Anxiety disorders are the most common class of psychiatric disorders and are often associated with significant functional impairment (
1). Symptoms of anxiety disorders can begin in infancy or toddlerhood (
2), and the median age at onset of anxiety disorders has been reported to range from 6 to 11 years (
1,
3). Most adults with anxiety disorders experienced symptoms during childhood (
4). The infant temperament behavioral inhibition, characterized by increased attention and distress to novel stimuli (
5,
6), is one of the most potent known risk factors for developing an anxiety disorder later in life (
7). This continuity between infant temperament, symptoms in early childhood, and disorders in adulthood suggests that behavioral inhibition and anxiety disorders represent an early departure from typical brain development (
8). Unfortunately, the timing and nature of this atypical brain development are poorly understood. Moreover, it is difficult to predict which neonates will exhibit high behavioral inhibition during infancy (
6), and behaviors in the first year of life are only moderately predictive of behavioral inhibition in boys and not predictive of behavioral inhibition in girls at age 2 (
9). Consequently, at-risk children may not be identified until many years after the onset of atypical brain development. The present study addresses these issues by identifying brain network variation at birth that is related to levels of behavioral inhibition at age 2 years.
Indirect evidence supports the hypothesis that the physiology of behavioral inhibition during infancy includes an overactive ventral attention network. The ventral attention network is a cortical functional brain network involved in directing attention to novel, unexpected stimuli (e.g., a stranger or a new toy) and includes portions of the right ventrolateral prefrontal cortex and right temporal-parietal junction (
10). Infants with high behavioral inhibition exhibit indirect behavioral and physiological signs of an overactive ventral attention network, including increased attention and increased right lateralized brain activity (as measured by EEG) after presentation of novel stimuli such as toys or a stranger (
11,
12). Reports of ventral attention network structural and functional abnormalities in older children and adults with anxiety disorders provide further support for this hypothesis (
13,
14).
While the ventral attention network may play a role in behavioral inhibition during infancy, previous work indicates that many other brain regions and systems are associated with behavioral inhibition in older children and adults. Adolescents and adults who had high behavioral inhibition during early childhood, for example, have increased amygdala reactivity to novelty (
15,
16) and altered striatal activity in response to reward (
17,
18) relative to peers. Similarly, older children and adults who had high behavioral inhibition during early childhood have altered functional connectivity within and between the default mode, salience, and other brain networks (
19), as well as altered connectivity between the amygdala and cortical regions that may be within these same networks (
20,
21). While amygdala functional connectivity alterations associated with behavioral inhibition are apparent at the time of birth (
22), it is unknown whether altered connectivity within and between cortical brain networks is also already evident in the neonatal period. Given the central role of functional brain networks in brain organization and information processing (
23), uncovering disruptions that are already present at birth is likely to have fundamental implications for the developmental neurobiology of behavioral inhibition and anxiety disorders.
