Agitation is a common neuropsychiatric symptom in demented elderly patients.
1–5 It is one of a group of behavioral complications that often accompany the core cognitive impairments in Alzheimer's disease (AD). Agitation in AD manifests as restlessness, excessive and repetitive verbal production, and perseverative locomotion such as pacing,
1 and it can escalate to noisy, aggressive activity.
6 The management of agitated elderly patients is a serious clinical issue and a key factor leading to institutionalization, since agitation may have detrimental effects on the agitated patient, on caretakers, on other patients, and on health care staff.
2–4,7,8 Most demented patients display agitation at some point, and it is amenable to treatment in most, but not all, cases.
4Pharmacologic strategies for dealing with agitation in dementia include both neuroleptic and non-neuroleptic agents, but no singularly effective drug has been found—perhaps in part because agitation has several types and causes. Neuroleptics offer only moderate efficacy in treating agitation,
8–10 and their extrapyramidal and cognitive side effects are not negligible, making their long-term risk/benefit ratio uncertain.
11 Non-neuroleptic drugs, primarily anticonvulsants and antidepressants, are also widely used to treat agitation in AD,
7,10 but these drugs have been little studied systematically in this context.
10 Other drugs, such as propranolol, used to treat aggression in younger psychiatric populations, have been tried in dementia, but these drugs may have serious adverse side effects in the aged.
12There is much evidence that cholinergic deficiencies in AD make important contributions to the neuropsychiatric dimensions of the disease, including agitation.
13 Demented patients, for example, are highly vulnerable to side effects of anticholinergic psychotropics.
9 In view of the long-standing interest in cholinergic replacement therapies for palliating cognitive impairment in AD,
14,15 it will be important to examine the potential of cholinergic drugs to alleviate agitation. If cholinomimetics can be identified that lessen agitation significantly at clinical doses and with minimal untoward effects, it is possible that such drugs could supplement or reduce neuroleptic or non-neuroleptic medications given for this purpose. Further, certain cholinomimetics at appropriate doses might offer both agitation reduction and cognitive enhancement.
16There is reason to suspect that some cholinergics may act to abate agitation. It has long been postulated that a functional equilibrium exists in the normal striatum between cholinergic excitation and dopaminergic inhibition, and that when this equilibrium is disrupted by a loss of dopamine (DA) input, the motoric symptom complex of Parkinson's disease (PD), dominated by hypokinesia, tremor, rigidity, and abnormal posture, emerges.
17 More recently, it was proposed that DA neurons synapse on cholinergic interneurons in the striatum, inhibiting them, and that the acetylcholine released by these large interneurons is excitatory to striatal output neurons.
18 Studies in nonhuman primates have demonstrated that parkinsonian symptoms can result from pathologically decreased activity of the “direct” (pallidal-nigral-thalamocortical) and/or increased activity of the “indirect” (pallidal-subthalamic nuclear-thalamocortical) striatal output pathway.
19 Despite the use of anticholinergics to treat PD since the time of Charcot and the probable inhibitory action of DA on cholinergic interneurons, the action of cholinomimetics on striatal neurons has not received broad attention from neuropharmacologists. Although cognitive, presumably cortically mediated, effects of cholinergic agonism have been demonstrated in primate models,
14,20–22 the potential behavioral effects of newer, better tolerated cholinomimetics upon striatal neurons, particularly in the absence of DA deficiency, have not been evidenced. In our work with this class of cholinomimetic, we have noticed that high doses given to primates tend to quiet the animals down, an action that may occur through cholinergic modulation of the output to frontal cortex of the indirect circuit. Modulation of this circuit, moreover, would not be likely to lead to sedation. Since the various motor symptoms of PD result both from cholinergic excess and from DA depletion, it is also possible that proper cholinergic agents given to normodopaminergic subjects would reduce agitation without the emergence of other, undesired, parkinsonian symptoms.
