With an incidence of 1 in 800 live births, Down’s syndrome is the most common genetic cause of mental retardation
(1). Down’s syndrome provides a rare opportunity to explore relationships among genetic, structural, and cognitive or developmental abnormalities. In 95% of cases, Down’s syndrome is caused by a full trisomy of chromosome 21
(2). Individuals with Down’s syndrome typically are microcephalic and have characteristic facies, hypotonia, and small stature. Although this physical phenotype is easily recognized, mental retardation of varying degrees is the most consistent feature of Down’s syndrome
(3). Cognitive development in Down’s syndrome has been extensively studied, yielding a profile of global delays with disproportionately impaired speech and language
(4). Some studies
(5–
7) also have identified major deficits in both short-term and long-term verbal memory. It is of interest that, compared to the dramatic impairment in these areas, visuospatial processing skills appear to be relatively preserved in individuals with Down’s syndrome
(8,
9).
Until recently, our understanding of the structural brain abnormalities in Down’s syndrome was almost exclusively based on autopsy studies. These have consistently shown a lower brain weight and brachycephaly, with a small cerebellum, frontal and temporal lobes, a simplified appearance of the sulci, and a narrow superior temporal gyrus
(3,
10,
11). Only in the past several years have improvements in magnetic resonance imaging (MRI) and image processing techniques allowed quantitative explorations of brain structure in living subjects with Down’s syndrome. Consistent with autopsy reports, volumetric neuroimaging studies of adults with Down’s syndrome
(12–
16) have revealed smaller overall brain volumes, with disproportionately smaller cerebellar, brainstem, frontal lobe, and hippocampal volumes. Basal ganglia volumes, however, have been reported to be normal in MRI volumetric studies of adults with Down’s syndrome
(14,
17). Considering the high prevalence of Down’s syndrome, surprisingly few MRI studies of affected children have been published. Jernigan et al.
(18) reported similarly smaller overall brain volumes, with disproportionately smaller volumes in frontal, temporal, and cerebellar regions, in a volumetric MRI study of six children with Down’s syndrome. As in the adult studies, volumes of thalamus and lenticular nuclei were noted to be normal. Caution must be exercised in interpreting these studies of brain structure in subjects with Down’s syndrome, as most have employed small numbers of subjects and much of the volumetric data were obtained with relatively low resolution image acquisition techniques (e.g., 5-mm brain slices with between-slice gaps of 2.5 mm), compared to the techniques now available and used in the present study, which allow 1–2-mm resolution and no between-slice gap
(12,
13,
18).
The goal of the present study was to obtain more precise quantitative neuroimaging data in children and young adults with Down’s syndrome and to determine whether the findings of previous imaging studies could be confirmed using newer high-resolution MRI acquisition techniques and an advanced segmentation and image processing protocol. Since we know of no prior studies that include children with Down’s syndrome under age 10, we also sought to determine whether measurable differences are present from earlier in childhood. On the basis of the results of prior neuroimaging studies, we hypothesized smaller overall brain volumes, with disproportionately smaller cerebellar and frontal volumes and selective preservation of subcortical structures. The pattern of deficits in language and memory, combined with evidence for aberrant language localization
(19–
21),
led us to explore the additional a priori hypothesis that the temporal lobes and, specifically, the superior temporal gyri would be significantly smaller in children with Down’s syndrome and would show an abnormal pattern of asymmetry. Additionally, the relative strengths in visuospatial skills in children with Down’s syndrome pointed to the parietal lobe as a potential region of interest
(5,
6).
