For example, low levels of dopamine metabolites have been measured in the CSF of depressed patients
(1). Reports of low levels of the dopamine metabolite dihydroxyphenylacetic acid in the CSF of depressed patients
(5) and in the basal ganglia of postmortem brains of depressed suicide victims
(6) lend additional support to a theory of dopamine abnormality in depression. Direct measurement of brain monoamine metabolites from the internal jugular vein of treatment-resistant depressed patients revealed low homovanillic acid levels that were highly correlated with illness severity
(7).
Additional evidence of abnormal dopamine function in depression is suggested by observations of a high prevalence of comorbid depression in patients with Parkinson’s and Huntington’s disease
(8,
9). In these disorders, affective symptoms may precede the onset of motor disturbances, suggesting that mood may be affected by even modest alterations in brain dopamine function and is not merely the result of a psychological reaction to progressive physical impairment
(10).
Imaging studies have also demonstrated abnormalities in the basal ganglia, where dopamine is the predominant monoamine
(11). Magnetic resonance imaging (MRI) signal intensities have been found to be higher in the basal ganglia of some depressed patients than in healthy comparison subjects
(12), while low basal ganglia volume has been detected in depression by using computed tomography (CT) and MRI techniques
(13,
14). Studies using single photon emission computed tomography (SPECT) have shown low regional cerebral blood flow (rCBF) in the basal ganglia, while other studies using positron emission tomography (PET) have identified low glucose metabolism in the basal ganglia of depressed patients. In an extensive review of rCBF and metabolic abnormalities in depressed patients, Soares and Mann
(15) concluded that the most consistent imaging findings are low rCBF and glucose metabolism in the prefrontal cortex and basal ganglia. Recent reports of low metabolism in the ventral striatum
(16) and high metabolism in the right dorsal head of the caudate nucleus
(17) are consistent with the Soares and Mann interpretation
(15).
Finally, experiments with laboratory animals have shown that long-term administration of antidepressants of various pharmacological classes increases activity of dopamine D
2-receptor-like receptors (i.e., D
2, D
3, or D
4)
(4). These effects have been demonstrated with tricyclic antidepressants, selective serotonin reuptake inhibitors, monoamine oxidase inhibitors (MAOIs), α
2 antagonists (e.g., mianserin), mixed reuptake inhibitors (e.g., venlafaxine), trazodone, nomifensine, and electroconvulsive therapy
(18–
21).
We hypothesized that dopamine function is altered in the basal ganglia of patients with depression. In this study, we used the technetium-labeled complex [
99mTc][2[[2-[[[3-(4-chlorophenyl)-8-methyl-8-azabicyclo[3.2.1]oct-2-yl]-methyl](2-mercaptoethyl)amino] ethyl] amino]ethane-thiolato(3-)-N2,N2′,S2,S2′]oxo-[1R-(exo-exo)]
(22) ([
99mTc] TRODAT-1) SPECT brain imaging to examine dopamine transporter activity in depressed patients and healthy comparison subjects.
Method
Patients
Outpatients at least 18 years of age from the University of Pennsylvania Depression Research Unit were enrolled in the study. All underwent a Structured Diagnostic Interview for DSM-IV (SCID)
(23) and met the DSM-IV criteria for a major depressive episode. All patients had a score of at least 16 on the 17-item Hamilton Depression Rating Scale
(24) at the time of the study procedure. All patients were in good physical health as determined by a physical examination and laboratory evaluation including a complete blood count, measurements of serum electrolytes, glucose, and hepatic enzymes, renal and thyroid panels, urinalysis, urine drug screen, and a 12-lead ECG. Women of child-bearing potential had negative results on pregnancy tests.
Patients were excluded from the study if they had a current DSM-IV axis I diagnosis other than major depression, a history of mania, a history of schizophrenia, alcohol or drug abuse within 3 months, or actively suicidal ideation.
The medical exclusion criteria included pregnancy or breast-feeding among women, unstable medical conditions (e.g., angina, diabetes mellitus, hypertension), a history of transient ischemic attacks, intracranial hemorrhage, brain tumor, encephalitis, normal-pressure hydrocephalus, Parkinson’s (or other basal ganglia) disease, malignancy, and hepatic or renal disease. Patients taking chemotherapy, over-the-counter preparations (e.g., St. John’s wort), phenothiazines, barbiturates, mood stabilizers, or antidepressant medications were also excluded.