The goal of this study was to identify variation in cortical functional brain networks evident soon after birth that is related to behavioral inhibition at age 2, in order to elucidate the neural substrate of behavioral inhibition and identify early risk factors. To achieve this goal, we used functional MRI (fMRI) to relate variation in resting-state functional connectivity of brain regions putatively within the ventral attention, default mode, and salience networks in 45 neonates at term-equivalent age to measures of behavioral inhibition at age 2. This prospective, longitudinal approach provides an unprecedented window into the initial stages of the developmental neurobiology of behavioral inhibition and anxiety disorders. By identifying variation in brain network connectivity shortly after birth and before temperamental characteristics of behavioral inhibition emerge, the results from this study most likely reflect variation in fetal brain development. Results cannot be attributed to early childhood experiences that are a consequence of having a behavioral inhibited temperament. Based on the indirect evidence cited above relating behavioral inhibition to the ventral attention network during infancy, our primary hypothesis was that functional connectivity of the ventral attention network at term-equivalent age would be associated with behavioral inhibition at age 2. We also tested whether functional connectivity of the default mode and salience networks was associated with behavioral inhibition, because of the role of these networks in older children and adults with behavioral inhibition. We previously (
22) related functional connectivity of the amygdala during the neonatal period to internalizing symptoms (including behavioral inhibition) at age 2 in a subset of this cohort, demonstrating the feasibility of our approach. To ensure that our primary analysis was sensitive to the strongest brain-behavior relations, we performed two follow-up analyses. In a confirmatory analysis, we related functional connectivity among 18 regions (comprising 153 region-region pairs) at term-equivalent age to behavioral inhibition at age 2. These regions are frequently implicated in psychiatric illnesses and are distributed across the brain (
13,
14,
19,
20,
24–
26). In a final exploratory analysis, we examined similar relations among 216 regions, covering most of the cerebral cortex and much of the subcortex and comprising 23,220 region pairs. The primary, confirmatory, and exploratory analyses converged on a consistent set of results, suggesting that our primary, hypothesis-driven analysis detected the strongest relations between neonatal functional connectivity and behavioral inhibition at age 2.
Method
This study was approved by the Washington University Human Studies Committee. Parental written informed consent was obtained for all subjects recruited from Washington University School of Medicine hospitals. A group of preterm infants (N=27; born at gestational age ≤30 weeks) were recruited from St. Louis Children’s Hospital Neonatal Intensive Care Unit, and a group of term infants (N=18; born at gestational age ≥36 weeks) were recruited from the adjoining mother-baby unit at Barnes-Jewish Hospital. Full-term infants underwent MRI scanning within 4 days of birth and were scanned at a mean postmenstrual age of 39.7 weeks SD=1.0). Preterm infants underwent MRI scanning at term-equivalent postmenstrual age 37.5 weeks (SD=1.3). Full inclusion and exclusion criteria, a comparison of included and excluded subjects, and MRI acquisition parameters are provided in the data supplement that accompanies the online edition of this article.
Follow-Up at Age 2
Age 2 assessments included the Infant-Toddler Social and Emotional Assessment (ITSEA), a 166-item parent-report measure with good psychometric properties (
27). Based on study hypotheses, this analysis focused on the behavioral inhibition subscale of the ITSEA, which has acceptable internal consistency (Cronbach’s alpha=0.77) and is within the internalizing domain. Previous work indicates that ITSEA ratings, including behavioral inhibition, correlate significantly with direct observations of infant behaviors by trained evaluators (
27,
28). ITSEA behavioral inhibition scores are also highly correlated with the inhibition to novelty score of the Infant Behavior Questionnaire, another commonly used measure of infant temperament (
28). We also examined several factors that could be associated with infant behavioral inhibition, including social disadvantage (
29) and a maternal history of affective symptoms (
30). Details of these measures are provided in the Supplementary Methods section in the
online data supplement. Zero-order relationships among all variables of interest in this study are described in the Supplementary Results section and Table S1 in the
data supplement.
MRI Preprocessing
Resting-state fMRI data were preprocessed with in-house software (
ftp://imaging.wustl.edu/pub/raichlab/4dfp_tools/); the FMRIB Software Library was also used for field map correction. Magnetization inhomogeneity–related distortions were corrected using a mean field map technique (
31). Atlas transformation was computed using infant templates. Volumetric time series in adult Talairach atlas space (3×3×3-mm voxels) were generated, combining field map correction and atlas transformation in a single resampling step. Additional preprocessing included regression of nuisance waveforms derived from rigid body motion correction, CSF and white matter regions, plus whole-brain global signal. The data were low-pass filtered (<0.08 Hz) and spatially smoothed. Frames affected by movement (volume-to-volume head displacement ≥0.25 mm) were excluded from the resting-state fMRI computations (
32). A minimum of 5 minutes of fMRI data, excluding censored frames, was required for inclusion in the analysis. Forty-five subjects survived this stringent volume-censoring threshold.