Discussion
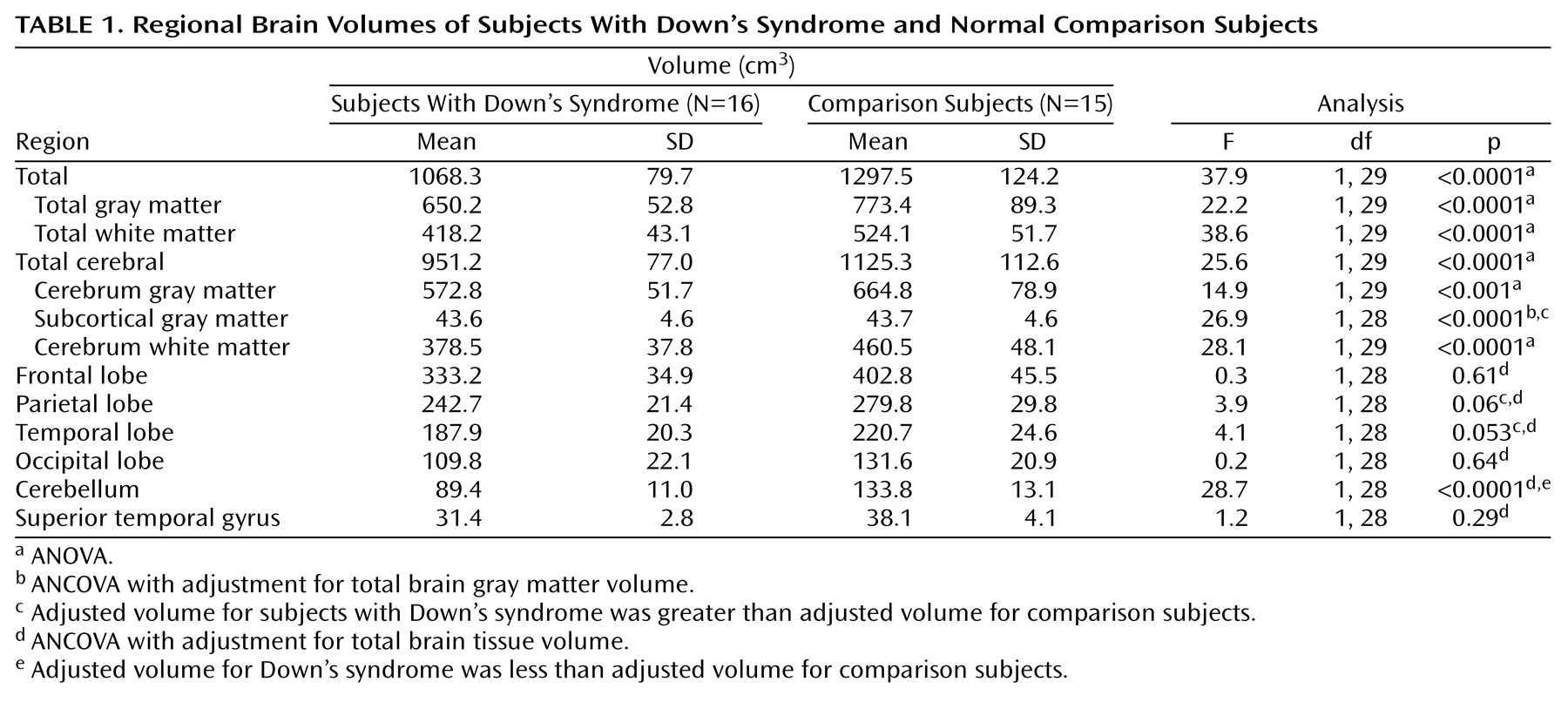
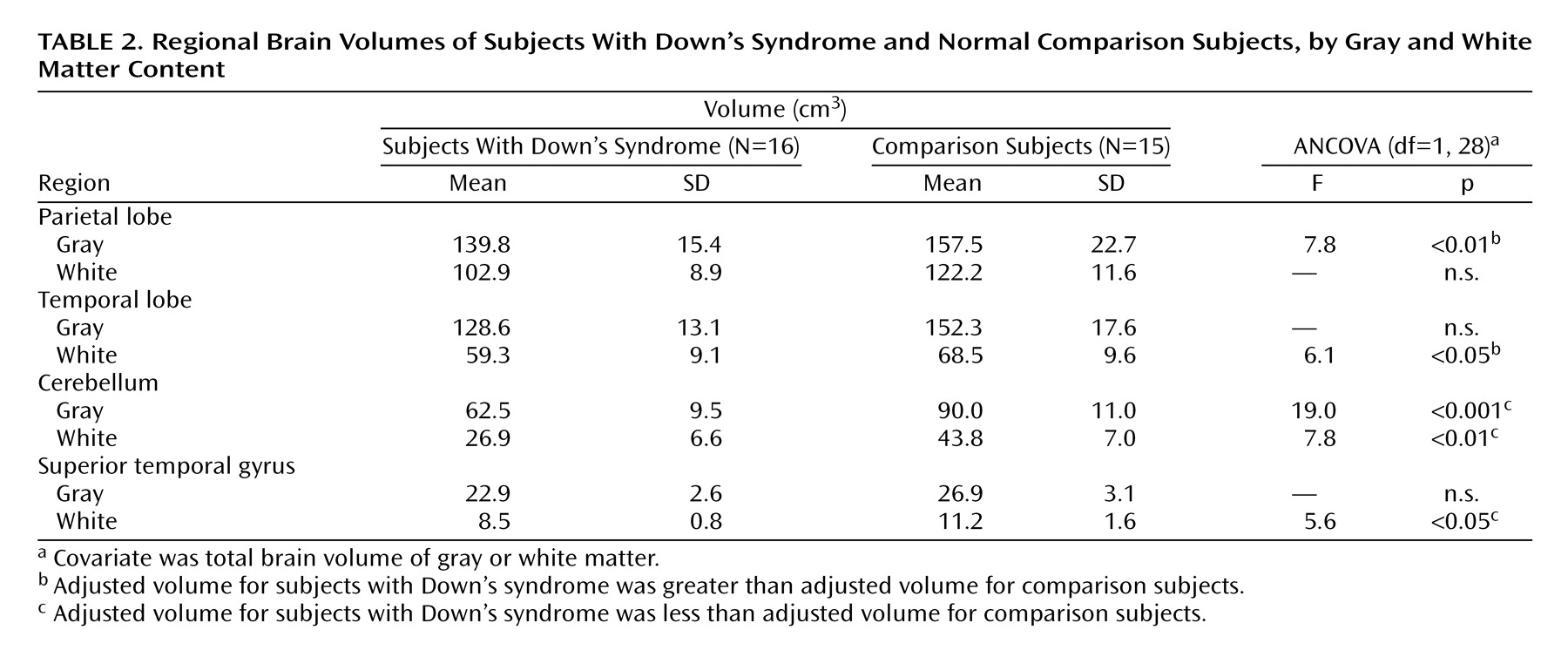
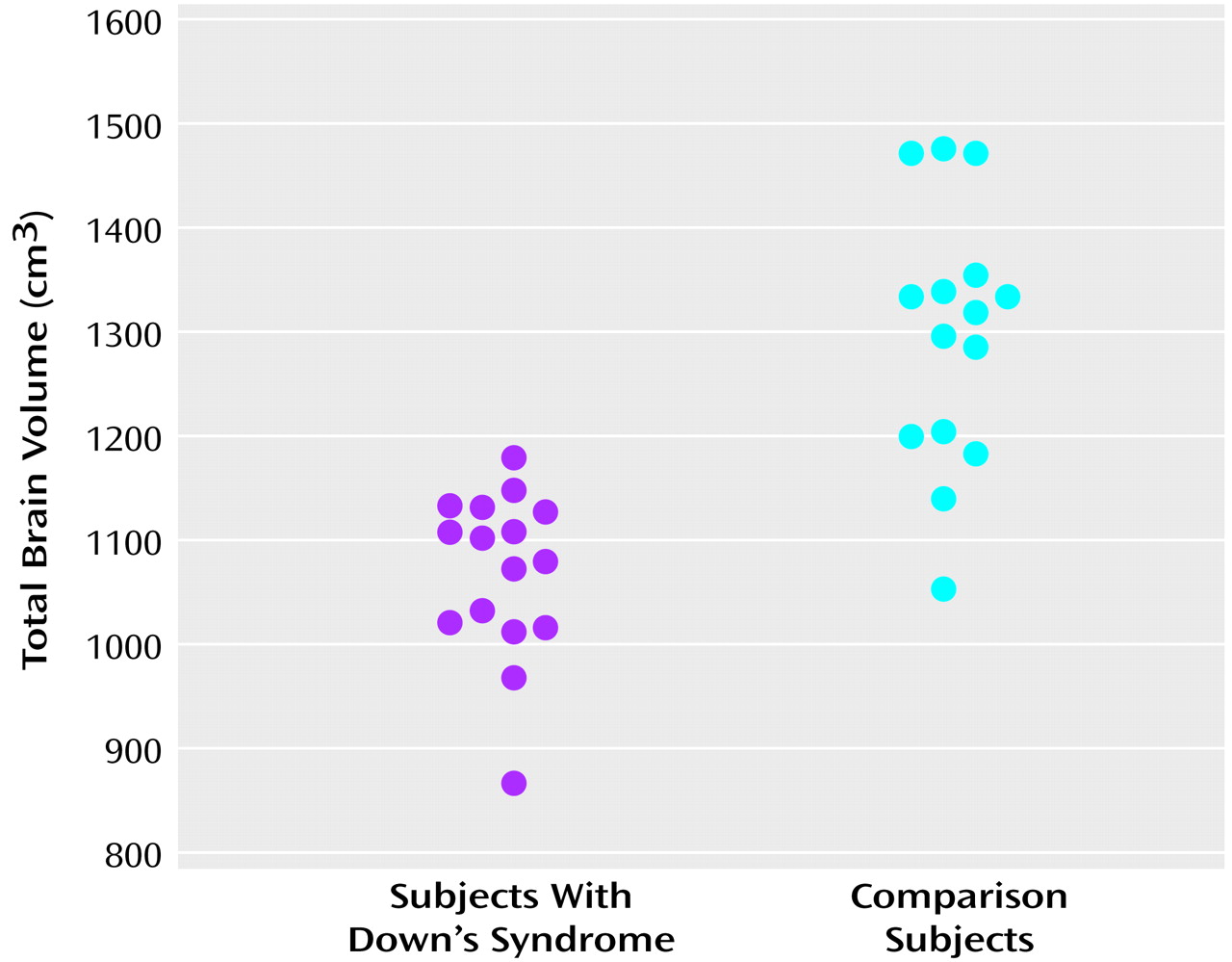
This study is the first we are aware of to specifically evaluate regional brain volumes and tissue composition in Down’s syndrome from early childhood through young adulthood. Our findings indicate that compared to matched developmentally normal subjects, the brains of individuals with Down’s syndrome show 1) overall smaller volumes due to smaller volumes of both cerebral gray and white matter, 2) a disproportionately smaller cerebellar volume, and 3) larger adjusted volumes of subcortical and parietal gray matter and temporal white matter components, with correction for overall brain volumes of gray or white matter, respectively.