Before enrollment in the study, patients were required to have taken no antidepressants for at least 7 days, no MAOIs for at least 2 weeks, and no fluoxetine for at least 3 weeks
(25).
Comparison Subjects
Age-matched nonpsychiatric healthy comparison subjects were recruited by investigators at the University of Pennsylvania Division of Nuclear Medicine to participate in [
99mTc]TRODAT-1 studies. The comparison subjects each had a psychiatric evaluation with the SCID format
(23) and had no DSM-IV axis I diagnosis. Each was given a complete physical and laboratory evaluation similar to that for the patients, and none had a significant medical illness or laboratory abnormality. All were free from psychotropic medication, and none had a history of central nervous system disease.
The comparison subjects were age- and gender-matched with the patient group and were consecutively selected from a pool of 101 healthy volunteers who had been studied under similar experimental conditions with [
99mTc]TRODAT-1 SPECT as part of an established normative database of healthy subjects
(26,
27).
Procedures
All subjects were provided with a detailed description of the study purpose and procedures in accordance with the ethical standards set forth by the institutional review board of the University of Pennsylvania. All subjects provided written informed consent before enrolling in the study.
Each subject was placed in a supine, resting position on the SPECT imaging table. A 22-gauge intravenous catheter needle was inserted into an antecubital vein, and a patent intravenous line was maintained with a physiological saline in a sterile cap. Three ECG chest leads were applied for continuous ECG recording. After 20 minutes, [99mTc]TRODAT-1, 740 MBq (20 mCi), was injected through the indwelling venous catheter. The individuals remained at rest for the next 25 minutes, while dynamic SPECT brain images were acquired.
Image Acquisition
Images of the brain were acquired with a triple-head gamma camera equipped with ultra-high-resolution fan-beam collimators (Picker 3000; Picker International, Cleveland). The acquisition variables included a continuous mode with 40 projection angles over a 120° arc to obtain data in a 128×128 matrix with a pixel width of 2.11 mm and a slice thickness of 3.56 mm. The center of rotation was 14 cm. The low-pass filter was based on a systematic analysis of variables that produced the best signal-to-noise characteristics in the images.
Manual demarcation of brain regions of interest was then performed. A set of standardized templates containing small and large regions of interest were fit on each scan. Within the x-y plane, small regions of interest in the template were smaller than the actual structures they represented in order to minimize resolution-induced problems with ill-defined edges. To reduce the effects of volume averaging in the axial direction, the small regions of interest were not placed on the slices that contained the uppermost and lowermost portions of the structures they represented. This tended to limit the small regions of interest to the central aspect of the structures they represented. In contrast, boundaries for the whole brain were drawn by hand on slices located 12 mm above the highest slice that included the basal ganglia.
The primary outcome measure was specific uptake 3 to 4 hours after [99mTc]TRODAT-1 administration, when the distribution of [99mTc]TRODAT-1 approached a transient near-equilibrium-like state that reflected the ratio k3/k4, which is related to [99mTc]TRODAT-1 binding potential.
Statistical Procedures
Intrarater reliability for analyses of the manually defined regions of interest was estimated by using intraclass correlation coefficients. The procedures were implemented with a commercial statistical package (StatS, Redmond, Wash., Think Point Software). This procedure resulted in a high correlation because the high-contrast images were particularly amenable to “punch biopsy” approaches to image analysis, while the “large” regions of interest were difficult to misplace. The reliability of the image analysis techniques was found to be high, with intraclass correlations consistently above 0.95, as expected for work with relatively high-contrast images
(28).
Group comparisons of mean specific uptake values of [99mTc] TRODAT-1 binding for each region of interest were analyzed by using the unpaired Student t test without correction for multiple comparisons. Statistical significance was defined as p<0.05.