Regions of Interest
Regions of interest were selected from a commonly used set of regions (the “Power 264 set”) in which the functional network identities in adults are well established (
23) and which have been used in previous studies of neonates (
33). The specific regions are listed in Table S2 in the
online data supplement. For the primary, hypothesis-driven analysis, we selected a region in the right ventrolateral prefrontal cortex for the ventral attention network, the dorsal anterior cingulate cortex for the salience network, and the medial prefrontal cortex for the default mode network. We chose these three particular seeds because they have been consistently implicated in studies of behavioral inhibition and anxiety disorders (
13,
14,
19,
20,
24–
26). Mean connectivity maps for these seed regions are presented in Figure S1 in the
data supplement and support the functional network assignments. For the confirmatory region-to-region based analyses, we examined relations among 18 regions from the Power 264 set that are distributed across the brain and are representative of five functional brain networks frequently implicated in neuropsychiatric diseases. These 18 regions define 153 region pairs (each of the 18 regions of interest can be correlated with the 17 others, and the total divided by two to avoid double counting). For a final exploratory analysis (see the
data supplement), we examined relations among all 214 regions in the Power 264 set that can be accurately mapped to the infant brain, plus individualized left and right amygdala regions that were the focus of a previous study using the present data set (
22). These 216 regions define 23,220 region-to-region pairs.
Statistical Analysis
Whole-brain voxel-wise analyses were performed using in-house software (
www.nil.wustl.edu/labs/fidl/). All other analyses were conducted with SPSS, version 22 (IBM, Armonk, N.Y.). For the primary, hypothesis-driven analysis, the functional connectivity of each of the three seed regions to every other voxel in the brain was computed in each subject. Correlation coefficients were Fisher z-transformed, generating z(r) correlation maps; average maps are depicted in Figure S1. These voxelwise whole-brain functional connectivity maps were the dependent variables in a series of regression analyses that included behavioral inhibition, sex, social risk (see the
data supplement for details on the social risk index), behavioral inhibition-by-sex interaction, behavioral inhibition-by-social risk interaction, and postprocessing residual average framewise displacement. Sex and social risk were included because they were related to behavioral inhibition. The three resulting voxel-wise maps corresponding to behavioral inhibition in the regression model (one for each seed region) were masked to include only voxels in cortical gray matter. Each map was corrected for multiple comparisons using AFNI tools as currently recommended (
34,
35). Further details are provided in the Supplementary Methods section in the
data supplement.
To test whether our primary seed-based analysis was sensitive to the strongest brain-behavior relationships, we performed a confirmatory region-to-region analysis. This analysis was done to ensure that our selection of three seeds for the primary analysis captured the functional connections at term most strongly associated with behavioral inhibition at age 2. For this confirmatory analysis, we first computed functional connectivity for each of the 153 region pairs for each subject. Next, we performed the identical regression analysis described above. Standardized betas were computed for the association between behavioral inhibition at age 2 and functional connectivity of each region pair at term. Betas that were significant at p<0.05, uncorrected, were examined to determine whether the hypothesis-driven seed-based analysis above captured the most relevant relationships between behavioral inhibition and functional connectivity. To further contextualize the findings, we performed an exploratory analysis examining relationships among 216 regions (comprising 23,220 region pairs); these exploratory results are described in the data supplement.
Results
The demographic characteristics of the sample are summarized in
Table 1. Behavioral inhibition at age 2 was significantly negatively associated with social risk score at age 2 (r=−0.34, p=0.02), and girls had nonsignificantly greater behavioral inhibition relative to boys (mean difference, 0.3 points; t=2.0, df=43, p=0.06). History of preterm birth and history of maternal affective disorders were unrelated to behavioral inhibition at age 2. Motion during neonatal scanning after frame censoring was also unrelated to behavioral inhibition at age 2. Based on these results, all imaging analyses below include sex, social risk, the interaction between sex and behavioral inhibition, the interaction between social risk and behavioral inhibition, and postprocessing motion (framewise displacement) as covariates.
