Bipolar disorder and schizophrenia are enduring categories of psychiatric disorder that continue to be based on the presence of characteristic symptoms and clinical course rather than on distinguishing clinical symptoms or confirmatory laboratory investigations. The lack of a validating criterion, the fact that neither disorder breeds true, and the existence of intermediate forms of illness have all led to doubts about the validity of each concept
(1) . Although there is some overlap
(2) in their typical presentations, speech and language abnormalities provide one possible means of making a diagnosis. Loosening of associations, disorganization, and the use of paraphrases or neologisms are all commonly described in the speech of people with schizophrenia. In contrast, mania is typified by copious speech and by flight of ideas, but the links between successive sentences and themes are usually maintained.
Functional magnetic resonance imaging (fMRI) tasks of covert verbal fluency and spontaneous word production are known to activate the dorsal prefrontal and ventral prefrontal cortex, regions implicated in the etiology of psychosis. Several studies have found abnormalities of the ventral prefrontal cortex (including orbitofrontal cortex) and striatal activation in bipolar disorder
(3,
4), whereas studies using similar tasks have found reduced dorsal prefrontal cortex activation
(5,
6) in schizophrenia. These findings, based on indirect comparisons of the two disorders, are of considerable interest, since dorsal areas of the prefrontal cortex have been associated with auditory hallucinations and working memory impairment, core symptoms of schizophrenia
(7), whereas the ventral prefrontal cortex has been implicated in hedonic processes
(8) linked to the core psychopathology of bipolar disorder.
This study sought to test the hypothesis that brain areas involved in contextualized word production separate individuals with bipolar disorder from those with schizophrenia and healthy comparison subjects. Specifically, we hypothesized that circuits involving the ventral prefrontal cortex and striatum would be impaired in patients with bipolar disorder, whereas those involving the dorsal prefrontal cortex would be more impaired in patients with schizophrenia. To test this hypothesis, we used the Hayling Sentence Completion Test in a sample of people with bipolar I disorder or schizophrenia who were selected to have a close affected relative with the same diagnosis. The two clinical groups were also compared with a group of unaffected comparison subjects with no personal or family psychiatric history.
Method
Participants
Individuals with bipolar I disorder or schizophrenia were identified from the case loads of consultant psychiatrists across Edinburgh, Scotland, and the Lothians. Case note diagnoses of bipolar disorder and schizophrenia were established using the Operational Criteria Checklist
(9) and confirmed using the Structured Clinical Interview for DSM-IV (SCID). Unaffected comparison subjects were identified from the same regions and communities as the patients. The diagnostic status of comparison subjects was also confirmed by the SCID. All bipolar and schizophrenia patients were understood to have at least one first- or second-degree relative with the same diagnosis. The diagnosis of affected family members was confirmed wherever possible using the SCID and Operational Criteria Checklist or, when necessary, the Family Interview for Genetic Studies
(10) .
Clinical symptoms were assessed using the Young Mania Rating Scale
(11), Hamilton Depression Rating Scale (HAM-D)
(12), and Positive and Negative Syndrome Scale (PANSS)
(13) . The premorbid full-scale IQ of all subjects was estimated using the National Adult Reading Test
(14) .
Scanning Procedure
Imaging was carried out at the Brain Imaging Research Centre for Scotland (Edinburgh, Scotland, United Kingdom) on a GE 1.5T Signa scanner (GE Medical, Milwaukee) equipped with 23 mT/m “echospeed” gradients, with a rise time of 200 msec. Subjects were imaged with a multislice T2-weighted fast spin-echo sequence [TR/TE=6300/102 msec]. Twenty slices (5-mm thickness, 1.5-mm gap), aligned parallel to the anterior commissure–posterior commissure line, covered the brain. Finally, axial gradient-echo planar images (TR/TE=4000/40 msec; matrix=64×128; field of view=220×440 mm) were acquired continually during the experimental paradigm. Thirty-eight contiguous 5-mm slices were acquired within each TR period. Each echo planar image acquisition was run for 204 volumes, of which the first four volumes were discarded. Visual stimuli were presented using a screen (IFIS; MRI Devices, Waukesha, Wisc.) placed in the bore of the magnet.