The present pilot study tests, in a small number of subjects, the hypothesis that centrally active cholinomimetic drugs reduce motoric agitation in nonhuman primates. At present there is no generally accepted animal model of AD, and so our investigation was restricted to normal-young and normal-aged primates. It is, as such, intended only as groundwork for future approaches to evaluating pharmacotherapies for agitation in AD, based on the notion that a drug that reduces agitation in normal subjects may, but need not necessarily, also reduce agitation in AD. In this monkey behavioral model of agitation, one agent from each of the two major classes of therapeutic cholinomimetics was compared with vehicle-only. We examined tacrine (tetrahydroaminoacridine; THA), a reversible cholinesterase inhibitor,
23 and AF102B (cevimeline), a rigid acetylcholine analogue that acts as a partial selective direct M
1 muscarinic agonist in nervous tissue.
24 Both AF102B
24–27 and THA
20–22,28–33 have yielded cognitive improvement in animal models and in AD patients. Whereas THA is hepatotoxic,
31,34 known side effects of AF102B are minor at therapeutic doses.
35 AF102B also has certain in vitro properties that may offer additional benefits in treating AD.
36,37 To our knowledge, effects of THA and AF102B on agitation have not been studied previously.
METHODS
One male (age 7 years) and 4 female (16, 26, 28, and 33 years)
Macaca radiata (bonnet) monkeys in good health weighing 4–9 kg took part. Ages were taken from birth records. All monkeys were kept under controlled conditions and had received cognitive training prior to the present study. The same dark/light cycle, feeding time, and behavioral testing time (11:00
a.m.) were kept for all subjects. Senescence begins near age 20 years in the macaque;
38–40 thus, our population comprised 2 “young” and 3 “old” monkeys.
We developed the behavioral model of restlessness and agitation as follows: For earlier studies,
41 we had trained monkeys to execute computer tasks on a touchscreen monitor. Subjects habitually performed these tasks in exchange for fruit juice reward while sitting alone in a primate restraint chair inside a behavioral recording chamber. Ordinarily, after placement in the chamber, monkeys sat quietly awaiting the first computer task. When, however, subjects were left sitting without computer activity for more than 3 min, they became restless and agitated, with a marked increase in random, often repetitive, movements rather than any specific attempt to escape from the chair. This agitated state could persist for 15 min or longer. Informally, we have noticed this behavior repeatedly over months of working with the monkeys and have observed no apparent habituation or other change in its character over time.
We videotaped subjects during the first 5 min of these restless periods, starting 1 hour after administration of normal saline vehicle-only (control) and of each of four cholinomimetic drug-dose combinations. No systematic minute-to-minute variation in agitation was evident over this 5-min period. Nor were any time-dependent effects in drug response anticipated in this timeframe, as the cholinomimetics selected (AF102B and THA), in our experience, exert their behavioral effects on the timescale of hours rather than minutes. Therefore, for each monkey and each condition, a 3-min length of videotape was chosen at random out of the 5 min recorded and divided into 12 consecutive 15-s segments for viewing. Two judges watched each segment independently on a 21″ monitor at a distance of 1 m and counted every spontaneous movement made by the monkey during the segment. Judges were independent and blind to the drug conditions pertaining in each segment. Judges did, however, take part in a practice session, prior to formal judging, using an additional length of videotape, to establish protocol agreement on counting procedures. For each judge, monkey, and drug condition, a mean was taken across the twelve 15-s segments. We chose 15 s as segment length and 3 min as overall length because this minimized monitor viewer fatigue, a possible source of inaccuracy.
Videotaping began 60±5 min after administration of vehicle or drug, including a final 3-min wait during which the monkey sat alone before the inactive touchscreen. The variation in postadministration time was due to the usual small differences in the exact time required to chair the animal, to place the chair in the chamber, and so on, commonplace in monkey behavioral work.
Drugs were given in five separate regimens, each occurring 2 weeks apart and in the following order for all subjects:
1.
1 cc intramuscular (IM) normal saline (vehicle-only).
2.
0.5 mg/kg IM AF102B (young monkeys) or 0.2 mg/kg IM AF102B (old monkeys) (Lo-AF102B).