Hypothesis-Driven Seed-Based Functional Connectivity Analyses
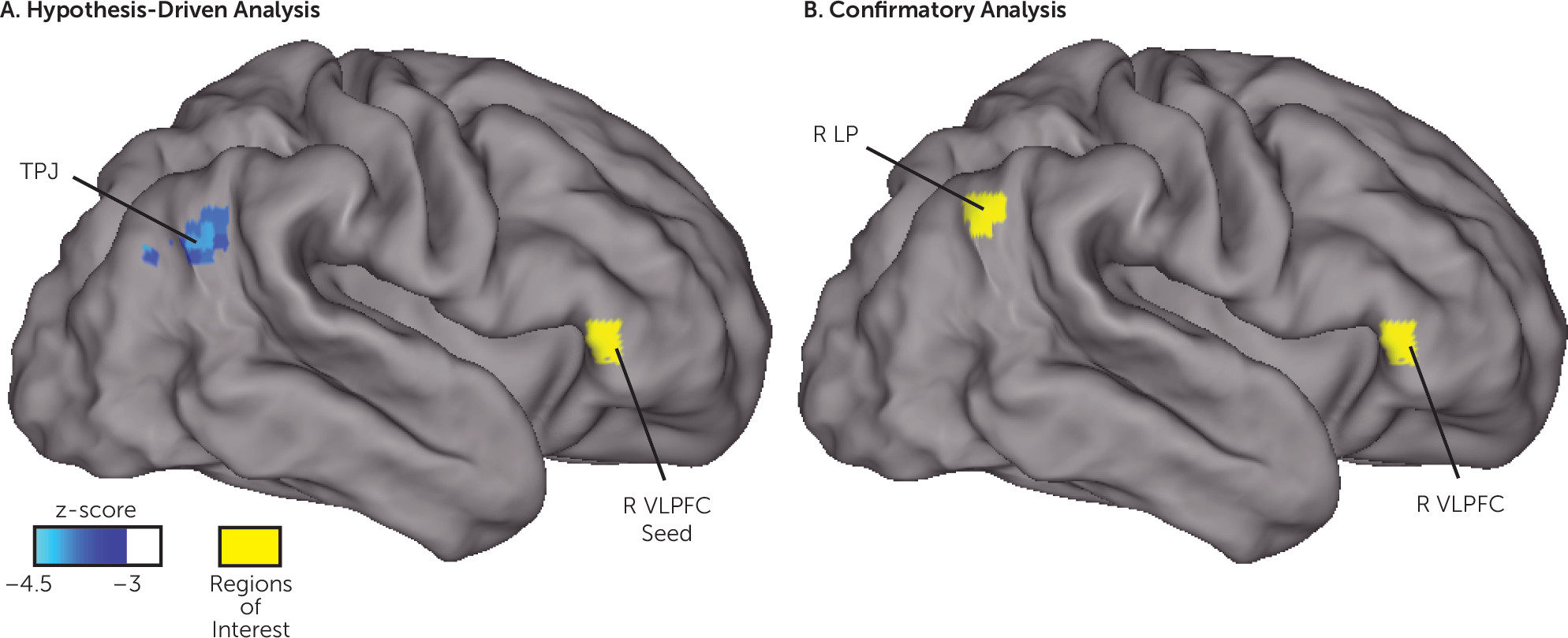
As depicted in
Figure 1A, functional connectivity between the right ventrolateral prefrontal cortex seed region (located in the ventral attention network in adults) and a cluster near the right temporal-parietal junction was negatively associated with behavioral inhibition at age 2 (peak z-score=−4.3, cluster size=1,323 mm
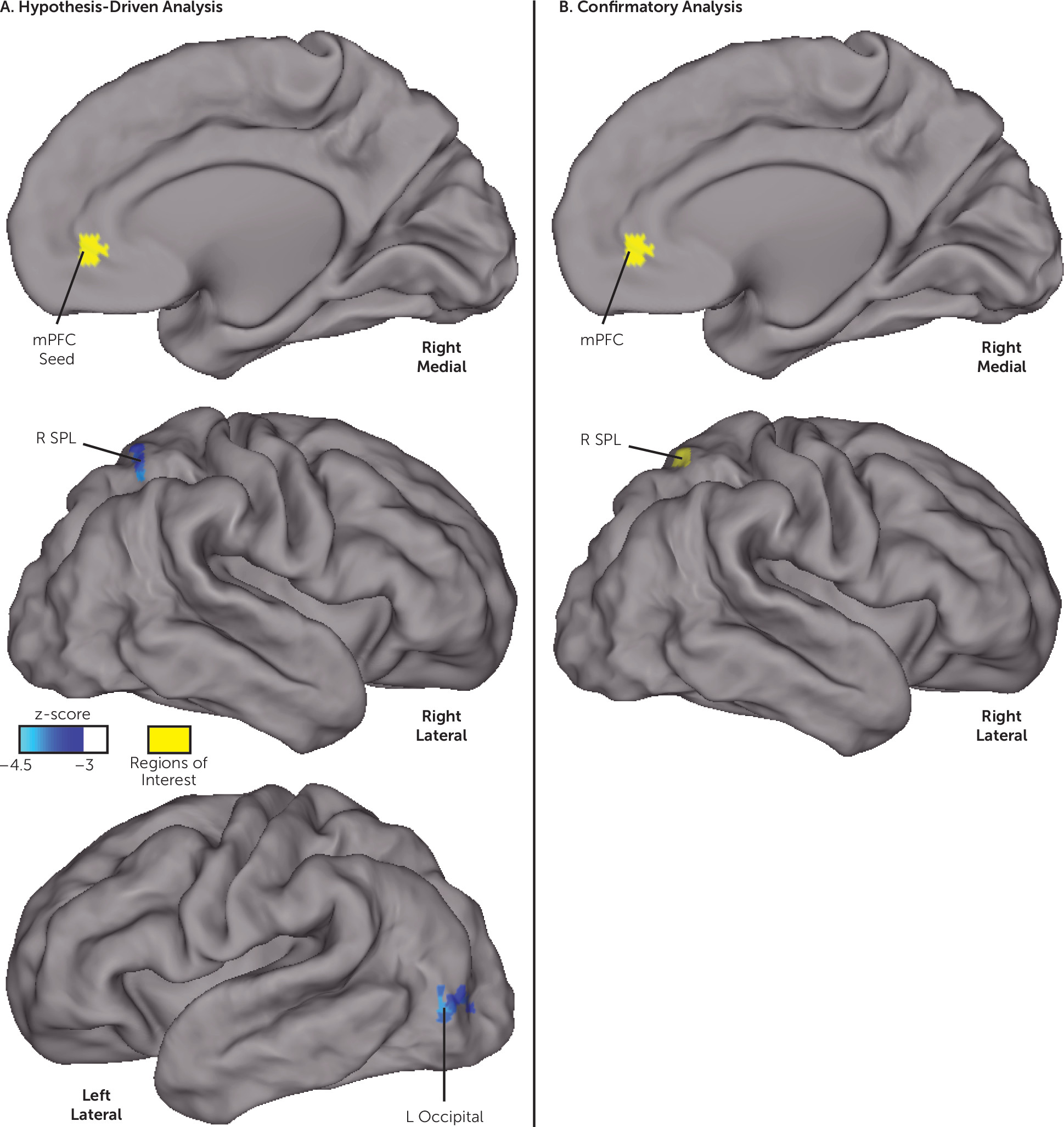
3, with all voxels z<−3.0, centered at 51, −60, 27 in Talairach coordinates). As depicted in
Figure 2A, neonatal resting-state functional connectivity between the medial prefrontal cortex (located in the default mode network in adults) and two distinct clusters was negatively associated with behavioral inhibition at age 2. These clusters were located within the right superior parietal lobule (peak z-score=−4.1, cluster size=1,188 mm
3, centered at 26, −59, 52) and in the left lateral occipital cortex (peak z-score=−4.0, cluster size=1,053 mm
3, centered at −42, −81, −2). The whole-brain analysis with the dorsal anterior cingulate cortex seed (from the salience network in adults) did not detect any relationships surviving correction for multiple comparisons.
