared environment in twins may also contribute to the development of anorexia nervosa.
In the best-fitting model, the genetic liabilities for anorexia nervosa and major depression are correlated, which suggests a modest overlap in the genetic liability that predisposes an individual to both disorders (34%). Because the correlation found was less than unity, separate genes appear to contribute uniquely to the liability for anorexia nervosa and major depression. No evidence was found for correlated environmental risk factors for the development of anorexia nervosa and major depression. These results indicate that there are both shared and unique genetic effects in the etiology of the two disorders.
The interpretation of these results is potentially ambiguous. While the monozygotic and dizygotic correlations for anorexia nervosa are consistent with the effect of shared environment, there are also substantial cross-twin, cross-trait correlations between major depression and anorexia nervosa. Since twin resemblance for major depression is due solely to genetic factors, these correlations strongly suggest that genetic factors also influence the risk for anorexia nervosa. Given that the causes of major depression can be observed with much greater precision than can the causes of anorexia nervosa, the results of the model fitting give greater weight to the cross-twin correlations between major depression than anorexia nervosa than to the twin correlations for anorexia nervosa itself.