Discussion
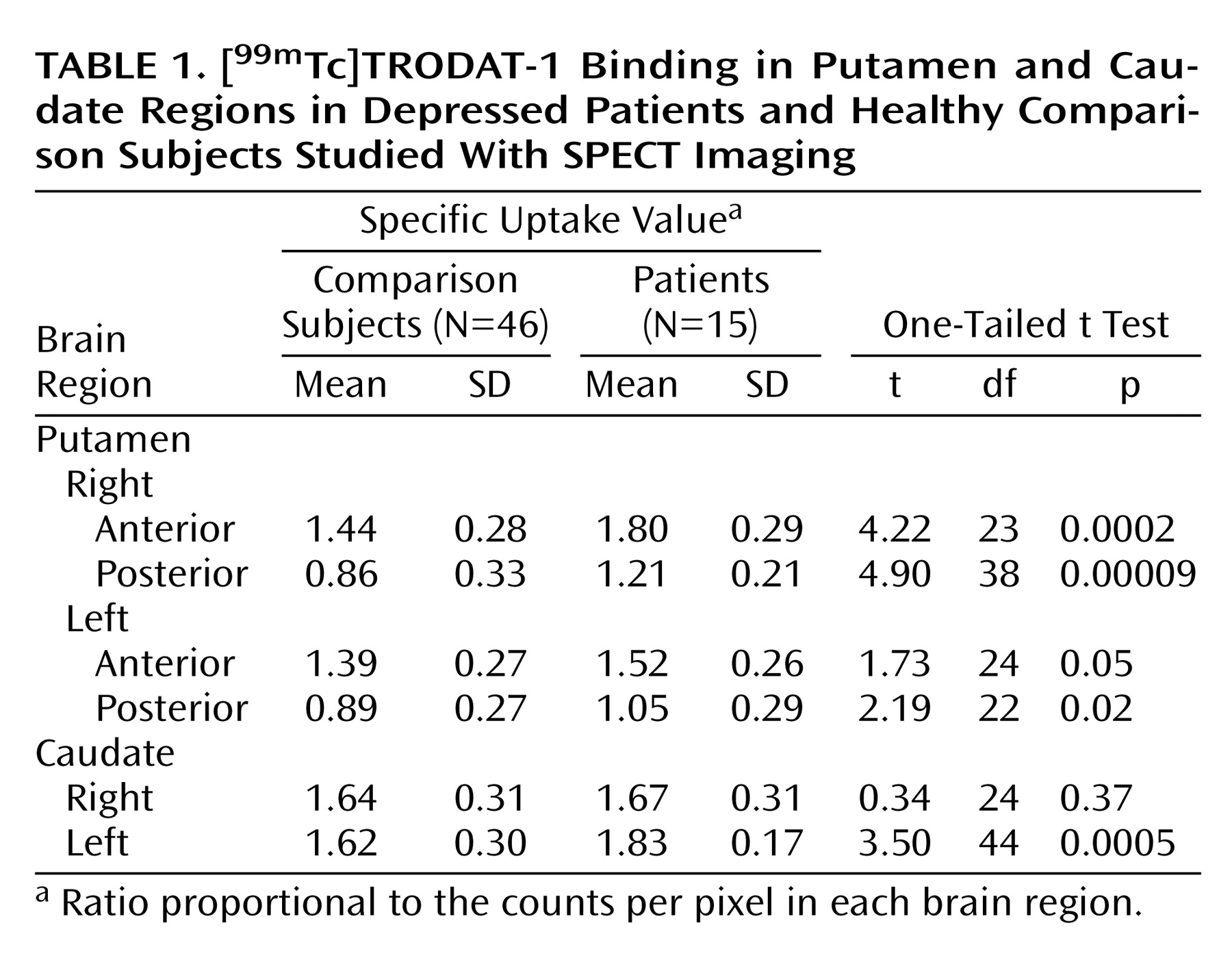
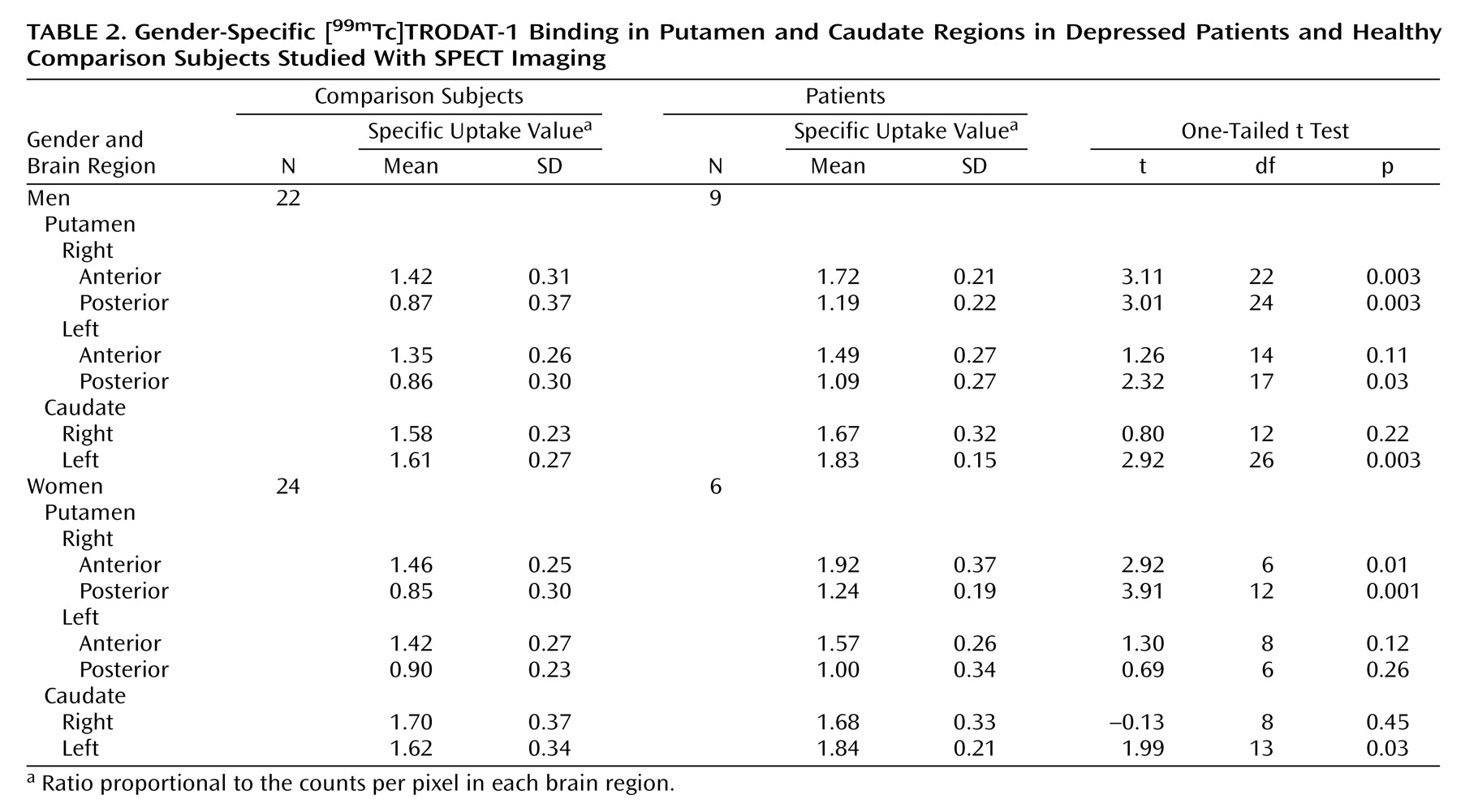
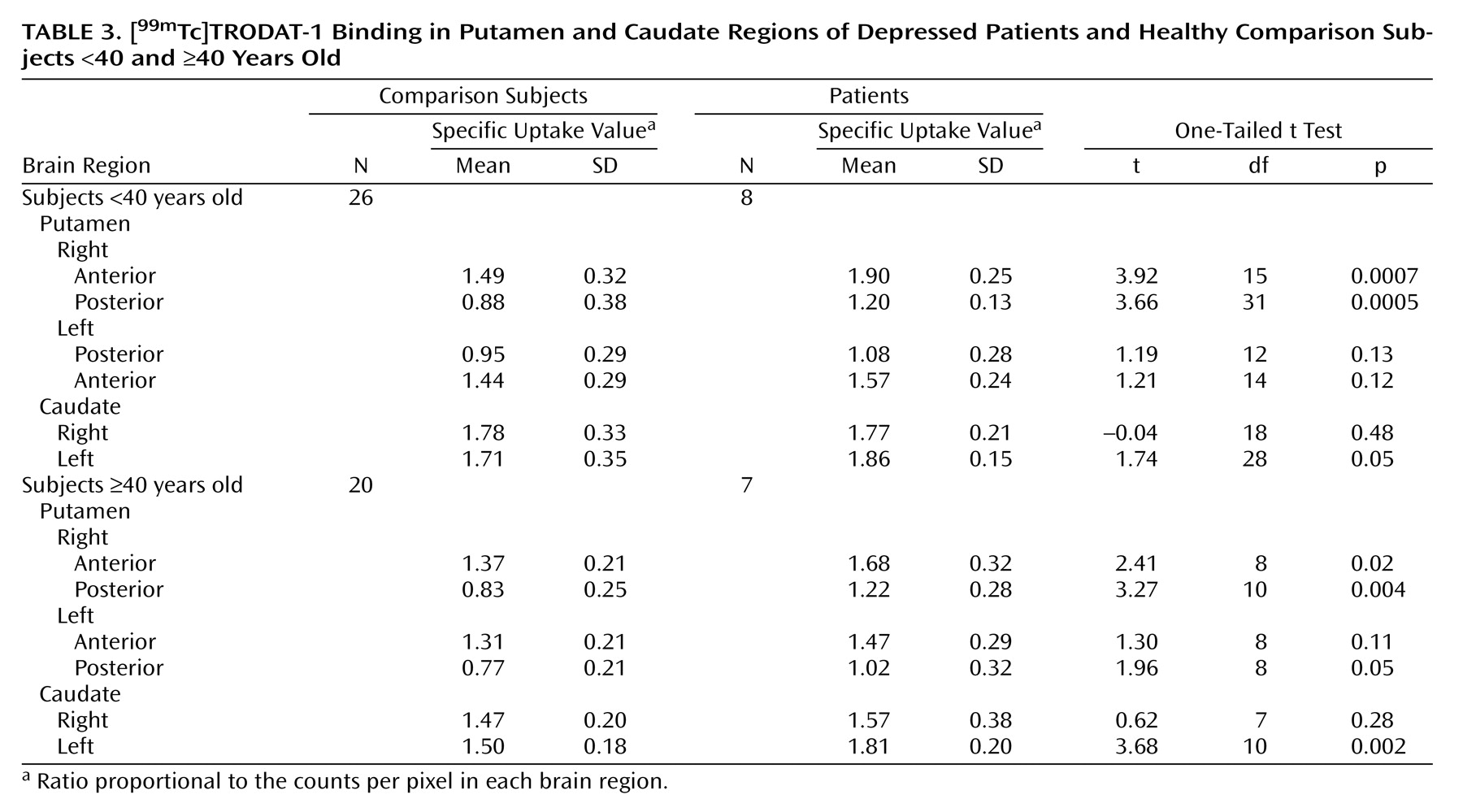
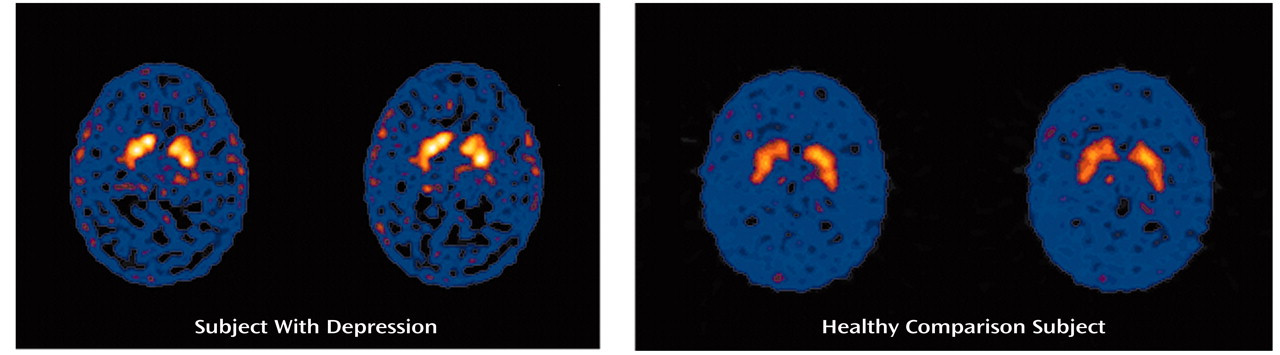
We performed SPECT brain imaging with [99mTc]TRODAT-1, a selective dopamine transporter ligand, to study the availability of dopamine transporters in the basal ganglia region of depressed patients. We observed higher specific uptake values for [99mTc]TRODAT-1 binding in several subregions of the putamen and caudate nuclei of depressed patients than in comparison subjects. Significantly greater [99mTc]TRODAT-1 binding was observed in the right anterior (23%) and right posterior (36%) putamen, with a more modest difference in the left posterior putamen (18%). A higher specific uptake value was observed in the left (12%), but not the right, caudate region.