Confirmatory Region-of-Interest Functional Connectivity Analyses
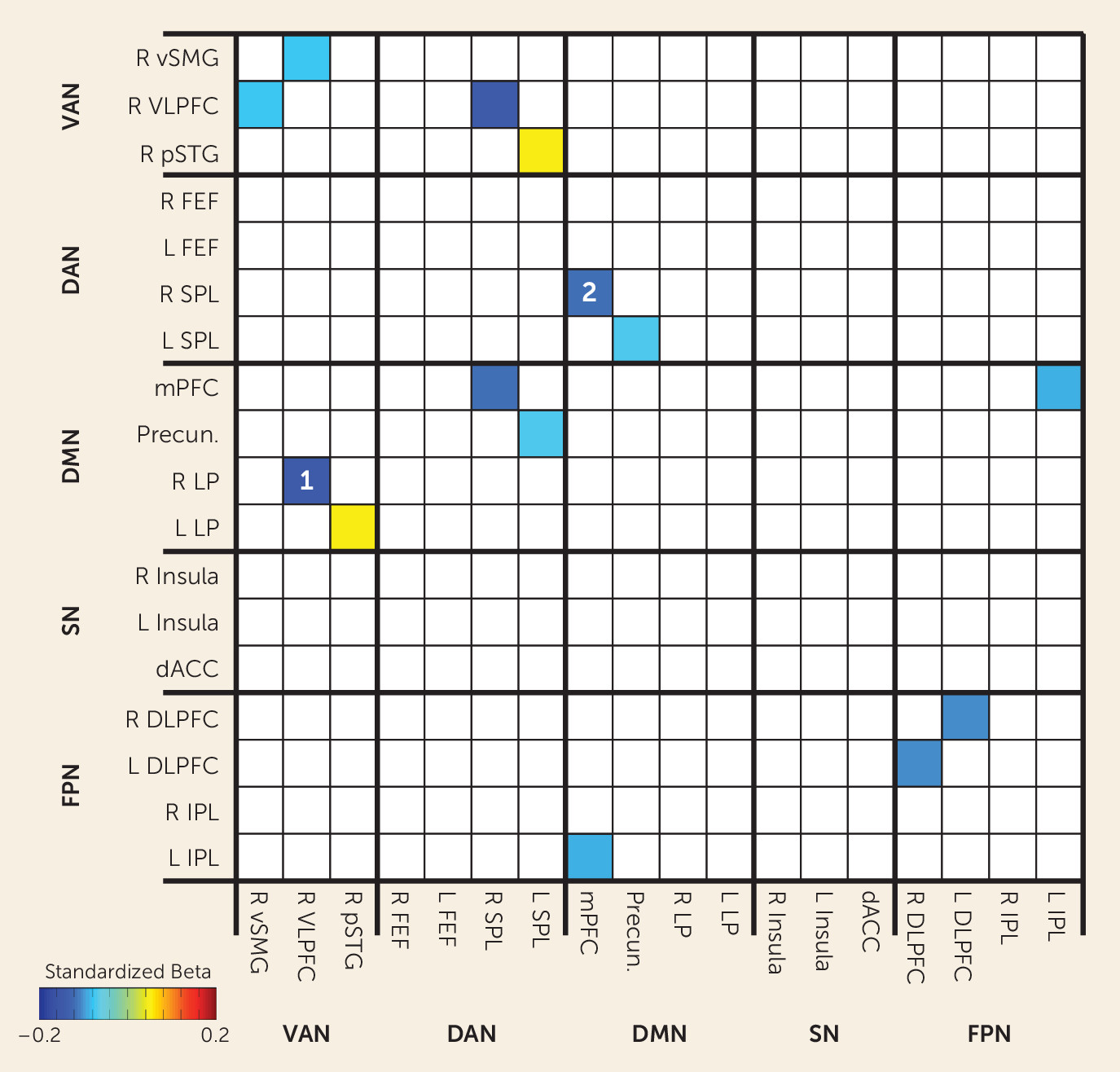
We performed an additional confirmatory analysis to verify that the three seeds we selected for the primary analysis captured the strongest relationships between functional connectivity at term-equivalent age and behavioral inhibition at age 2. For this analysis, the goal was to determine whether we would have obtained stronger results if we had chosen to use other seed regions that are commonly used in studies of neuropsychiatric disorders. In this confirmatory analysis, we examined functional connectivity among 18 regions of interest (yielding 153 region-region pairs) from five functional brain networks commonly implicated in neuropsychiatric illnesses; three of these 18 regions were used as seeds in the hypothesis-driven analysis described above.
Figure 3 depicts standardized beta weights for the regression analyses relating each of these region-region pair functional connectivity values during the neonatal period to behavioral inhibition at age 2. Only standardized beta weights in which p<0.05 are presented. Of the 153 region pairs examined, the region pair with largest effect size included the right ventrolateral prefrontal cortex region that was used as a seed for the ventral attention network and a region in the right lateral parietal cortex, near the temporal-parietal junction, that maps to the default mode network in adults (standardized beta=−0.14, indicating that for every one-standard-deviation increase in behavioral inhibition, the connectivity value decreases by 0.14). The region pair with the second largest effect size included the medial prefrontal cortex region that was used as a seed for the default mode network and a region in the right superior parietal lobule that maps to the dorsal attention network (standardized beta=−0.11).
Figures 1B and
2B depict these two connections with results from the whole-brain analyses side by side to illustrate that both the primary and confirmatory analyses converge on similar results. In the Supplementary Results section and Figure S2 in the