Our finding of smaller brain volumes in subjects with Down’s syndrome, including disproportionately smaller volumes of the cerebellum, is consistent with the results of prior pathologic and neuroimaging studies
(3,
10,
16,
18). The cerebellar hypoplasia evident in Down’s syndrome has been suggested to be a causal factor for its characteristic hypotonia and motor coordination difficulties, as well as articulatory speech disturbances, found in most children with Down’s syndrome
(29). Schmahmann and Sherman
(30) showed that patients with cerebellar lesions have impairments in many nonmotor areas, including those involving executive function, verbal fluency, and specific language deficits, including agrammatism. Silveri et al.
(31) also reported specific agrammatism resulting from a focal cerebellar lesion. Functional neuroimaging studies, with both positron emission tomography and functional MRI, have provided further evidence for an important role for the cerebellum in language and cognition
(32,
33). Taken together, the lesion and functional neuroimaging evidence point to the possibility that the syntactic difficulties seen in individuals with Down’s syndrome may be due to cerebellar hypoplasia and dysfunction
(4).
Frontal lobe volumes also were significantly smaller in the subjects with Down’s syndrome, but when they were adjusted for overall brain volume, these smaller volumes were not significant. The frontal lobes have been frequently implicated in the cognitive deficits of Down’s syndrome, including executive dysfunction, inattention, and a tendency toward perseveration. Although our findings do not confirm previous reports of specific “hypofrontality”
(18), their proportionately much smaller volumes may still reflect sufficient underdevelopment to account for dysfunction.