The Hayling fMRI Paradigm
The participants in the study performed the verbal initiation section of the Hayling Sentence Completion Test
(15) in the scanner. Subjects were shown sentences with the last word missing and asked to silently think of an appropriate word with which to complete the sentence and then press a button when they had done so. Sentences were selected from a set of sentence completion norms
(16) . The task had four levels of difficulty, according to the range of suitable completion words suggested by the sentence context. Sentences were presented in blocks of fixed difficulty; each block lasted 40 seconds and included eight sentences. Sentences were presented for a period of 3 seconds followed by a fixation cross for 2 seconds. Subjects were instructed to respond at any time (by button press) until the next sentence appeared. The rest condition consisted of viewing a screen of white circles on a black background for 40 seconds. The order of the blocks was pseudorandom, and each block was repeated four times using different sentences. This design allowed both a sentence completion versus rest and parametric analysis (examining areas of increasing activation with increasing task difficulty). Standardized verbal instructions were given prior to scanning.
Immediately after scanning, subjects were given the same sequence of sentences on paper and requested to complete each sentence with the word they first thought of in the scanner. Word appropriateness scores were determined from the word frequency list of sentence completion norms
(16), which provides probabilities of possible responses. A score of 1 was given to the most frequently produced word in the word frequency list, a score of 2 for the next most frequently produced word, etc. Mean reaction times were also determined for each constraint level.
Scan Processing
The echo planar images were converted to ANALYZE format (Mayo Foundation, Rochester, Minn.). Scan analysis was performed using SPM2 (Wellcome Department of Cognitive Neurology, Institute of Neurology, London) running in Matlab (MathWorks, Natick, Mass.). For each subject, echo planar image volumes were realigned to the mean volume in the series using rigid body transformations. Seven subjects presented significant motion artifacts and were excluded from further analysis. The images were then normalized to a study-specific echo planar image template generated from all individuals in the study using a linear affine transformation followed by nonlinear deformation, and resampled using sinc interpolation to cubic voxels of size 8 mm 3 . Normalized images were spatially smoothed with a 6-mm full-width half-maximum Gaussian filter to minimize residual intersubject differences and in order to meet assumptions for statistical analysis.
Statistical Analysis
Statistical analysis was performed using the general linear model as implemented in SPM2. At the individual subject level, the data were modeled with five conditions (the four difficulty levels and the rest condition), each modeled by a boxcar convolved with a synthetic hemodynamic response function. The estimates of the subjects’ movement during the scan were also entered as “covariates of no interest.” Before fitting the model, the subjects’ data were filtered in time using a high-pass filter (400-second cutoff), and temporal auto-correlations were accounted for by imposing an AR
(1) model. Contrasts were constructed to examine all four sentence completion conditions versus rest and areas of increasing activation with increasing task difficulty (the parametric contrast).
For each contrast of interest (sentence completion versus rest and parametric effects), one contrast image per subject was entered into a second-level random effects analysis to examine areas of activation within each of the three groups and differences in activation between the groups (analysis of variance [ANOVA]). Statistical maps were thresholded at a level of p=0.005 (uncorrected), and regions were considered significant at a cluster level of p<0.05, corrected for multiple comparisons. All p values quoted in the text are at the corrected cluster level. Coordinates are reported using the Montreal Neurological Institute convention. Whole brain analyses were supplemented using a prefrontal cortex small-volume correction image.
In order to assess whether between-group differences in activation separated the three groups with potentially clinically useful precision, a discriminant function analysis was conducted. We assumed that the groups could be separated by a straight line on a scatter plot of their activation scores or, in other words, that the discriminant function was linear. The usefulness of fMRI activations as discriminators between the groups was assessed by comparing a subject’s known diagnosis with that predicted from their pattern of fMRI activation.
Neuropsychological Assessment
All patients underwent the intra/extradimensional set shifting task, which is part of the Cambridge Neuropsychological Test Automated Battery. The task consists of nine levels of progressively increasing difficulty. The final level, extradimensional reversal learning, tests the ability of a subject to reverse a previously learned rule. Evidence from functional brain imaging in humans using adapted reversal learning tasks has shown associations between task performance and ventral frontostriatal activations
(17) . In the present study, reversal learning errors were therefore related to ventral prefrontal and striatal activations within the three groups where significant differences in blood-oxygen-level dependent activity had been demonstrated. This analysis was conducted in order to provide further evidence that the findings were not the result of susceptibility artifact and to provide a potential mechanism for impairments shown in other studies.