data supplement, results from the whole-brain, hypothesis-driven analyses are further contextualized in an exploratory analysis examining relationships among 216 regions of interest that cover most of the brain (yielding 23,220 region pairs).
Discussion
The goal of this study was to examine associations between functional connectivity of cortical functional brain networks evident soon after birth and behavioral inhibition at age 2. Hypothesis-driven and confirmatory analyses converged on a common set of results: the functional connectivity of a right ventrolateral prefrontal cortex seed (which maps to the ventral attention network in adults) and a medial prefrontal cortex seed (which maps to the default mode network in adults) in the neonatal period was associated with behavioral inhibition at age 2. Notably, neonatal connectivity was related to behavioral inhibition at age 2 beyond information provided by sociodemographic factors, as all analyses controlled for these other factors.
Our results provide the first direct evidence for the previously hypothesized link between the ventral attention network and behavioral inhibition. Previous authors have hypothesized that behavioral inhibition is associated with an overactive ventral attention network because behavioral inhibition includes increased attention to novel stimuli, the putative function of the ventral attention network (
13,
36). In addition, infants with behavioral inhibition have increased right lateralized brain activity, as measured by EEG, in response to novel stimuli relative to infants without behavioral inhibition (
37); the ventral attention network is a plausible contributor to this increased activity given that it is also right lateralized. Our results also indicate that previously identified functional connectivity alterations associated with behavioral inhibition in the default mode network in childhood and adulthood (
19–
21) are already evident in early infancy. Given the role of the default mode network in self-focused processes, previous authors have suggested that default mode network connectivity alterations in behavioral inhibition may reflect an increased internal focus (
19); our results suggest that the precursors of this process are evident at birth.