3.
3.5 mg/kg IM AF102B (young) or 2.0 mg/kg IM AF102B (old) (Hi-AF102B).
4.
Oral THA mixed in with 2 cc peanut butter vehicle: 1.0 mg/kg (young) or 0.5 mg/kg (old) (Lo-THA).
5.
3.0 mg/kg (young) or 2.0 mg/kg (old) oral THA in 2 cc peanut butter (Hi-THA).
In our previous published
41–43 and unpublished experience, we have seen AF102B yield cognitive enhancement in the 0.1–4.5 mg/kg dose range for young monkeys, with best enhancement around 1.1 mg/kg, whereby both the dose ranges and the best doses vary for individual subjects. For old monkeys on AF102B, the range has been 0.1–0.6 mg/kg (best dose near 0.3 mg/kg). For THA, the ranges have been 0.5–2.5 mg/kg (best dose near 1.3 mg/kg) for young monkeys and 0.5–2.0 mg/kg (best dose near 1.0 mg/kg) for old monkeys. For comparison, daily oral doses of THA ranging from 0.5 to 2.3 mg/kg are recommended for AD patients.
44 Comparable human dosage data are not available for AF102B. Cognitive enhancement in monkeys was previously assessed as improvements in behavioral performance, including increases in accuracy and decreases in reaction time in object working memory
41,43 and visual focused attention
42 tasks.
