Cognitive deficits constitute a core feature of schizophrenia (
1), are persistent across the course of the illness (
2), and are the best predictor of long-term functional outcome (
3). At least some of these deficits arise during childhood and adolescence, before the onset of the psychosis associated with a clinical diagnosis of schizophrenia (
4).
The impairments in certain cognitive processes, such as working memory, appear to reflect alterations in specific elements of dorsolateral prefrontal cortex (DLPFC) circuitry known to be critical for working memory in primates (
5). For example, reciprocal excitatory connections among DLPFC layer 3 pyramidal cells are thought to mediate sustained neuronal activity during the maintenance phase of working memory in monkeys (
6); in individuals with schizophrenia, these neurons have fewer dendritic spines, the principal site of glutamatergic inputs (
7). In contrast, the psychotic symptoms of schizophrenia are associated with excessive dopamine release in the associative striatum (
8).
These two domains of schizophrenia pathology and pathophysiology have been hypothesized to be related via the excitatory projections from the DLPFC to the ventral mesencephalon, the location of the dopamine cells that project to the striatum (
9). That is, the deficit in dendritic spines, and the resulting reduction in excitatory drive to DLPFC pyramidal cells, is thought to lead to the hyperdopaminergic state subcortically (
10). Consistent with this hypothesis, conditional deletion in mice of the actin-related protein-2/3 (ARP2/3) complex, which acts as a common final pathway for multiple signaling cascades that regulate the actin cytoskeleton required for dendritic spine formation and maintenance (
11,
12), was shown to produce a loss of dendritic spines in prefrontal cortical pyramidal cells and elevated striatal dopamine neurotransmission (
13). These changes were accompanied by cognitive deficits and antipsychotic-responsive locomotor hyperactivity (
13,
14). The idea that a spine deficit in prefrontal cortical pyramidal neurons was the upstream cause of increased subcortical dopamine was supported by proof-of-concept evidence that viral re-expression of the ARP2/3 complex in frontal cortical pyramidal neurons lowers striatal dopamine levels and reduces locomotor hyperactivity (
13).Together, these findings provide a mechanistic basis for the prior observation that diminished activity in the DLPFC predicts subcortical hyperdopaminergia in schizophrenia (
15). However, it is not known whether expression of the ARP2/3 complex in DLPFC pyramidal cells is deficient in schizophrenia.
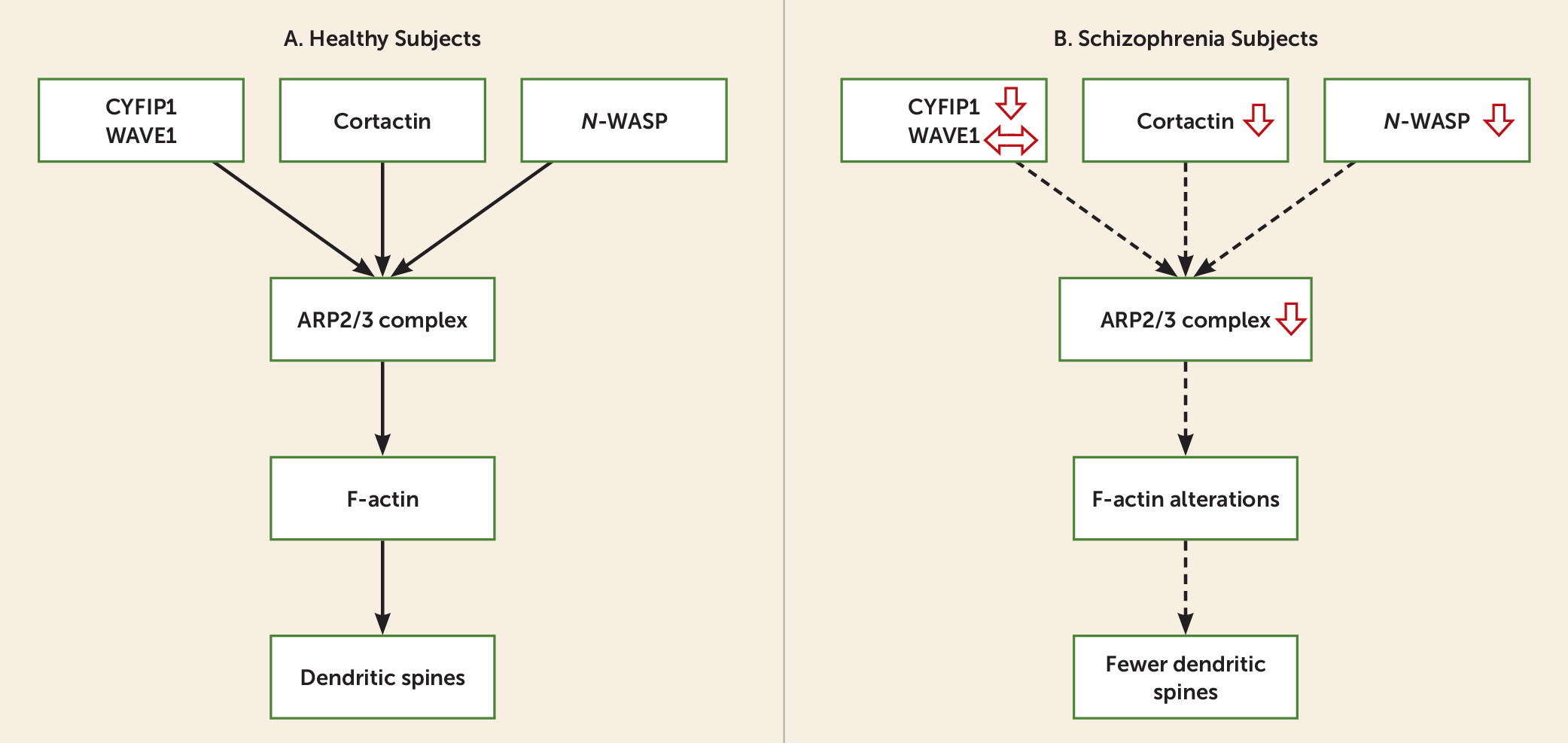
The goal of the present study was to examine the ARP2/3 complex signaling pathway (
Figure 1A) in DLPFC deep layer 3 pyramidal cells, where dendritic spine alterations are most pronounced in schizophrenia (
16,