Although several SPECT studies have previously investigated dopamine uptake sites in depression, we believe that the present study is the first to examine the dopamine uptake site in depression by using the [
99mTc]TRODAT-1 radioligand. Laasonen-Balk et al.
(29) measured striatal dopamine transporter sites in 15 drug-naive depressed patients and 18 healthy comparison subjects by using [
123I]β-CIT, an iodinated radioligand that binds to both dopamine and serotonin transporter sites. They found significantly higher striatal [
123I]β-CIT binding to “presumed” dopamine transporter sites in the depressed patients. In contrast, Malison et al.
(30) reported no significant difference in [
123I]β-CIT binding to striatal dopamine uptake sites between 15 depressed patients and 15 healthy comparison subjects.
The greater dopamine transporter availability observed in the present study could be the result of a decrease in synaptic levels of dopamine or a diminished dopaminergic “tone” in depression. In this regard, the dopamine transporter plays a critical role in regulating synaptic dopamine concentrations by rapidly taking up extracellular dopamine into presynaptic terminals following its release. The dopamine uptake site is the primary determinant of extracellular dopamine concentrations and maintains the balance between dopamine homeostasis and function
(31).
It is also possible that our observation of greater dopamine transporter availability in depression reflects a compensatory mechanism resulting from increased dopaminergic “tone” in depression. For example, D
2 receptor agonists increase dopamine transporter turnover
(32). Since the activity of the dopamine transporter is regulated by the endogenous dopamine “tone,” it may also represent a useful biomarker for studying changes in central dopamine activity in depression.
The greater dopamine transporter specific uptake values observed by Laasonen-Balk et al.
(29) could be related to the psychomotor disturbances associated with depression. The caudate and putamen have direct effects on psychomotor activity
(33), and some investigators have suggested that dopamine may be involved in the pathogenesis of psychomotor slowing in melancholic depression
(34).
Altered dopamine transporter binding could also be related to the presence of mood changes in depression. Transient dysphoria resulting from tryptophan depletion has been found to produce reductions in striatal rCBF and glucose metabolism. Bremner et al.
(35) observed low glucose metabolism in the caudate and putamen by using PET, while Smith et al.
(36) observed low rCBF in the left, but not the right, caudate region with PET. The finding supports our observation of high dopamine transporter specific uptake values in the left caudate region in depression. Similarly, others have reported that transient sadness is associated with increased rCBF in the caudate and putamen
(37).
The presence of striatal dopamine dysregulation could affect cortical and limbic lobe function as well. In this model, the caudate nucleus is part of a complex system in which specific cortical pathways project to the striatum and then to the globus pallidus, substantia nigra, and finally, back to the cortical areas
(9,
38). In addition, the striatum receives neuronal inputs from other brain regions, such as the thalamus and amygdala. Several of these pathways involve neuronal circuits that modulate mood and emotional behavior. As a result, a dysfunction of dopamine in the striatum could impair the corticostriatal circuits disrupting limbic input back to the cortex in depression
(39,
40).
Finally, our observation of differences in [
99mTc]TRODAT-1 binding between the right and left sides of the striatum may result from chirality in dopamine receptor density, which appears to be higher in the right than in the left striatum
(41).
Several caveats should be considered in the interpretation of our findings. For example, women were studied without regard to the phase of the menstrual cycle. This factor may influence dopamine transporter number and [
99mTc]TRODAT-1 binding
(42). In addition, diagnostic heterogeneity, differences in illness length and episode duration, and degree of psychosocial stress may have influenced the differences in [
99mTc]TRODAT-1 binding between the patients and comparison subjects. Finally, differences in physical well-being, activity level, disruption of normal sleep, and circadian rhythms may have influenced [
99mTc]TRODAT-1 binding in the patients.
In conclusion, by using the highly selective dopamine transporter radioligand [99mTc]TRODAT-1 and SPECT imaging to examine the availability of striatal dopamine transporter sites in 15 depressed patients and 46 healthy subjects, we found significantly higher specific uptake values of [99mTc]TRODAT-1 in the right anterior and right posterior putamen and in the left posterior putamen and left caudate regions of the depressed patients.