We used the Pearson correlation coefficient (r) to compare the results of the two judges. Drug effects were tested in separate repeated-measures analyses of variance (R-ANOVA) for each agent, using differences between movement rates on drug and on vehicle-only across all 5 subjects, with dose (Lo vs. Hi) as a factor. R-ANOVAs were done both for differences in absolute movement rates and for percentage differences in movement rates normalized to rates on vehicle-only. One-way analyses of variance (1-ANOVA) were used for direct comparisons between the young and the old monkeys. Absolute and percentage differences between drug and vehicle were also examined for both drugs at both doses with one-sided t-tests to see if means across monkeys were significantly below zero. Criterion for statistical significance was P<0.05 in this exploratory, pilot study.
RESULTS
For most subjects, reduced restlessness and motoric agitation were readily apparent on segments taped after cholinergic administrations relative to segments taped after vehicle administrations. Both frequency and amplitude of spontaneous movements were clearly decreased even on superficial inspection. Movement amplitude was not further investigated, but the finding of reduced frequency was tested objectively by the 2 blind judges.
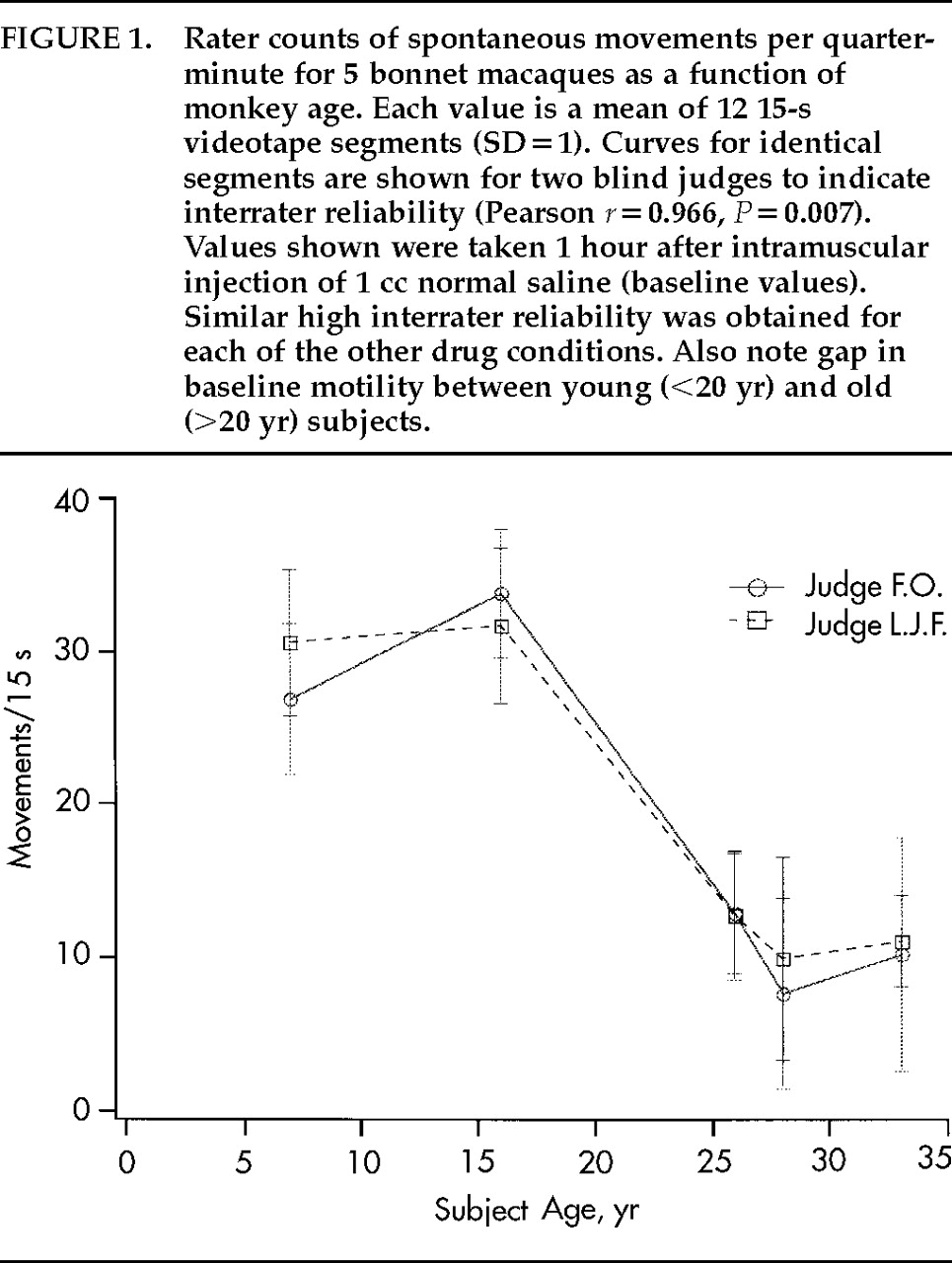
Figure 1 plots average spontaneous movements per 15-s segment counted by each of the 2 judges for each of the 5 monkey subjects under the vehicle-only condition. The curves for the 2 judges overlap well (
r=0.966,
P<0.05). Similar correlations between the 2 judges were found for the other four conditions (not shown). Therefore, mean values for the 2 judges are cited below.
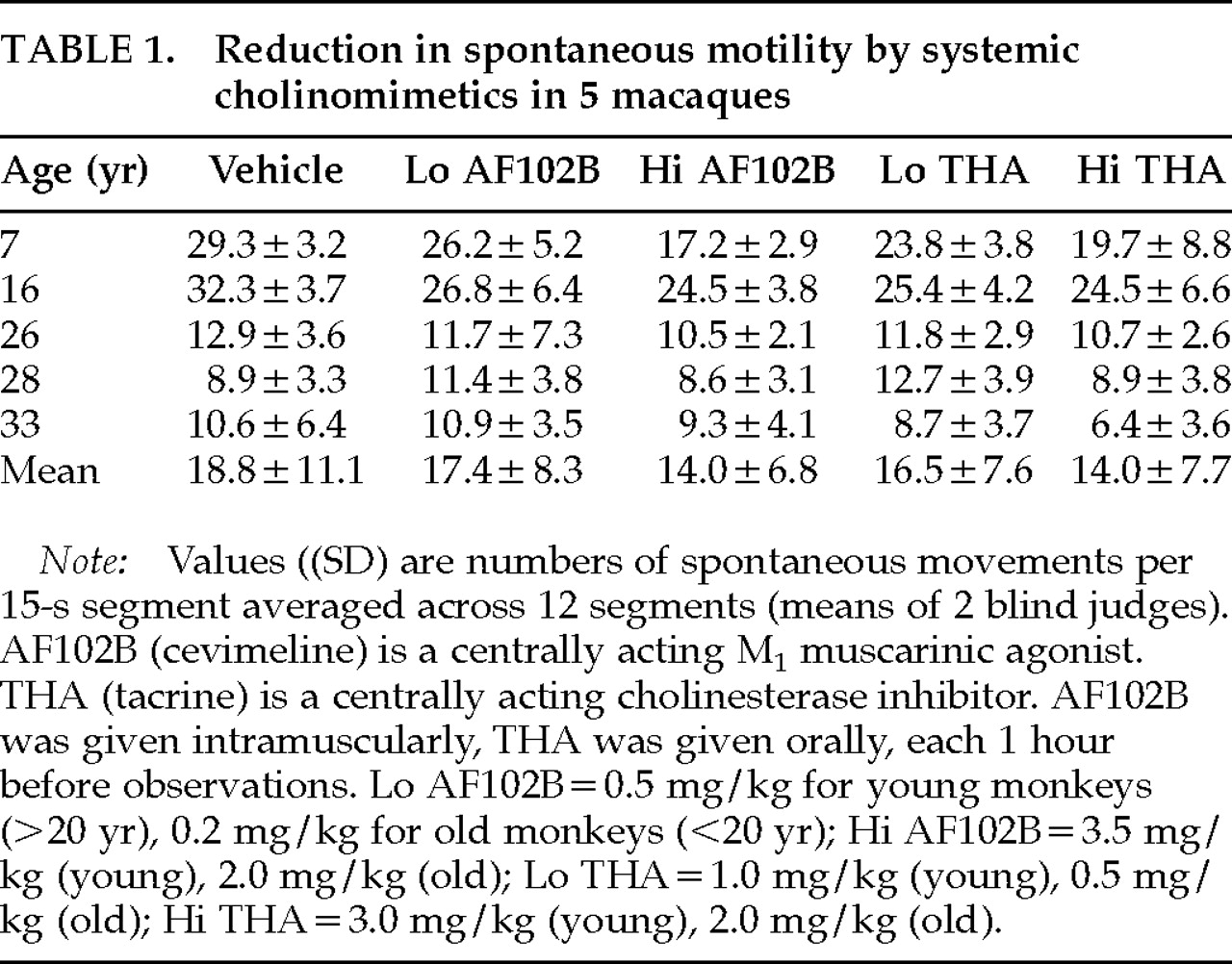
Mean spontaneous movements per quarter-minute are listed for each monkey under each drug condition in
Table 1. The 2 young monkeys had significantly higher motility than the 3 old monkeys under all drug conditions (e.g., 1-ANOVA for vehicle:
F=114.65, df=4,
P=0.002). Low-dose AF102B was associated with movement reduction for 3 of 5 monkeys. Movement decrement, slight for the old monkeys, was recorded for 5 of 5 monkeys on high-dose AF102B. Movement reduction was found for 4 of 5 monkeys on both low- and high-dose THA. The 28-year-old monkey responded poorly to both doses of both drugs. Comparing dose-responses