17). We used laser microdissection to capture individual pyramidal cells in DLPFC deep layer 3 from subjects with schizophrenia and matched unaffected comparison subjects and assessed gene expression in pools of neurons from each subject by microarray analysis. In order to assess the cell-type specificity of the findings, we analyzed microarray data from layer 5 pyramidal cells and layer 3 parvalbumin interneurons, and we measured expression levels of certain transcripts in total DLPFC gray matter using quantitative reverse-transcription polymerase chain reaction (qPCR). In each analysis, we evaluated transcript levels for all subunits of the ARP2/3 complex and of nucleation promotion factors that regulate the activity of the ARP2/3 complex (
Figure 1A).
Method
Human Subjects
Brain specimens (N=124) were obtained during routine autopsies conducted at the Allegheny County Office of the Medical Examiner (Pittsburgh) after consent was obtained from next of kin (
18). Subject groups for the entire cohort (N=62 pairs; for individual subject details, see Table S1 in the
data supplement that accompanies the online edition of this article), as well as for the subsets of pairs used in specific studies, did not differ significantly in mean age, postmortem interval, RNA integrity number (Agilent Bioanalyzer, Santa Clara, Calif.), tissue storage time at −80°C, or race (
Table 1). Mean brain pH was significantly different between subject groups for the entire cohort (t=2.6, df=122, p=0.01), but the difference was quite small (0.1 pH unit) and of uncertain biological significance. All procedures were approved by the University of Pittsburgh’s Committee for the Oversight of Research and Clinical Trials Involving the Dead and Institutional Review Board for Biomedical Research.
Laser Microdissection Procedure
As described previously (
19,
20), cryostat sections (12 μm) containing DLPFC area 9 were thaw-mounted onto glass PEN membrane slides (Leica Microsystems, Bannockburn, Ill.) and stained for Nissl substance with thionin. Using a Leica laser microdissection system, DLPFC deep layer 3 and layer 5 were identified in portions of the section cut perpendicular to the pial surface. Nissl-stained pyramidal neurons, identified by their characteristic triangular shape and prominent apical dendrite, were individually dissected from each laminar location. Parvalbumin interneurons were identified by dual labeling of NeuN-positive interneurons and
Vicia villosa agglutinin (VVA) labeling of perineuronal nets in layers 3 and 4 (
21).
Microarray Analyses
For pyramidal cell microarray analyses (
19,
21), 200 DLPFC pyramidal cells from deep layer 3 and from layer 5 were dissected individually and then pooled together by layer for each subject (N=36 pairs) (
19). Array results were not readable for layer 5 samples from two schizophrenia subjects, and thus two subject pairs were excluded. For parvalbumin interneuron microarray analyses, 360 parvalbumin interneurons were dissected from 14 of the 36 subject pairs (
21). The cDNA from each sample was loaded on an Affymetrix GeneChip HT HG-U133
+ PM Array plate (Affymetrix, Santa Clara, Calif.) designed to assess transcript levels from the human genome.
