Thyroid hormone (T
3 ) has long been of interest to the field of psychiatry because of the effects a variety of psychiatric conditions have on circulating levels of thyroid hormone as well as reports that T
3 may combat depressed mood. T
3 binds to one of several (alpha and beta) nuclear receptors, which then modulates the expression of genes containing binding sequences (T
3 response elements) for the hormone-receptor complex. In peripheral tissues, T
3 increases protein synthesis, cellular metabolism, heat generation, and cardiovascular function. In the brain, T
3 is critical for normal brain development (through effects on neuronal proliferation, organization, arborization, synapse formation, migration, and myelinization) and neuronal performance as well as a variety of neuroendocrine functions, including regulation of the hypothalamic-pituitary-thyroid (HPT) axis, reproduction, and appetite and satiety. The widespread effect of T
3 on the central nervous system is suggested by the high density of T
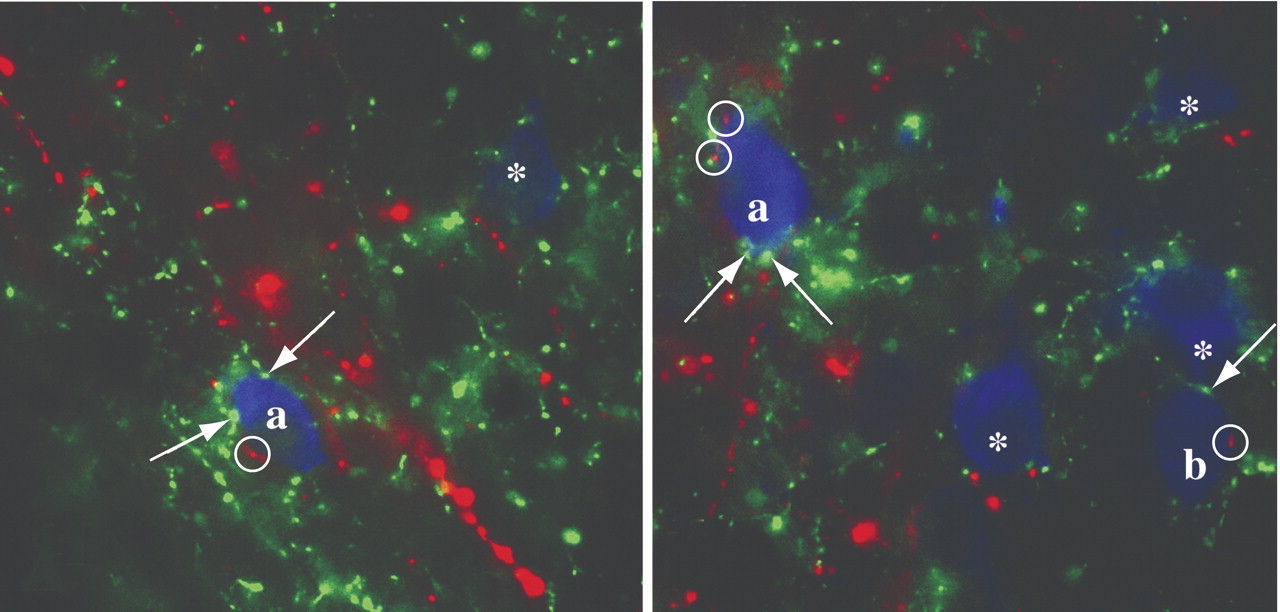
3 receptors in the anterior pituitary, neocortex, hippocampus (dentate gyrus and CA subfields), cerebellum, olfactory bulb, caudate-putamen, and hypothalamus. Maintaining normal circulating levels of thyroid hormone is dependent upon a group of hypothalamic neurons located in the paraventricular nucleus that produce the tripeptide, thyrotropin-releasing hormone (TRH). These neurons regulate the secretion of TRH from the anterior pituitary and hence, the release of thyroid hormone from the thyroid gland. Under certain conditions such as anorexia nervosa, circulating thyroid hormone levels are suppressed primarily due to central inhibition of the HPT axis. A mechanism by which this inhibition may occur is through the afferent input of leptin-sensitive neurons derived from the hypothalamic arcuate nucleus. Two independent groups of neurons in the hypothalamic arcuate nucleus with opposing functions are known to synthesize either alpha-melanocyte stimulating hormone (α-MSH) or agouti-related protein (AGRP), the latter an endogenous antagonist of α-MSH. These neurons exert regulatory control over the HPT axis through a dual innervation of TRH neurons in the paraventricular nucleus (
Figure ). Energy depletion-induced suppression of the HPT axis may be explained, at least in part, by reducing melanocortin signaling at melanocortin 4 receptors as a result of inhibition of α-MSH and stimulation of AGRP production in arcuate nucleus neurons, preventing the activating effects of α-MSH on the phosphorylation of CREB in the nucleus of TRH neurons.