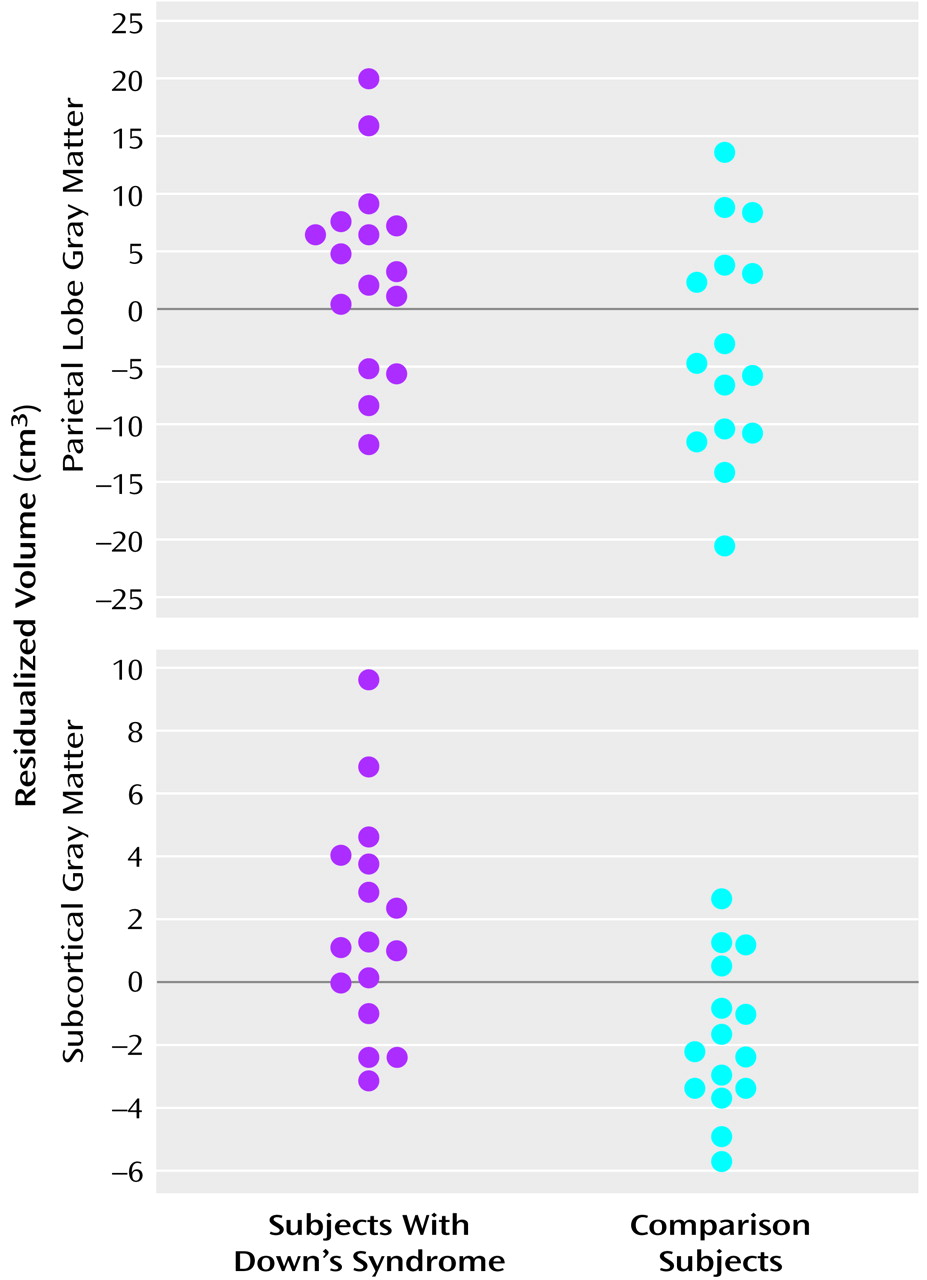
Contrary to the absolute smaller cerebellar and frontal lobe volumes found, but consistent with findings from prior studies of both adults and children with Down’s syndrome
(14,
17,
18), was the remarkable preservation of subcortical structures, reflected in our study by larger adjusted subcortical gray matter volumes in the Down’s syndrome group. The relatively large size or preservation of these structures in children with Down’s syndrome in the context of significantly smaller overall cerebral volumes suggests that there is a temporal dissociation for the development of cortical versus subcortical regions. The embryologic data supporting this view include studies showing that neither brain volume differences nor neuropathologic abnormalities are seen in fetal Down’s syndrome brains until the third trimester, well after the majority of basal ganglia development is complete, but while dendritic arborization, synaptogenesis, and laminar organization continue in the cerebral cortex
(11,
34–
36). One interpretation of relatively larger subcortical volumes is that the basal ganglia are relatively normal due to their having developed extensively before the onset of major abnormalities. On the other hand, relatively larger size also may reflect insufficient programmed cell death, resulting in excessive cell numbers and large but dysfunctional basal ganglia and thalami.