We also used microarray analyses to evaluate ARP2/3 complex signaling pathway transcripts in DLPFC deep layer 3 pyramidal cells from monkeys with chronic exposure to olanzapine, haloperidol, or placebo (see the Supplementary Methods section in the online data supplement).
qPCR
For each subject in the full cohort (N=62 pairs), area 9 gray matter was collected and cDNA prepared as previously described (
18) (see the Supplementary Methods section in the
data supplement). Based on their stable levels of expression across schizophrenia and comparison subjects, three reference genes (beta-actin, cyclophilin-A, and glyceraldehyde-3-phosphate dehydrogenase [GAPDH]) were used to normalize target mRNA levels. The difference in cycle threshold (dCT) for each target transcript was calculated by subtracting the geometric mean CT of the three reference genes from the CT of the target transcript. Since dCT represents the log
2-transformed expression ratio of each target transcript to the reference genes, the relative level of the target transcript for each subject is reported as 2
−dCT.
Data Analysis
A comprehensive description of the statistical analyses used for the microarray data is provided by Arion et al. (
19). Briefly, Affymetrix CEL files were normalized and log
2-transformed using RMA Express. After filtering to remove uninformative probe sets, the random intercept model with Bayesian information criterion variable selection (RIM-BIC) was used to analyze the microarray data set. An adaptively weighted Fisher’s method was used to combine the differential expression information for each transcript, and the Benjamini-Hochberg protocol was used to control the false discovery rate (<0.05) for the meta-analyzed p values.
In a separate analysis, the microarray signals for all probe sets targeting each ARP2/3 complex transcript were averaged within each sample and used in two analysis of covariance (ANCOVA) models. The paired ANCOVAs used the level of each mRNA as the dependent variable, diagnostic group as the main effect, subject pair as a blocking factor, and postmortem interval, storage time, brain pH, and RNA integrity number as covariates. However, subject pairing may be considered an attempt to account for the parallel processing of tissue samples from a pair and to balance diagnostic groups for sex and age, and thus not to be a true statistical paired design. Consequently, we also used an unpaired ANCOVA that included age, sex, postmortem interval, storage time, brain pH, and RNA integrity number as covariates. These same two models were used to analyze the data from the qPCR analyses. Because the two models gave the same results with regard to statistical significance, only the results from the unpaired model are presented here (for the paired model results, see Table S3 in the online data supplement).
The influence of comorbid factors on ARP2/3 complex signaling pathway transcript expression was assessed by ANCOVA (see the Supplementary Methods section in the data supplement).
For all ANCOVAs, reported statistics include only the covariates that were statistically significant. As a result, the reported degrees of freedom vary across analyses.
Results
Expression Levels of ARP2/3 Complex Signaling Pathway Genes in DLPFC Deep Layer 3 Pyramidal Cells in Schizophrenia
Using a false discovery rate of 5%, seven of the 12 transcripts in the ARP2/3 complex signaling pathway were significantly down-regulated in DLPFC deep layer 3 pyramidal cells from schizophrenia subjects, and another three transcripts showed a nonsignificant (p<0.09) down-regulation (
Table 2). Because a number of transcripts were represented on the Affymetrix array by more than one probe set, the microarray signal for redundant probe sets was averaged within each sample. The resulting mRNA levels were compared between subject groups by ANCOVA as described in the following paragraphs.