Functional/Clinical Correlations
Regression analyses were conducted in order to examine the relationship of between-group differences in clinical variables. Significant clusters were used to extract average contrast values for all included subjects and imported into SPSS. These values were then regressed against the scores from the Young Mania Rating Scale and HAM-D and against medication variables to ascertain whether the results reported were potentially confounded by residual psychiatric symptoms or current medication.
Results
Participant Characteristics
One hundred and six participants completed an MRI scan of the brain. All three groups were closely matched for both age and sex, and patients with bipolar disorder had a similar mean age of onset to patients with schizophrenia (
Table 1 ). Premorbid IQ differed significantly among the groups, although the only significant post hoc difference was between subjects with schizophrenia and comparison subjects. Patients with schizophrenia had higher positive symptom rating scores (from the PANSS) than patients with bipolar disorder. Depression and mania rating scores did not differ significantly among the groups.
All participants indicated that they had completed the task as instructed while in the scanner. Seven subjects were, however, excluded because of >4 mm peak-to-peak movement in any direction or >0.6° rotation around any axis. There were no significant differences in any movement parameters, word appropriateness scores, or reaction times among the groups for any of the constraint levels.
Active Versus Rest Analyses
All groups demonstrated the expected pattern of activation for the sentence completion versus rest condition, with widespread activations of the ventral and dorsal prefrontal cortex, temporal cortex, parietal cortex, and striatum, as shown in our previous publications using this task
(6) . In the between-group contrasts, bipolar patients significantly underactivated the left insula (Brodmann’s area, 13) relative to comparison subjects (corrected p=0.002, z=4.46, cluster size=1151 voxels; Montreal Neurological Institute coordinates: –26, –30, 24). Patients with schizophrenia underactivated the left insula (corrected p=0.013, z=4.40, cluster