An intriguing finding in our study is the striking preservation of parietal lobe gray matter. This confirms and further specifies the finding by Jernigan et al.
(18) in six young Down’s syndrome subjects of preserved posterior (parietal and occipital) cortical gray matter. Our finding is particularly interesting in light of the neuropsychological profile of Down’s syndrome subjects, which reveals deficits predominantly in language skills (including verbal memory), with relative strengths in visuospatial processing, including visuospatial short-term memory
(5,
6,
8). Evidence that the parietal lobes are important for visuospatial skills comes from both lesion studies
(37,
38) and from functional neuroimaging studies showing bilateral parietal activations in short-term visuospatial memory tasks
(39). The selective preservation of parietal lobe gray matter in Down’s syndrome is consistent with the observed relative strength in visuospatial skills.
In regard to our investigation of structures potentially involved in the language deficits of Down’s syndrome, we found no neuroimaging evidence for our hypothesis of disproportionately smaller overall temporal lobe volumes. In fact, we found larger corrected volumes of the temporal lobes that approached significance. Further segmentation by tissue type revealed this large size to be due to significantly larger corrected temporal lobe white matter volumes in the Down’s syndrome group. It is unclear whether the adjusted larger temporal lobe white matter volumes observed in this study could be related to cognitive deficits. The possibility of an association between larger regional volumes and cognitive dysfunction is supported by studies showing larger parahippocampal gyrus volumes in adults with Down’s syndrome
(13,
14) and an inverse relationship between parahippocampal gyrus volumes and IQ in subjects with Down’s syndrome
(14). Selective smaller hippocampal volumes shown in MRI studies of adults with Down’s syndrome
(13–
15) have highlighted the possibility that important temporal lobe subregional volume abnormalities in either direction may be present throughout development and may contribute to language and memory deficits.
Our expectation of finding smaller volumes of the superior temporal gyrus, on the basis of both functional MRI reports of significant activation of the superior temporal gyrus in auditory and language processing in normal adults
(40–
42) and of the predominant language deficits in children with Down’s syndrome, was not supported by our results. We found no significant difference in adjusted superior temporal gyrus total tissue volumes between the Down’s syndrome and comparison groups, although the corrected superior temporal gyrus white matter volume was significantly smaller in the Down’s syndrome group. It is possible that a selectively smaller white matter volume in this region could contribute to both the observed language deficits and the frequently reported narrowness of the superior temporal gyrus in individuals with Down’s syndrome.
We also found no volumetric evidence for an abnormal pattern of asymmetry of the brain (including the temporal lobes) in Down’s syndrome. This is in contrast to the finding of abnormal rightward temporal asymmetry by Jernigan et al.
(18) in children with Down’s syndrome. Neuropsychological studies of individuals with Down’s syndrome have indicated a pattern of cognitive deficits like that seen in patients with left-hemisphere brain damage
(9). Furthermore, dichotic listening studies have suggested abnormal language lateralization in subjects with Down’s syndrome
(19–
21). Although our results cannot explain these findings on a volumetric basis, functional neuroimaging studies may provide support for lateralized dysfunction.
The limitations of our study include our small group size and its limited age range, which did not include children under age 5. Future studies comparing children without Down’s syndrome but with similar levels of cognitive impairment would be valuable and potentially helpful in critically evaluating whether our findings are specific to subjects with Down’s syndrome. Evidence for syndrome-specific regional brain abnormalities has come from quantitative neuroimaging studies
(18,
43,
44). These have shown that other genetic causes of mental retardation, including fragile X and Williams syndromes, also have unique patterns of structural abnormalities.
Our results largely confirm the findings of previous studies with respect to overall patterns of brain volumes in individuals with Down’s syndrome and also provide new evidence for abnormal volumes of specific regional tissue components. Larger studies are needed to further investigate the parietal lobe gray matter abnormality identified, as well as to specifically evaluate hippocampal and parahippocampal volumes in children with Down’s syndrome. This report represents a starting point for larger longitudinal studies of the neuroanatomic abnormalities of Down’s syndrome throughout childhood.