The ARP2/3 complex, which stimulates de novo actin nucleation and polymerization to generate F-actin branched filament networks that modulate spine morphogenesis, is composed of seven subunits (
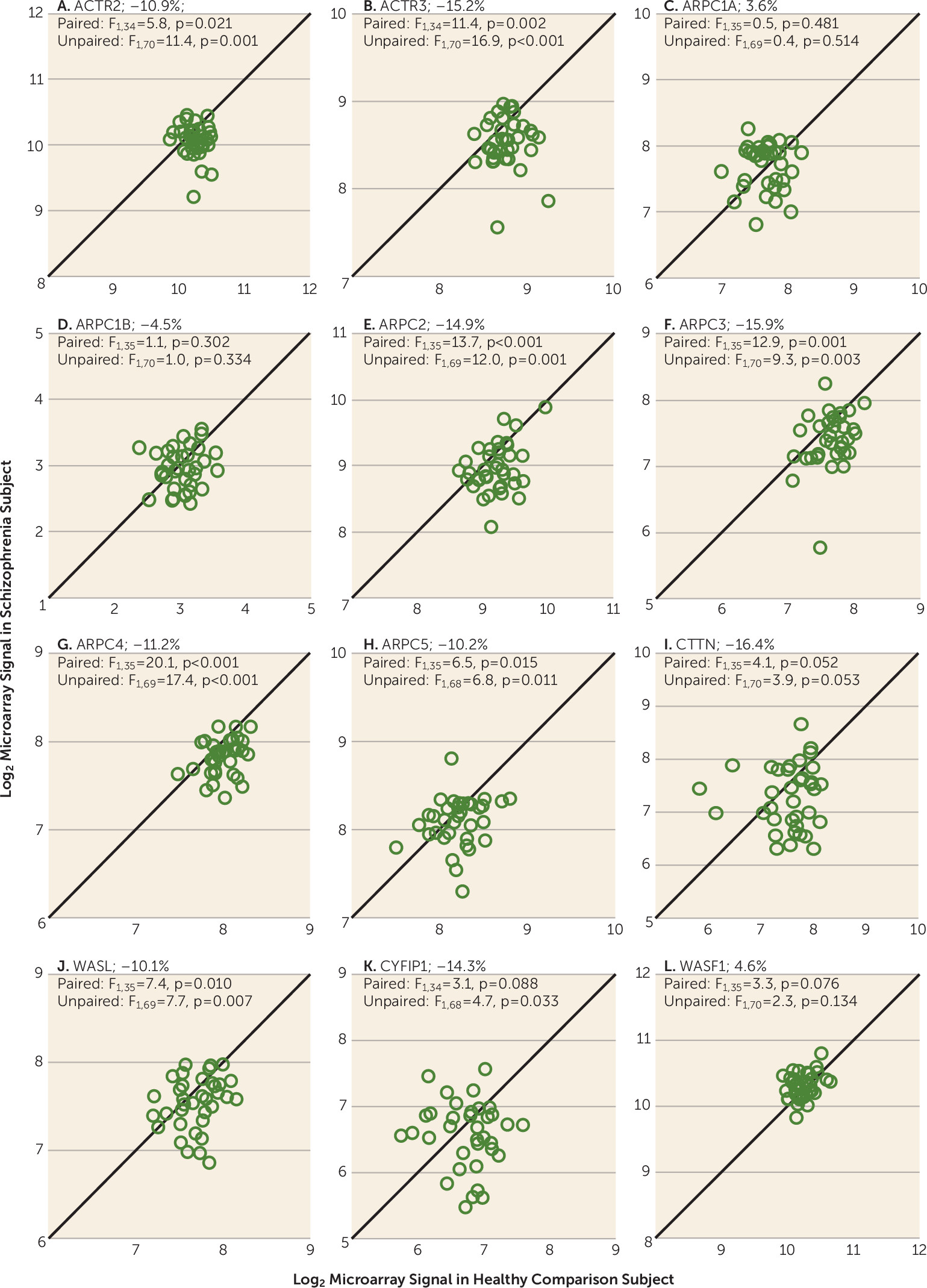
11). The ACTR2 and ACTR3 transcripts encode the ARP2 and ARP3 subunits, which function as the ATP-binding component of the complex. In layer 3 pyramidal cells, mean transcript levels were significantly lower for both ACTR2 (−10.9%; F=11.4, df=1, 70, p=0.001) and ACTR3 (−15.2%; F=16.9, df=1, 70, p<0.001) in the schizophrenia subjects (
Figure 2). The five ARPC subunits of the ARP2/3 complex are critical for actin binding and nucleation. In subjects with schizophrenia, mean transcript levels were lower for ARPC2 (−14.9%; F=12.0, df=1, 69, p=0.001), ARPC3 (−15.9%; F=9.3, df=1, 70, p=0.003), ARPC4 (−11.2%; F=17.4, df=1, 69, p<0.001), and ARPC5 (−10.2%; F=6.8, df=1, 68, p=0.011). The mean levels of the two human isoforms of ARPC1 (ARPC1A and ARPC1B) did not differ significantly between subject groups.