size=809 voxels; Montreal Neurological Institute coordinates: –24, –28, 26), left middle temporal gyrus (Brodmann’s area 20; corrected p=0.003, z=4.12, cluster size=1098 voxels; Montreal Neurological Institute coordinates: –56, –38, –18), and right caudate nucleus (corrected p=0.015, z=4.05, cluster size=790 voxels; Montreal Neurological Institute coordinates: –26, –18, 30) relative to comparison subjects.
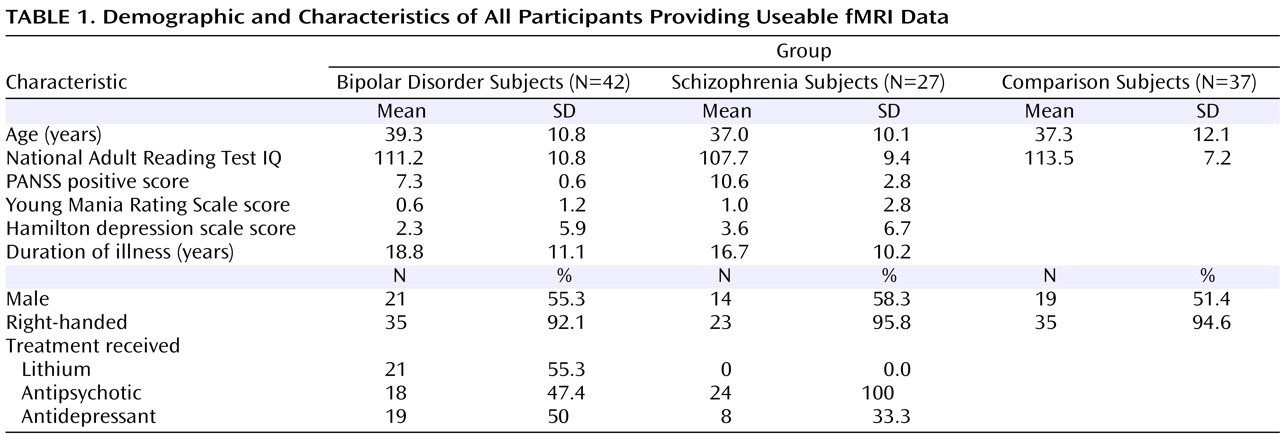
Patients with schizophrenia activated the right insula (Brodmann’s area 13) more strongly than patients with bipolar disorder (corrected p=0.007, z=4.14, cluster size=915 voxels; Montreal Neurological Institute coordinates: 46, –18, 8 [
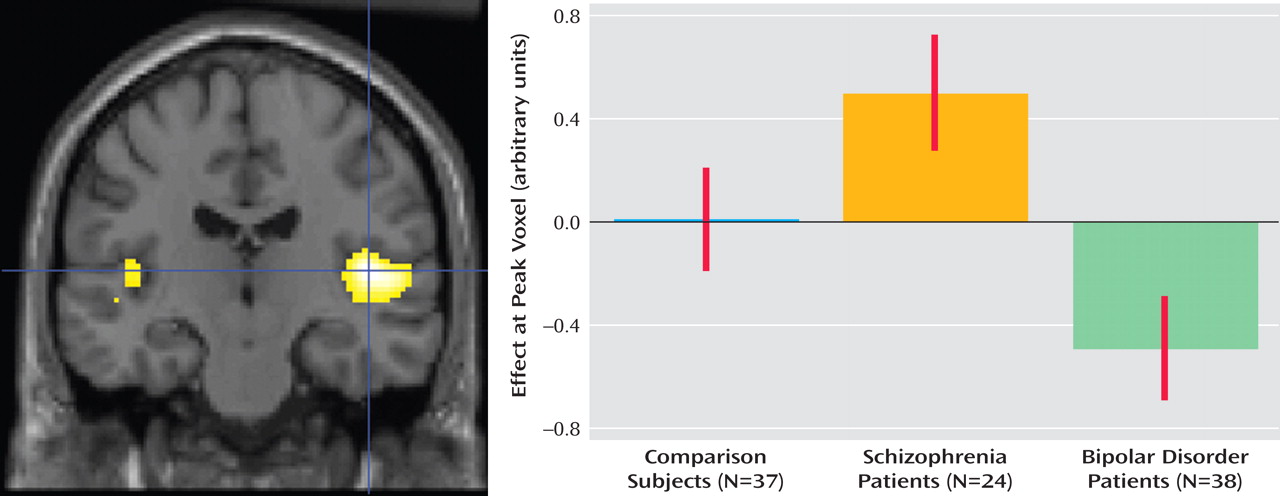
Figure 1 ]), whereas patients with bipolar disorder activated the dorsal prefrontal cortex (Brodmann’s area 9) more strongly than patients with schizophrenia (corrected p=0.015, z=4.10, cluster size=524 voxels; Montreal Neurological Institute coordinates: –12, 56, 42 [
Figure 2 ]).
Parametric Analyses
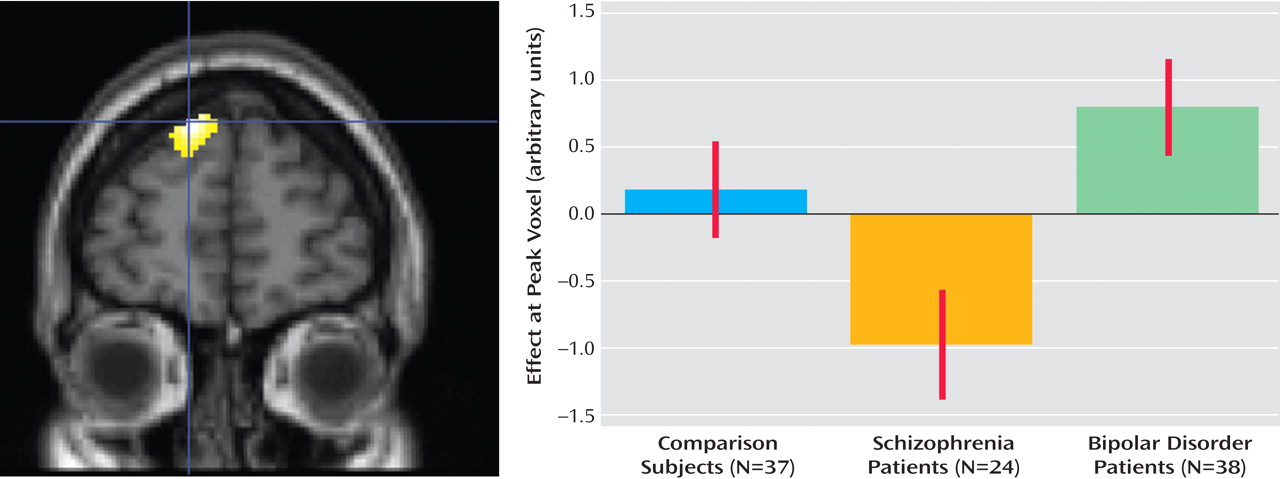
All three groups demonstrated the expected pattern of increasing activation for the linear contrast of decreasing sentence constraint (increasing difficulty), namely, widespread increasing activation of the anterior, medial, and dorsal prefrontal cortex. Patients with bipolar disorder recruited the following three regions of the brain to a greater extent than comparison subjects (
Figure 3 ): the right ventrolateral prefrontal cortex (Brodmann’s area 47; corrected p=0.003, z=4.30, cluster size=1300 voxels; Montreal Neurological Institute coordinates: 38, 42, –4), right ventral striatum (corrected p=0.04, z=4.33, cluster size=747 voxels; Montreal Neurological Institute coordinates: 10, 12, –14), and left caudal middle temporal gyrus (corrected p=0.055, z=4.30, cluster size=691; Montreal Neurological Institute coordinates: –50, –52, 8). Patients with schizophrenia, in contrast, showed no significant differences in activation relative to comparison subjects at the chosen threshold of p=0.005. At lower thresholds of p=0.01 and p=0.05, patients with schizophrenia showed significantly less recruitment of the dorsal prefrontal cortex relative to comparison subjects.