within drugs, spontaneous movement rates were lower at high-dose than at low-dose for both cholinomimetics for all monkeys. In R-ANOVA computed on differences between movements per 15 s on drug and movements per 15 s on vehicle, the effect of dose of AF102B did not reach significance for differences in absolute numbers (
F=5.57, df=1,4,
P=0.078), but did become significant when values were expressed as percentage differences (
F=13.00, df=1,4,
P<0.05). For THA, the opposite applied: the effect of dose of THA was significant for absolute numbers (
F=13.54, df=1,4,
P<0.05), but not for percentage differences (
F=6.70, df=1,4,
P=0.061). One-sided
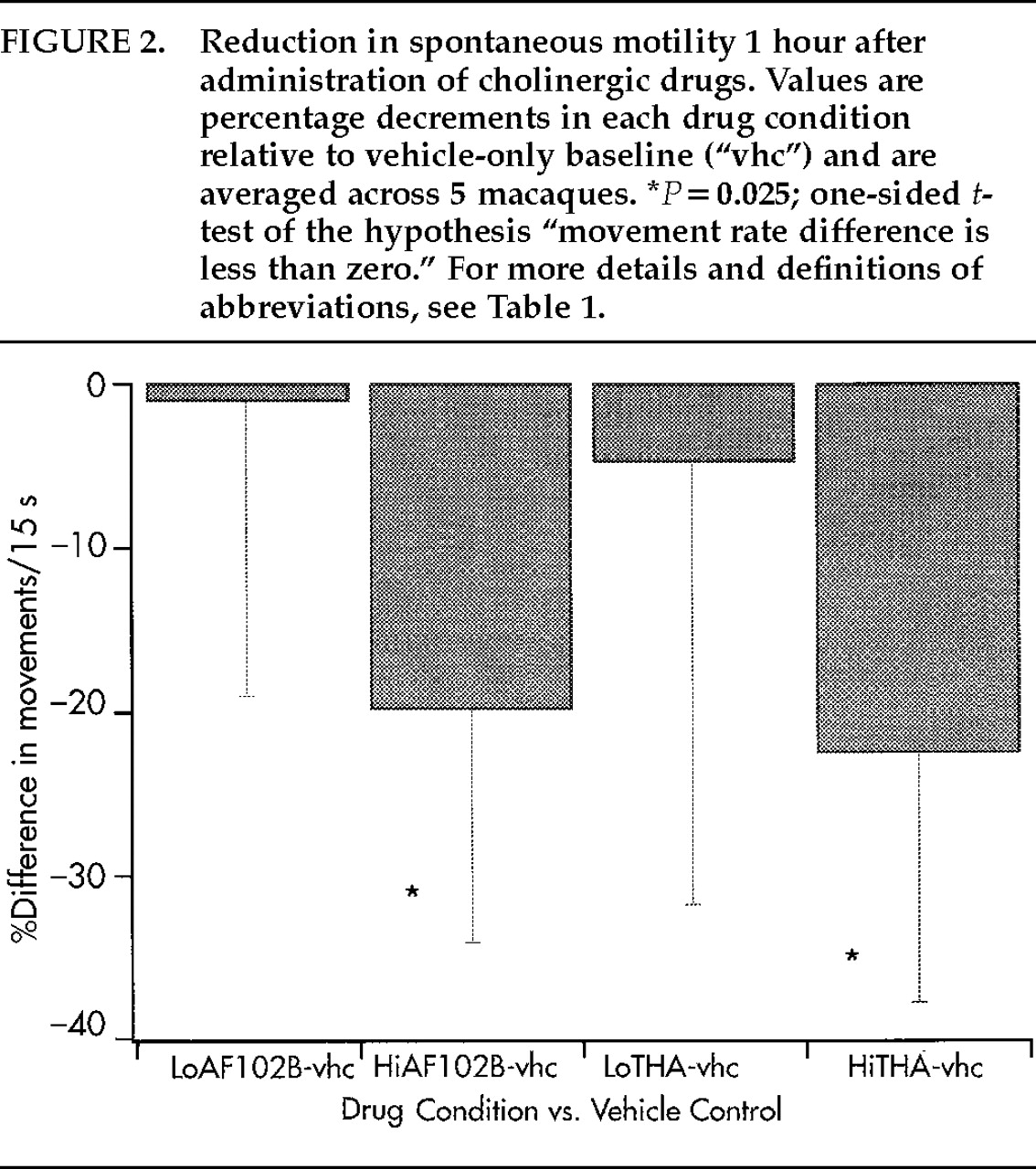
t-tests across all 5 monkeys (
Figure 2) indicated movement-rate drops to levels significantly below control for Hi-AF102B and for Hi-THA (for both conditions: absolute numbers
P<0.05, percent differences
P=0.025), but not for Lo-AF102B or for Lo-THA (
P>0.10 for both measures and both conditions). Tremor, rigidity, and abnormal posture were not visible; nor were other side effects, in particular sedation and hypercholinergic signs, noted for any monkey for either drug at any dose.