The ARP2/3 complex is regulated by nucleation promotion factors, including cortactin (CTTN), Neural Wiskott-Aldrich syndrome (
N-WASP) proteins, and WASP family Verprolin-homologous (WAVE) proteins, such as WAVE1, which is modulated by cytoplasmic FMR1 interacting protein 1 (CYFIP1) (
11,
22). In layer 3 pyramidal cells, mean CTTN transcript levels were lower in schizophrenia subjects, although this difference did not achieve statistical significance (−16.4%; F=3.9, df=1, 70, p=0.053). Mean transcript levels for WASL, which encodes
N-WASP, were lower in schizophrenia (−10.1%; F=7.69, df=1, 69, p=0.007). For the WAVE complex, mean CYFIP1 transcript levels were lower in schizophrenia (−14.3%; F=4.7, df=1, 68, p=0.033), whereas mean transcript levels for WAVE1 (encoded by WASF1) were unaltered (4.6%; F=2.3, df=1, 70, p=0.134).
Effects of Comorbid Factors on ARP2/3 Complex Signaling Pathway Transcripts
Levels of ARP2/3 complex signaling pathway mRNAs that differed between subject groups did not differ among the schizophrenia subjects as a function of diagnosis of schizoaffective disorder; history of substance dependence or abuse; use of nicotine, antipsychotics, antidepressants, or benzodiazepines and/or sodium valproate at the time of death; or death by suicide (see Figure S1 in the online data supplement), with the exception that schizophrenia subjects with a history of substance dependence or abuse had significantly lower ARPC4 (F=16.74, df=1, 34, p=0.001) and ARPC5 (F=5.96, df=1, 33, p=0.020) mRNA levels relative to those without such a history.
Effects of Antipsychotic Exposure on ARP2/3 Transcripts in Monkeys
Transcript levels for ARP2/3 complex signaling pathway in DLPFC layer 3 pyramidal cells did not differ significantly among monkeys chronically exposed to olanzapine, haloperidol, or placebo (see Figure S2 in the online data supplement).
Cellular Specificity of Altered Expression ARP2/3 Complex Signaling Pathway Transcripts in Schizophrenia
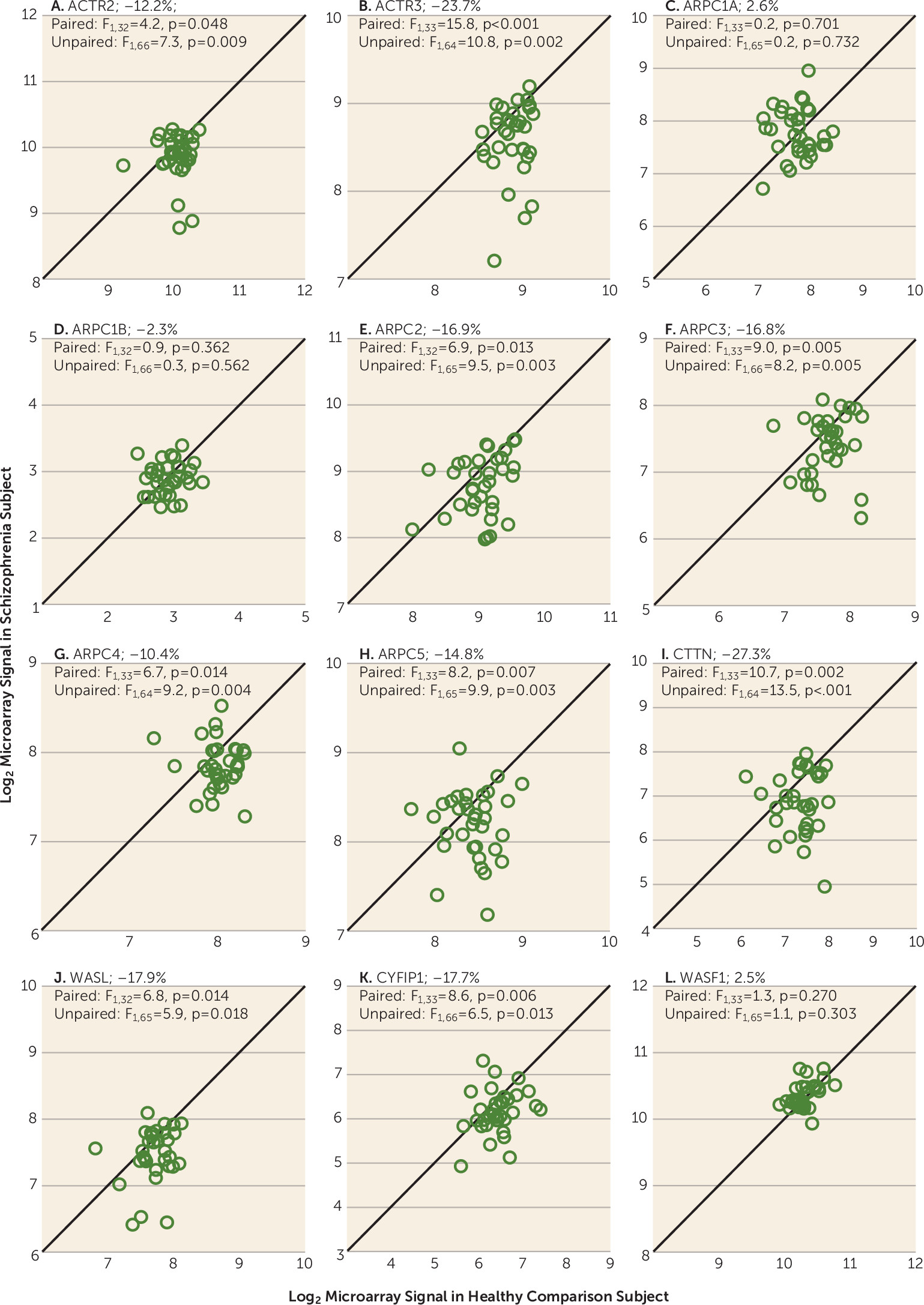
To determine whether these transcript alterations in the ARP2/3 complex signaling pathway were also present in other cell types, we conducted similar microarray analyses in DLPFC layer 5 pyramidal cells. Using a false discovery rate of 5%, each probe set in the ARP2/3 complex signaling pathway showed the same pattern of down-regulation in both layer 5 and layer 3 pyramidal cells in schizophrenia; probe sets for two transcripts (CTTN and CYFIP1) that only approached statistical significance in layer 3 were clearly significantly lower in layer 5 pyramidal cells (