Discriminant Function Analysis
Data were extracted from the left insula cluster (Brodmann’s area 13, cluster 1), which was significant in the active versus rest comparison of bipolar subjects and comparison subjects, and from the left dorsal prefrontal cluster (cluster 2) and right insula cluster (cluster 3), which were significantly different in the bipolar versus schizophrenia comparison using the same contrast of sentence completion versus rest. Data were also extracted from the three regions (ventrolateral orbitofrontal cortex: cluster 4; ventral striatum: cluster 5; middle temporal gyrus: cluster 6) that appeared to differentiate bipolar subjects from comparison subjects in the parametric contrast of decreasing sentence constraint.
The model that most clearly differentiated bipolar subjects and comparison subjects included the following two clusters: the ventral striatum (cluster 5) and middle temporal gyrus (cluster 6). This model correctly classified 71.1% of bipolar subjects and 75.7% of comparison subjects. The model that most efficiently classified bipolar and schizophrenia subjects also included the following two variables: the dorsal prefrontal cortex and right insula. This correctly classified 92.1% of bipolar subjects and 58.7% of participants with schizophrenia.
Relationship With Reversal Learning and Clinical Variables
The significant ventral prefrontal and ventral striatal clusters were used to extract average values for the parametric contrast of decreasing sentence constraint over these areas in the bipolar patients. Reversal learning errors were significantly negatively associated with the gradient of response in both the orbitofrontal (r s =0.60, p=0.003) and ventral striatal (r s =–0.55, p=0.008) regions in patients with bipolar disorder. There was no significant relationship between reversal learning errors and either orbitofrontal (r s =0.02, p=0.92) or striatal (r s =0.37, p=0.07) activations in the comparison group.
An increased gradient of activation of the orbitofrontal cortex was also found to be positively associated with PANSS positive scores in bipolar subjects (r=0.46, p=0.004). Nevertheless, differences between bipolar subjects and comparison subjects remained significant after PANSS positive scores were included in the statistical model (p=0.025). No further associations were found between any significant cluster and any class of medication or other symptom type in either bipolar or schizophrenia patients.
Discussion
A key objective in studies of major mental illness has been to demonstrate biological variables that differentiate people with schizophrenia from those with affective disorders. To our knowledge, the present study is the first to show that altered patterns of dorsal prefrontal cortex and insula activity are able to differentiate individuals with schizophrenia from individuals with affective disorders with potentially clinically useful precision. In addition, we found new evidence, using a parametric contrast of increasing difficulty, that patients with bipolar disorder show altered patterns of orbitofrontal cortex and ventral striatal activity relative to comparison subjects. This finding was further supported by correlation between the gradient of activation in bipolar disorder patients and performance on tests of reversal learning known to rely on the orbitofrontal cortex and related circuitry. These findings confirm our original hypothesis that circuits involving the dorsal prefrontal cortex are implicated in the pathophysiology of schizophrenia, whereas circuits involving the lateral orbitofrontal cortex are more specifically implicated in bipolar disorder.