DISCUSSION
The high interrater correlation on the movement frequency measure suggests that results recorded are reliable and that the videotape technique may represent a practical means of documenting and quantifying models of primate agitation. As the form of agitation commonly found in dementia features frequent purposeless stereotypies such as “hand-wringing, foot-tapping, picking,”
45 behaviors akin to those observed in our restless, partially restrained monkeys, the present model may have some relevance for the study of agitation in dementia.
Significant decrements in spontaneous nonpurposive movement rates of nonhuman primates were found in response to the cholinesterase inhibitor THA and the M
1 muscarinic agonist AF102B, at clinically nontoxic doses. Although the study has a small population sample, and we therefore can claim little in terms of statistical power, the presence and relative uniformity of cholinergic-associated movement decrement across subjects suggest that a notable effect may be present. Consequently, we think further studies on larger numbers of animals and in agitated dementia patients are needed. We postulate that dampening of spontaneous, nonpurposive movements (restlessness, agitation) may be exerted through cholinergic action in the striatum
46 in the absence of DA deficiency (nonparkinsonian subjects) and may be mediated through indirect pathway connections with frontal cortices elaborating movement.
47A high dose and a low dose of each drug were given to each subject. For both cholinomimetics, high doses were more effective than low doses in reducing movement, even though low doses were closer to previously observed best doses for cognitive task performance, and we therefore expect greater cognitive enhancement at low than at high doses. This dissociation between agitation-reducing and cognition-enhancing doses may be due to a receptor population–related differential sensitivity between cortex and striatum. Ideally, maximal agitation reduction would occur at doses yielding best cognitive enhancement. But, given the likelihood that cognitive functions and motoric agitation are mediated by somewhat independent (e.g., cortical versus subcortical) mechanisms in the primate brain, such coincidence of doses seems unlikely. Therefore, a cognitive benefit might attend in some, but not all, AD patients who were treated with AF102B or THA for agitation.
AF102B and THA were each given 1 hour before videotaping. The aforementioned enhancements in macaque memory and attention
41–43 were also recorded 1 to 2 hours after dosing with AF102B or THA. In future studies, it might be useful to examine agitation at later times postdrug, since, for example, the THA metabolite velnacrine still improves cognitive function as late as 24 hours after ingestion.
48 Because of the small number of subjects, the vehicle, AF102B, and THA were given in the same order to all subjects in the present study, introducing a potential bias from order effects. We have not detected order effects in our past monkey behavioral work with THA and AF102B,
41 but this question nonetheless should be examined more systematically.
In the present study, as well as in previous work with chronic administration of AF102B and THA in macaques,
41 we have only rarely noted externally visible adverse effects at doses tested. We cannot comment on possible internal organ pathology induced by these drugs, since we did not test for it. Importantly, in the present study reduction in agitation was observed without obvious tremor, rigidity, or abnormal posture at the doses tested. One explanation of this result might be that elevated central cholinergic agonism leads to hypokinesia, but not to other parkinsonian symptoms, as long as normal DA levels are maintained. Motility was also lowered in the absence of sedation. This finding is consistent with our previous work,
49 in which we found that EEG slow-wave production was decreased in the macaque in response to central cholinergics. Therefore, further research with cholinomimetics is recommended, since reduction of agitation, with possible cognitive enhancement, was achieved at doses not inducing parkinsonoid or other systemic side effects.
ACKNOWLEDGMENTS
This work was supported by the U.S. Department of Veterans Affairs Merit Review Program. It was previously presented at the Fourth International Conference for Progress in Alzheimer's and Parkinson's Diseases, Tel Aviv, Israel, May 14–17, 1997.