Table 2). Consistent with these findings, ANCOVA analyses revealed lower mean mRNA levels for the ATP-binding component of the complex (ACTR2 and ACTR3) and for the subunits that mediate actin binding and nucleation (ARPC2, ARPC3, ARPC4, and ARPC5) (
Figure 3). In addition, mean mRNA levels of the nucleation promotion factors (CTTN, WASL, and CYFIP1) were also lower (
Figure 3).
We also evaluated expression of transcripts in parvalbumin interneurons in a subset of subject pairs using data from an existing microarray data set (
21). With the exception of ARPC3 mRNA, which was significantly lower in parvalbumin interneurons (−51.5%; F=7.4, df=1, 26, p=0.012), mean mRNA levels of ARP2/3 complex signaling pathway transcripts in layer 3 parvalbumin neurons did not differ significantly between subject groups. These findings do not appear to be false negatives due to a smaller sample size, because in these same 14 subject pairs, multiple transcripts were significantly down-regulated in layer 3 pyramidal cells in schizophrenia: ACTR2 (−12.7%; p=0.04), ACTR3 (−20.0%; p=0.002), ARPC2 (−21.3%; p=0.003), ARPC4 (−13.8%; p=0.009), and WASL (−14.5%; p=0.019); levels of two other transcripts were also lower but did not achieve statistical significance: ARPC3 (−18.2%; p=0.073) and CTTN (−24.6%; p=0.062). These findings suggest that within the same cortical layer, most alterations in the ARP2/3 complex signaling pathway are specific to, or at least enriched in, pyramidal cells relative to parvalbumin interneurons. However, caution is warranted in interpreting these findings given the technical caveat that the most severely affected parvalbumin interneurons may not have been identifiable for capture by laser microdissection (
21).
ARP2/3 Complex Signaling Pathway Transcripts in DLPFC Gray Matter in Schizophrenia
To further test the cell-type specificity of alterations in the ARP2/3 complex signaling pathway, we analyzed transcript levels in total DLPFC gray matter by qPCR (see Table S3 in the online data supplement) in the same 36 pairs of subjects used for the pyramidal cell profiling. In contrast to the findings in pyramidal cells, mean mRNA levels in the schizophrenia subjects were higher for ARPC3 (3.8%; F=3.2, df=1, 69, p=0.034), CTTN (5.9%; F=7.6, df=1, 67, p=0.007), and WASL (3.4%; F=6.5, df=1, 70, p=0.013). None of the other transcripts significantly differed between subject groups, with the exception of mRNA levels for ARPC5, which were significantly lower (−6.0%; F=5.4, df=1, 68, p=0.023), but this decrement was smaller than in pyramidal cells.