The continuity of the infant temperament behavioral inhibition, symptoms of anxiety in childhood, and increased risk for anxiety disorders throughout the lifespan suggest that behavioral inhibition and anxiety disorders represent an early departure from typical brain development (
38). The present results suggest that this departure from typical brain development may have its origin in fetal brain development, since alterations in functional connectivity are already evident soon after birth. We previously demonstrated, in this same cohort, that functional connectivity of the amygdala during the neonatal period is associated with internalizing symptoms, including behavioral inhibition at age 2 (
22). It appears, therefore, that disruptions in the ventral attention network, the default mode network, and amygdala functional connectivity may represent the initial stage in the cascade that leads to behavioral inhibition and potentially to anxiety disorders. Other problems may emerge as functional connectivity continues to develop, and other forces may turn early risk into clinically symptomatic anxiety disorders. Longitudinal studies are required to test this hypothesis.
Identifying the first stages in the pathway to an anxiety disorder provides an opportunity for early identification and intervention of high-risk individuals. Previously identified risk factors are only moderately predictive at birth of behavioral inhibition and anxiety disorders in later childhood (
6,
29,
30), so most children with anxiety disorders are not identified until many years after the initial departure from typical brain development. Our results suggest that functional connectivity soon after birth could serve as a biomarker for guiding early interventions such as parent training and early socialization of at-risk infants. Given that the present study suggests that the origins of behavioral inhibition may lie in fetal brain development, functional connectivity shortly after birth could also serve as an outcome measure for perinatal interventions targeting risk for anxiety disorders (
39).
This study’s results should be interpreted in light of several limitations. The borders of functional brain networks in infants are the subject of ongoing work, so the functional network identities of regions are not fully defined. We label regions based on adult network identities, but there are almost certainly differences in functional network organization between infants and adults. The results should therefore be interpreted as reflecting properties of regions that become parts of particular functional networks in later childhood and adulthood rather than definitively residing in those networks at birth. Current work generally supports the hypothesis that adult network identities can be applied to data on newborns, although local connections and bilateral connections between homologous regions are much stronger than longer-range anterior-posterior connections (
40). Whole-brain fMRI analyses have historically been contaminated by unacceptably high false positive rates; in this study we utilized a conservative correction criterion defined by our own data set, as currently recommended by experts (
34,
35). Contamination of functional connectivity by submillimeter frame-to-frame motion is also a major source of artifacts (
32); we used strict motion-correction procedures in this study. Finally, behavioral inhibition was assessed by maternal report, and future studies could extend these results by combining maternal report with laboratory observational measures.
In summary, decreased functional connectivity of regions in the ventral attention and default mode networks at term-equivalent age was associated with behavioral inhibition at age 2. Neonatal functional connectivity was related to behavioral inhibition beyond sociodemographic factors, including sex and social risk. These data potentially identify the earliest stage of the neurodevelopmental cascade leading to behavioral inhibition and anxiety disorders. These results have fundamental implications for the neurobiology of anxiety disorders and could aid in early risk stratification and intervention.
Acknowledgments
Supported by NIH (grants K23MH109983, K23MH105179, R01HD057098, R01HD061619, KL2TR000250, UL1TR000448, K02NS089852, R01NS06424, and R01NS32970), the Intellectual and Developmental Disabilities Research Center at Washington University (grant P30HD062171), the McDonnell Center for Systems Neuroscience, the Taylor Family Institute, the Parker Fund for Young Investigators in Psychiatry, the McDonnell Foundation Collaborative Activity Award, the Child Neurology Foundation, the Cerebral Palsy International Research Foundation, the Dana Foundation, and the Doris Duke Foundation.