Reduced activation of the dorsal prefrontal cortex was found in subjects with schizophrenia and clearly separated them from patients with bipolar disorder. Abnormalities of the dorsal prefrontal cortex structure and function have been widely replicated in schizophrenia
(5,
18) . The dorsal prefrontal cortex is a region that has been implicated in several cognitive functions, including working memory and attentional processes
(19) . Several studies have shown reductions in activation relative to comparison subjects, and there is considerable evidence that abnormalities may be expressed premorbidly
(20) and relate to the action of specific susceptibility alleles
(21,
22) . Functional deficits in the dorsal prefrontal cortex were found by Yurgelun-Todd et al.
(23) in bipolar disorder, although patients had high levels of affective symptoms at the time of the scan, potentially reducing their performance. In contrast, the patients selected for the present study had low levels of symptoms, task performance was similar among the three groups, and there was no relationship between symptoms and dorsal prefrontal cortex activation. This suggests that the observed differences are not simply the result of differences in an individual’s current mental state and may represent a difference between the neurobiological substrates of schizophrenia and bipolar disorder.
Right anterior insula activation was significantly decreased in subjects with bipolar disorder relative to those with schizophrenia and clearly separated the two clinical groups. The insula has been implicated in language processing
(24), unsuccessful response inhibition
(25), and emotional processing
(26) in a number of studies. The differences seen in the present study may therefore relate to inhibitory system impairments in bipolar disorder. Functional abnormalities of the insula in schizophrenia are well replicated, and there is some evidence that increased activation is related to auditory hallucinations
(27) . Two studies
(28,
29) have shown increased activation of this region in bipolar disorder using different tasks, and Kruger et al.
(30) reported that activation may be associated with low mood. No association was found between insular activation and mood state in the present study.
The finding of an increased gradient of orbitofrontal cortex and ventral striatal activation in bipolar patients relative to comparison subjects suggests that the orbitofrontal cortex and striatum may be sensitized to respond to nonemotional stimuli in bipolar subjects. This finding supports earlier work showing abnormal activation of the ventral prefrontal cortex and ventral striatum during several tasks
(3,
4,
31 –
34) . The inconsistency in direction of effect in earlier studies may be related to task performance or load-response relationship. These potential explanations can, however, only be addressed with detailed performance data and parametric contrasts, such as those provided in the present study. The lateral orbitofrontal cortex may have a role in inhibiting behavior in a variety of context, and the ventral striatum is understood to mediate the motivational responses to emotional stimuli. Both regions are understood to be involved in affect regulation
(35,
36), and evidence from structural imaging of unaffected relatives would suggest that abnormalities of these regions may be related to a trait vulnerability to bipolar disorder
(37,
38) . Greater activation in the lateral orbitofrontal cortex and ventral striatum in bipolar disorder may reflect an underlying inefficiency in inhibiting inappropriate behavior, which is compensated in well individuals by increased cortical recruitment. We would speculate that during manic episodes, compensation for this deficit ceases.
The present study is one of the first, and currently the largest, study to examine the diagnostic specificity of fMRI findings in schizophrenia and bipolar disorder. Curtis et al.
(39) examined patterns of prefrontal activation in 15 male subjects using a verbal fluency and semantic decision task. The results show little overlap with our own, although greater activations were found in bipolar subjects relative to comparison subjects in the inferior frontal gyrus.
Although we could find no other fMRI studies comparing patients with schizophrenia to those with bipolar disorder, there are many comparing patients with schizophrenia or bipolar disorder with a comparison group. The confounding role of affective symptoms and the lack of data on task performance are limitations to much of the published literature in respect to bipolar disorder. The present study selected a clinically stable sample of patients from the community and specifically aimed to balance task performance across groups. The results presented are arguably more generalizable to the population of bipolar patients as a whole compared with studies that consider severely symptomatic or euthymic patients alone. The HAM-D and Young Mania Rating Scale scores in the present study were low, and the activations shown were not related to residual affective symptoms.
We have shown that there are important differences between patients with schizophrenia and bipolar disorder in terms of brain activation on fMRI that substantially differentiate between these diagnostic groups. Importantly, patients with schizophrenia and bipolar disorder were studied in the same experiment, using a single functional imaging paradigm in which results were not confounded by performance differences. These findings confirm our hypothesis that a circuit involving the dorsal prefrontal cortex is involved in the etiology of schizophrenia, while an emotional network involving the ventral prefrontal cortex, ventral striatum, and insula is involved in the etiology of bipolar disorder.