To control for possible false negative findings due to sample size, the transcripts in the ARP2/3 complex signaling pathway were evaluated in DLPFC gray matter in a larger subject cohort (N=62 pairs). Of the nine transcripts that were significantly lower in layer 3 and/or 5 pyramidal neurons in subjects with schizophrenia, three were increased in expression, three were not different, and three were decreased, but the deficits were much smaller than in pyramidal cells (see Table S3).
Discussion
Our results identify lower mRNA levels of multiple subunits of the ARP2/3 complex, and of nucleation promotion factors that regulate the activity of the ARP2/3 complex, in DLPFC deep layer 3 and 5 pyramidal cells in schizophrenia (
Figure 1B). These alterations appear to reflect the disease process of schizophrenia, as they were not attributable to chronic treatment with antipsychotic medications, other factors frequently comorbid with schizophrenia, or potential confounders. In contrast, expression levels of ARP2/3 complex signaling pathway transcripts were up-regulated, not altered, or only modestly lower in DLPFC parvalbumin interneurons and DLPFC gray matter. Furthermore, the expression deficits in ARP2/3 complex signaling pathways in pyramidal cells do not appear to reflect nonspecific factors, as multiple other transcripts are up-regulated in these neurons (
19,
23). Because the ARP2/3 complex signaling pathway is a critical determinant of F-actin nucleation and polymerization, dysregulation of the ARP2/3 complex signaling pathway in schizophrenia may contribute to actin cytoskeleton impairments and spine deficits in DLPFC pyramidal cells.
Predicted Consequences of Down-Regulated ARP2/3 Complex Signaling Pathway in DLPFC Pyramidal Cells
In DLPFC pyramidal cells from schizophrenia subjects, the down-regulation in mRNA levels for multiple ARP2/3 complex transcripts (e.g., ACTR2, ARPC2, ARPC3) would be predicted to contribute to spine deficits by suppressing de novo actin polymerization, which generates the branched actin filament networks required for spine formation (
Figure 1B) (
24–
26). In addition, high concentrations of the ARP2/3 complex are found in the spine head, where it is localized proximal to F-actin filaments (
27). Therefore, reduced gene expression of ARP2/3 complex subunits would likely impair the actin nucleation and cross-linking of F-actin filaments, which are critical for the activity-dependent structural plasticity of spines, contributing to decreased spine stability and ultimately spine loss.
The ARP2/3 complex also acts as a molecular hub downstream of several signaling pathways that promote the structural stabilization of F-actin filaments necessary for spine maintenance (
28–
31). For example, nucleation promotion factors are required to activate the intrinsically inactive ARP2/3 complex; specifically, the nucleation promotion factors
N-WASP, cortactin, and CYFIP1 bring the ARP2/3 complex and F-actin monomers together to initiate the synthesis of new F-actin filaments (
32). Therefore, the down-regulation of these nucleation promotion factors in schizophrenia may further diminish ARP2/3 complex activity in the following ways (
Figure 1B).
First, activation of ARP2/3 complex by
N-WASP favors branch nucleation, which allows stabilization of newly synthesized F-actin filaments (
33,
34); thus, lower levels of both
N-WASP and the ARP2/3 complex in schizophrenia would likely result in reduced nucleation of spine precursors such as lamellipodia and filopodia. Indeed, knockdown of endogenous
N-WASP by RNA interference has been shown to decrease dendritic spine number in hippocampal pyramidal neurons (
34).
Second, CYFIP1 acts through the WAVE regulatory complex and cortactin to regulate the actin-nucleating activity of the ARP2/3 complex (
11,
22,
35); thus lower levels of CYFIP1, cortactin, and ARP2/3 complex subunits would further impair F-actin filament turnover and actin cytoskeleton dynamics, which are necessary for formation of dendritic spines.
Third, CYFIP1 inhibits dendritic protein translation in an activity-dependent fashion and promotes actin cytoskeleton remodeling (
29) by acting downstream of brain-derived neurotrophic factor (BDNF) and its postsynaptic partner TrkB, which are crucial for spine enlargement and stabilization (
36). Thus, the lower levels of both BDNF and TrkB in the DLPFC of subjects with schizophrenia (
37,
38) would likely further exacerbate the negative impact of impaired ARP2/3 complex signaling on spine number in DLPFC pyramidal cells.
Laminar-Specificity of Spine Pathology in DLPFC Pyramidal Cells in Schizophrenia
Previous postmortem studies have revealed lower basilar dendritic spine density on deep layer 3, but not on layer 5 or 6 pyramidal neurons, from the same subjects with schizophrenia (
16,
17). Given this apparent laminar specificity of spine deficits, we expected that alterations in the ARP2/3 complex signaling pathway would be more marked in deep layer 3 than layer 5 pyramidal cells. However, the presence of ARP2/3 complex signaling pathway alterations in pyramidal cells in both layers suggests that these deficits 1) are not a secondary consequence of a reduced number of pyramidal cell dendritic spines and 2) may be a necessary but not a sufficient cause of reduced spine density.
The prominence of spine deficits in deep layer 3 pyramidal cells may be due to other factors upstream of the ARP2/3 complex, such as disturbances in certain components of the Rho family GTPase cell division cycle 42 (CDC42) signaling pathway (
23,
39) for the following reasons. First, some of the components of this pathway, such as CDC42 effector proteins (CDC42EPs), are preferentially expressed in layers 2–3 of the DLPFC (
40) and the CDC42-CDC42EP pathway is dysregulated in DLPFC layer 3 pyramidal cells in schizophrenia (
23,
39). Second, another CDC42 signaling pathway (CDC42-p21-activated serine/threonine protein kinases [PAK]-LIM domain-containing serine/threonine protein kinases [LIMK] signaling pathway) is also altered in DLPFC layer 3 pyramidal cells; these alterations could destabilize actin dynamics by reducing F-actin turnover through cofilin, a family of actin depolymerizing proteins (
23). Third, in addition to disrupting the spine cytoskeleton in their own right, these impairments in CDC42 signaling could magnify the impact of the alterations in the ARP2/3 complex signaling pathway. For example, the intrinsically autoinhibited
N-WASP is directly activated by CDC42 (
41,
42); thus, deficits in CDC42 signaling could exacerbate the consequences of deficits in the
N-WASP-ARP2/3 complex cascade. In summary, the prominence of spine deficits in DLPFC deep layer 3 pyramidal cells in schizophrenia may reflect the convergent effects of alterations in several different signaling pathways, each of which could destabilize the actin cytoskeleton.
Do Actin Cytoskeleton Impairments Reflect Genetic Liability for Schizophrenia?
The altered expression of genes in the CDC42 and ARP2/3 signaling pathways may be related to genetic risk factors for schizophrenia. For example, the interaction of single-nucleotide polymorphisms in CYFIP1, ARP2, and ARP3 has been reported to be associated with an increased risk for schizophrenia (
43). Genome-wide association studies in schizophrenia have revealed enrichment of risk variants in genes whose products are involved in the activity-regulated cytoskeleton-associated scaffold protein (ARC), postsynaptic density (PSD) protein complex, and complement component 4 (C4) genes, which are heavily localized to dendritic spines and neuronal synapses (
44–
47). Aberrant expression of these gene products during postnatal development may exacerbate spine pruning and synapse loss in subjects with schizophrenia. In addition, de novo mutations in schizophrenia are overrepresented among loci encoding cytoskeleton-associated proteins that regulate actin (
48). These genetic findings are consistent with the idea that abnormalities intrinsic to DLPFC deep layer 3 pyramidal cells represent an “upstream” component in the disease process of schizophrenia (
49,
50). That is, variants or mutations in genes that regulate the actin cytoskeleton, in concert with altered expression of cell type-specific gene products, may be the pathogenic substrate for spine deficits in schizophrenia.
Implications for the Disease Process of Schizophrenia
The disease process of schizophrenia has been proposed to involve pathology in the DLPFC, which contributes to the pathophysiology of psychosis (
9). This hypothesis is supported from a temporal perspective by findings that cognitive deficits, including those that depend on DLPFC circuitry, emerge and progress years before the onset of psychosis (
51,
52). In addition, activation of the DLPFC during cognitive tasks is inversely related to measures of striatal dopaminergic function in subjects with schizophrenia (
15). Evidence for causality in this association was recently provided by findings that deletion of the ARP2/3 complex in mice, resulting in cortical spine deficits, also leads to a subcortical hyperdopaminergia that improves with antipsychotic medications and is reversible with restoration of prefrontal ARP2/3 expression (
13). The findings of the present study demonstrate that this mechanism is plausible in schizophrenia by showing that the ARP2/3 complex signaling pathway is altered in DLPFC pyramidal cells. The resulting loss of dendritic spines on layer 3 pyramidal cells in the DLPFC could represent an upstream pathology that eventually gives rise to excessive dopamine function in the associative striatum and the appearance of psychosis (
10).
Acknowledgments
The authors gratefully acknowledge Mary Brady, B.S., for technical assistance.